2.2. 508-Da MAA with an Absorption Maximum at 334 nm
A MAA with an absorption maximum at 334 nm (
Figure 1) was purified from the field-isolated natural
N. commune colonies (
Table 1,
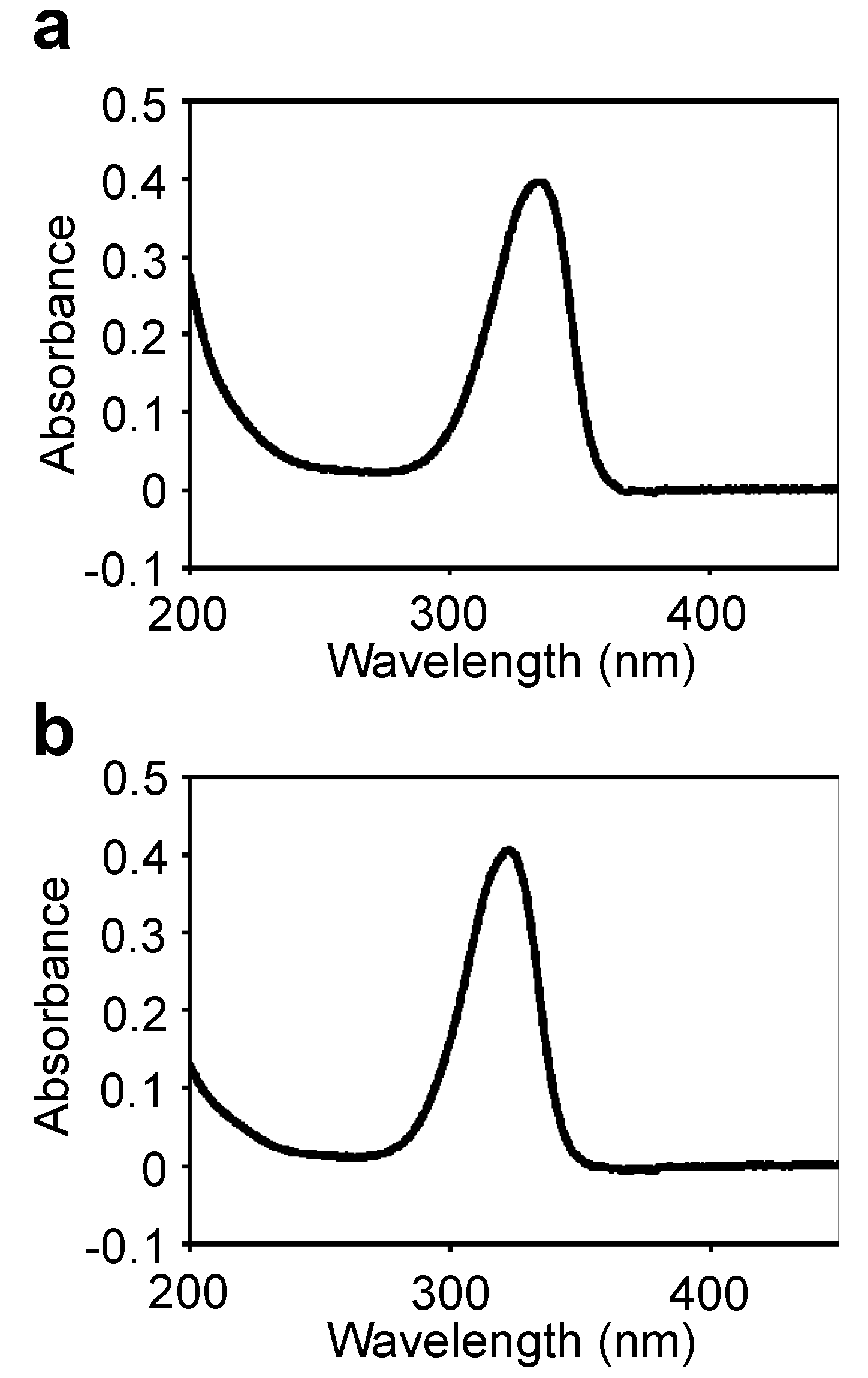
Figure S1). The UV absorption spectrum of the purified MAA showed a single absorption peak at 334 nm (
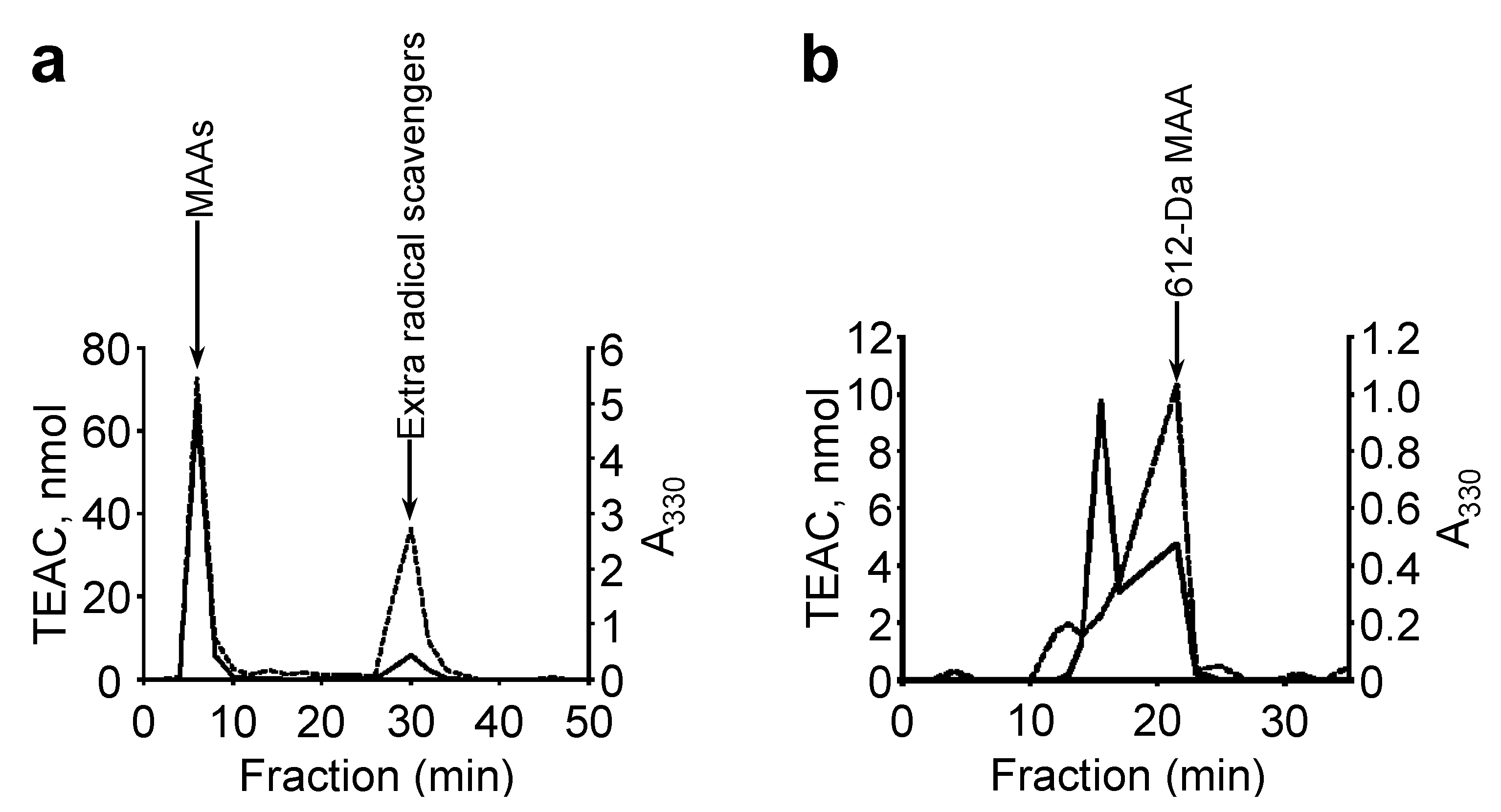
Figure 2a). The absorption maximum shifted reversibly to 332 nm in a high acidic solution (pH < 2) and to 334 nm in alkaline solutions. The molecular mass was 508 Da, according to MALDI-TOF MS (
Table 2). The absorption coefficient of this MAA in water was 71.4 L·g
−1·cm
−1 at 334 nm and the calculated molar absorption coefficient at 334 nm was 3.63 × 10
4 M
−1·cm
−1. Because no MAA with a molecular mass of 508 Da had been previously reported, the chemical structure of the 508 Da MAA was further characterized.
Table 1.
Purification of 508-Da, 450-Da and 612-Da mycosporine-like amino acids (MAAs) with absorption maxima at 334, 322 and 322 nm, respectively, from N. commune a.
Table 1.
Purification of 508-Da, 450-Da and 612-Da mycosporine-like amino acids (MAAs) with absorption maxima at 334, 322 and 322 nm, respectively, from N. commune a.
| Step | Volume (mL) | MAA concentration b (mg·L−1) | MAA amount (mg) | Yield (%) |
|---|
| Water extract | 640 | 19.4 | 12.4 | 100 |
| 70% Ethanol solution | 1960 | 5.4 | 10.6 | 85 |
| Vacuum concentration | 3.5 | 2374 | 8.3 | 67 |
| Reverse-phase HPLC | 508-Da | 0.5 | 3525 | 1.8 | 15 |
| 450-Da | 0.5 | 1969 | 1.0 | 8 |
| 612-Da | 0.5 | 2404 | 1.2 | 10 |
| Gel filtration | 508-Da | 0.5 | 2474 | 1.2 | 10 |
| 450-Da | 0.5 | 1782 | 0.9 | 7 |
| 612-Da | 0.5 | 2226 | 1.1 | 9 |
Figure 2.
UV absorption spectra of the purified MAAs in H2O. (a) 508-Da MAA with absorption maxima at 334 nm (ε = 3.63 × 104 M−1·cm−1 at 334 nm); (b) 612-Da MAA with absorption maximum at 322 nm (ε = 2.82 × 104 M−1·cm−1 at 322 nm).
Figure 2.
UV absorption spectra of the purified MAAs in H2O. (a) 508-Da MAA with absorption maxima at 334 nm (ε = 3.63 × 104 M−1·cm−1 at 334 nm); (b) 612-Da MAA with absorption maximum at 322 nm (ε = 2.82 × 104 M−1·cm−1 at 322 nm).
Table 2.
Summary of MALDI-TOF MS analysis of the purified 508-Da MAA with absorption maximum at 334 nm.
Table 2.
Summary of MALDI-TOF MS analysis of the purified 508-Da MAA with absorption maximum at 334 nm.
| | Mass of fragment | Relative abundance | Neutral loss | Deleted fragment |
|---|
| MS of purified MAA | 509.12 | 100 | | |
| 465.13 | 81 | 44 | CO2 |
| 451.12 | 10 | 58 | CO2 + CH2 |
| 427.07 | 27 | 82 | - |
| 425.04 | 58 | 84 | - |
| 421.15 | 58 | 88 | 2CO2 |
| 419.15 | 24 | 90 | CO2 + 2CH2 + H2O |
| 407.10 | 40 | 102 | 2CO2 + CH2 |
| 403.13 | 37 | 106 | 2CO2 + H2O |
| 387.11 | 57 | 122 | - |
| 385.07 | 35 | 124 | 2CO2 + 2H2O |
| MS2 of m/z 509 | 509.12 | | | |
| 347.27 | 100 | 162 | Hexose |
| 303.24 | 6 | 206 | Hexose + CO2 (or C2H4O) |
| 279.20 | 7 | 230 | - |
| MS2 of m/z 465 | 465.13 | | | |
| 303.29 | 100 | 162 | Hexose |
| 285.27 | 3 | 180 | Hexose + H2O |
| 235.22 | 4 | 230 | - |
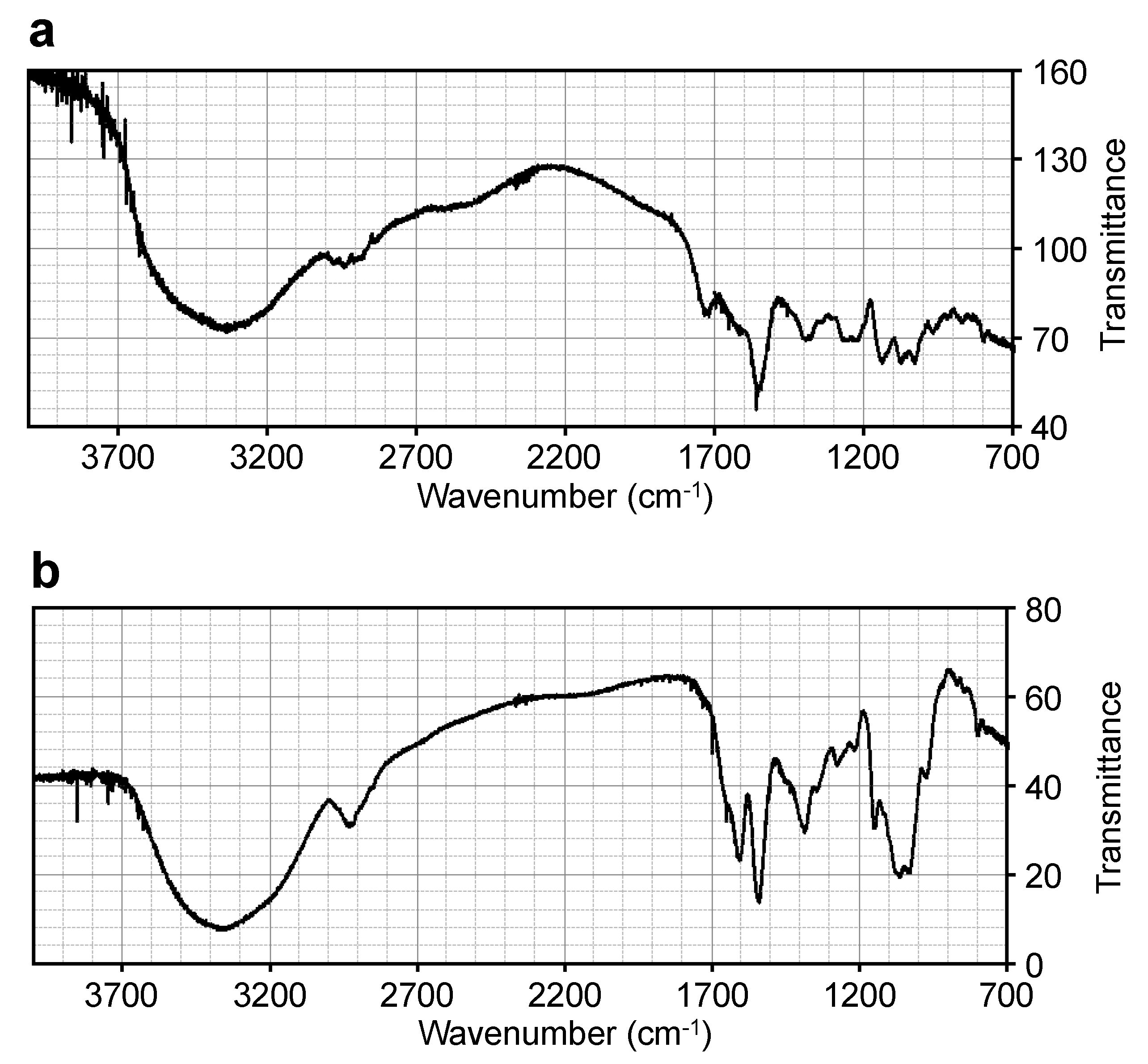
The IR spectrum of the 508-Da MAA (
Figure 3a) was compared to those of known MAAs, including porphyra-334 [
35] and a glycosylated porphyra-334 with a molecular mass of 478 Da from
N. commune [
29] (
Table 3). Similar IR absorption peaks were observed, which suggested that the 508-Da MAA had a similar structure to the known MAAs. The absorption peak at 1560 cm
−1 in the 508-Da MAA indicated the presence of the conjugated imine as a characteristic structure of MAA chromophore, corresponding to those at 1540 cm
−1 in porphyra-334 and at 1558 cm
−1 in the 478-Da glycosylated porphyra-334 derivative. The characteristic absorption peak at 3334 cm
−1 in the 508-Da MAA indicated the presence of several hydroxyl functional groups, and the absorption peaks in the range of 1200–1300 cm
−1 suggested the presence of a sugar, which was consistent with the predicted structure as described below.
MALDI-TOF MS/MS analysis was performed on the parent molecular ion fragment with
m/
z 509 and molecular ion fragment with
m/
z 465 (
Table 2). A fragment with
m/
z 347, a molecular mass identical to that of porphyra-334 [
36,
37], was detected in the second MS of the fragment with
m/
z 509. The neutral loss of 162 Da suggested the deletion of a hexose (C
6H
10O
5) from the fragment with
m/
z 509. The fragment with
m/
z 303 suggested the deletion of CO
2. The fragment with
m/
z 303 was also detected in the second MS of the fragment with
m/
z 465. The deletion of 162 Da from the fragment with
m/
z 465 to yield
m/
z 303 indicated the presence of a hexose in the MAA structure (Table 2). Additionally, in the MS analysis the fragments with
m/
z 465 and
m/
z 421 suggested the consecutive deletion of CO
2 and the presence of two carboxyl groups in the MAA structure (
Table 2). These fragmentation patterns were similar to those from the MS analyses of known MAAs [
38]. These data suggest that the 508-Da MAA is a hexose-bound porphyra-334 derivative.
Figure 3.
FT-IR spectra of (a) the 508-Da MAA with an absorption maximum at 334 nm and (b) the 612-Da MAA with an absorption maximum at 322 nm.
Figure 3.
FT-IR spectra of (a) the 508-Da MAA with an absorption maximum at 334 nm and (b) the 612-Da MAA with an absorption maximum at 322 nm.
Table 3.
Comparison of the IR spectra of the purified MAAs with that of porphyra-334.
Table 3.
Comparison of the IR spectra of the purified MAAs with that of porphyra-334.
| 508-Da MAA with Amax at 334 nm | Porphyra-334 | 478-Da MAA with Amax at 335 nm |
|---|
| Wavenumber (cm−1) |
|---|
| | | 3400 |
| 3334 | | |
| | 3300 | |
| 2942, 2975 | | |
| 1718 | | |
| | 1600 | 1606 |
| 1560 | 1540 | 1558 |
| 1379, 1395 | 1380 | 1382 |
| 1301, 1355 | | 1310 |
| 1231, 1257 | | 1273 |
| 1138 | | 1132 |
| 1075 | 1080 | 1072 |
| 1030 | | 1006 |
| 968 | | |
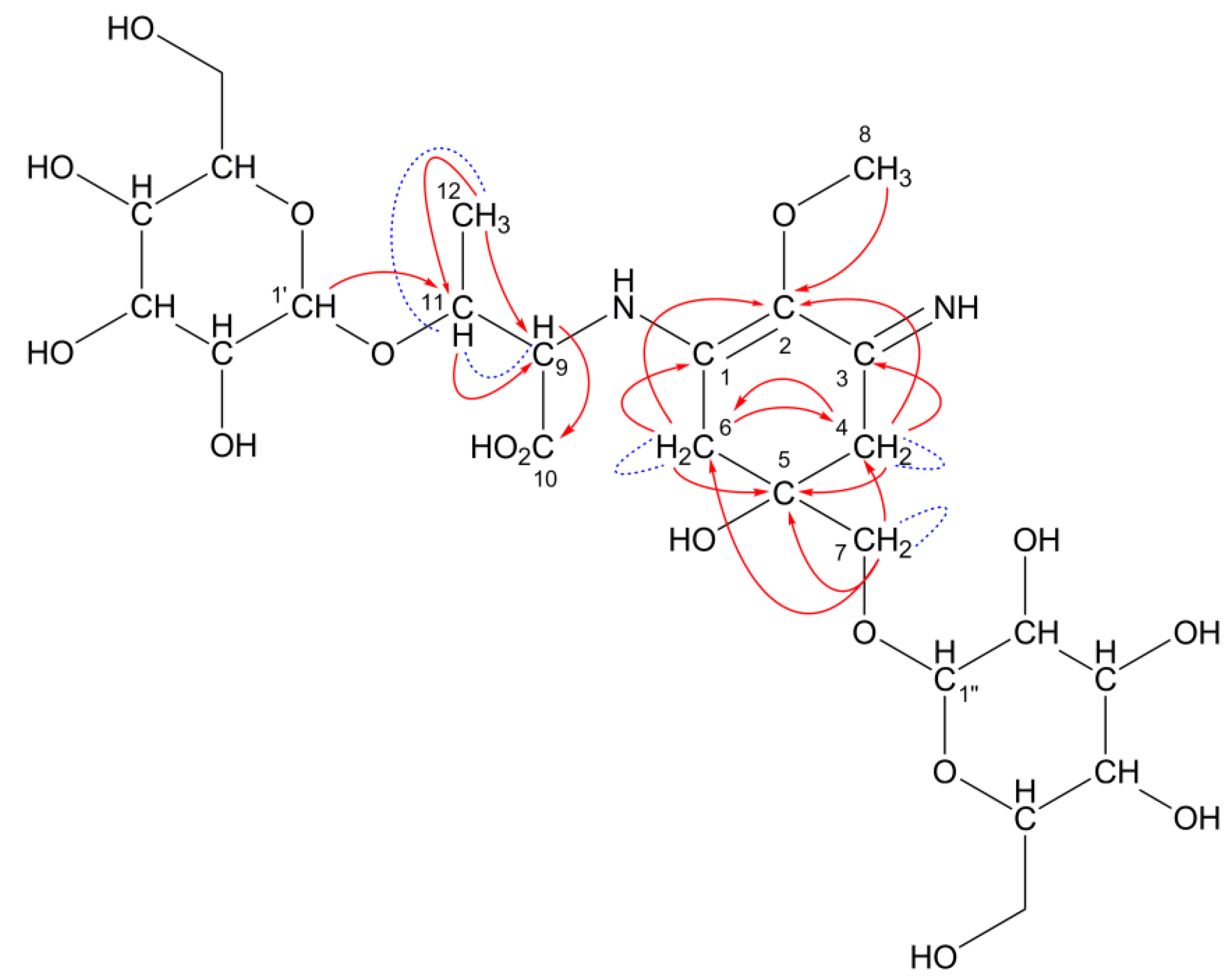
This predicted structure was confirmed by NMR spectroscopic analysis. Both the known chemical shifts for porphyra-334 and hexose were observed in the 508-Da MAA (
Table 4). Typical
13C-chemical shifts assignable to a cyclohexenimine chromophore (C1, 2, 3, 4, 5, 6) and amino acid substituents (C9, 10, 11, 12, 13, 14) of the 508-Da MAA were observed at the same regions in porphyra-334. The characteristic coupling pattern (AB quartet) of methylene protons at C4 and C6 was determined (
J = 17.4 Hz), which is not shown in porphyra-334. It was hard to identify which carbon (C4 and C6) should be assigned to the glycine-substituted side and
vice versa because chemical shifts at C9 and C11 protons were very close to recognize the correlation in heteronuclear multiple bond correlation (HMBC) spectrum. However, the
13C-chemical shifts in the 508-Da MAA were highly similar to those in the pentose-bound porphyra-334 derivative, which has been reported previously (
Table 4). The correlations in COSY and HMBC spectra are summarized in
Table 5 and shown in
Figure 4 as red arrows and blue dashed lines, respectively. Characteristic correlations assignable to the MAA backbone appeared and a probable HMBC correlation suggesting a putative hexose-binding site was observed (shown as red dashed arrow in
Figure 4).
Table 4.
Summary of the NMR analysis of the 508-Da MAA with an absorption maximum at 334 nm.
Table 4.
Summary of the NMR analysis of the 508-Da MAA with an absorption maximum at 334 nm.
| C | 508-Da MAA with Amax at 334 nm | 478-Da pentose-bound porphyra-334 | Porphyra-334 |
|---|
| | 13C | 1H | 13C | 1H | 13C | 1H |
|---|
| 1 a | 161.4 | - | 161.6 | - | 161.6 | - |
| 2 | 128.6 | - | 128.4 | - | 126.0 | - |
| 3 a′ | 163.4 | - | 163.2 | - | 163.2 | - |
| 4 b | 36.1 | 2.82, 3.03 (ABq, 17.4) | 36.4 | 3.00 | 32.5 | 2.75 |
| 5 | 73.3 | - | 73.1 | - | 71.3 | - |
| 6 b′ | 36.6 | 2.80, 2.91 (ABq, 17.4) | 35.9 | 2.83 | 33.0 | 2.77 |
| 7 | 76.1 | 3.47, 3.76 (d, 10.1, each) | 77.7 | 3.90, 3.67 | 67.1 | 3.61 (s) |
| 8 | 62.3 | 3.67 (s) | 62.2 | 3.70 (s) | 59.0 | 3.73 (s) |
| 9 | 49.6 | 4.09 (d, 1.8) | 49.5 | 4.06 (d, 2.4) | 47.0 | 4.07 (s) |
| 10 c | 177.6 | - | 177.7 | - | 177.6 | - |
| 11 | 67.5 | 4.10 (d, 4.6) | 67.3 | 4.09 (d, 4.5) | 64.0 | 4.12 (d, 5.0) |
| 12 c′ | 178.2 | - | 178.2 | - | 178.0 | - |
| 13 | 74.4 | 4.34 (dq, 4.6, 6.4) | 71.0 | 4.31 (dq, 4.5, 6.5) | 68.0 | 4.33 (m, 5.0, 6.4) |
| 14 | 22.4 | 1.24 (d, 6.4) | 22.3 | 1.26 (d, 6.5) | 19.0 | 1.26 (d, 6.4) |
| | | | | | Methyl α-
d-Man |
| | | | | | 13C | 1H |
| 1′ | 101.8 | 4.96 (d, 3.2) | 106.4 | 4.37 (d, 7.6) | 102.2 | 4.66 |
| 2′ | 71.4 | 3.88 | 73.5 | 3.60 (dd, 7.6, 9.6) | 71.4 | 3.82 |
| 3′ | 72.2 | 3.97 | 75.0 | 3.68 (dd, 9.6, 3.6) | 72.1 | 3.65 |
| 4′ | 70.9 | 3.82 | 71.1 | 3.95 (m) | 68.3 | 3.53 |
| 5′ | 72.3 | 3.80 | 69.1 | 3.92, 3.65 | 73.9 | 3.51 |
| 6′ | 64.2 | 3.55, 3.72 | | | 62.5 | 3.79, 3.65 |
Table 5.
Characteristic correlations in COSY and HMBC spectra assignable to the 508-Da MAA backbone.
Table 5.
Characteristic correlations in COSY and HMBC spectra assignable to the 508-Da MAA backbone.
| | COSY | HMBC |
|---|
| H4 | H4 | C2, C3, C5, C6 |
| H6 | H6 | C1, C2, C4, C5 |
| H7 | H7 | C4, C5, C6 |
| H8 | | C2 |
| H9 | | C3, C10 |
| H11 | H13 | C1, C12 |
| H13 | H11, H14 | |
| H14 | H13 | C11, C13 |
| H1′ | | C7 a |
Figure 4.
A predicted structure for the 508-Da MAA with an absorption maximum at 334 nm and a molecular formula of C20H32N2O13. Hexose is bound to porphyra-334 at C7 position. Red arrows and blue dashed lines represent the apparent HMBC and COSY correlations, respectively. The red dashed arrow represents probable HMBC correlation.
Figure 4.
A predicted structure for the 508-Da MAA with an absorption maximum at 334 nm and a molecular formula of C20H32N2O13. Hexose is bound to porphyra-334 at C7 position. Red arrows and blue dashed lines represent the apparent HMBC and COSY correlations, respectively. The red dashed arrow represents probable HMBC correlation.
The presence of a hexose moiety in the 508-Da MAA was confirmed by the
13C chemical shift in methyl α-
d-mannose; chemical shift at the anomer carbon (C1′) was almost the same as that in methyl α-
d-mannose, indicating that the substitution position is at the anomer position. Remaining peaks were assigned to one CH
2 (C6′) and four CH carbon (C2′, 3′, 4′, 5′) from the result of Distortionless Enhancement by Polarization Transfer (DEPT) spectrum. The chemical shifts of these peaks were also consistent with those of methyl α-
d-mannose. In the 508-Da MAA, the hexose binding position could be thought of as at C7 position because the CH
2 protons at C7 position coupled to each other (
J = 10.1 Hz). This coupling pattern was not observed in porphyra-334 but was observed in the pentose-bound 478-Da MAA (
Table 4). In our previous report, the pentose-binding position at the C7 methylene was identified because of the apparent correlation between the C7 carbon and the anomer (C1′) proton in the HMBC spectrum [
29]. In the 508-Da MAA, the correlation between C7 and H1′ was not observed apparently (
Table 5,
Figure 4 red dashed arrow), but geminal coupling at C7 protons suggesting magnetic anisotropy was observed. Thus the high rotation barrier around C5–C7 bond due to a large substituent can be predicted and this idea agrees with the presence of a monosaccharide at C7 position of the 508-Da MAA. The dihedral angle dependence of C5–C7 rotational energy was calculated by using Chem-3D software. The rotational barrier of the C7 substituted porphyra-334 was much greater than that of porphyra-334, which cannot be surmounted at room temperature (data not shown). This simulation can be supported by the previous report concerning neocedumoside derivatives who have a similar chemical structure around a hexose-binding site [
40].
Table 6 shows the chemical shifts of the hexose-binding sites of the 508-Da MAA, porphyra-334, neocedumoside and its hydrolysate. The geminal coupling of magnetically anisotropic methylene protons (
J = 10.1 Hz) was detected due to the deshielding effect of the neighboring σ-electron. The equatorial proton was observed at lower magnetic field region by 0.43 ppm rather than the axial proton. The
1H-chemical shifts at C7 position in the 508-Da MAA showed two apparently separated signals by 0.29 ppm with geminal coupling between them. The hydrolysis of neocedumoside results in axial and equatorial peaks getting closer to their center of balance [
40]. The center of balance of two separated signals in the 508-Da MAA was 3.61 ppm, which is identical to the corresponding chemical shift in porphyra-334. These data support the idea that magnetic anisotropy of methylene protons is caused by the substitution of hexose. In addition, the
13C chemical shift in neocedumoside is downfield shifted by 8.2 ppm from that of hydrolysate, suggesting a characteristic NMR feature on glycolysis. A similar shift (9.0 ppm) from porphyra-334 was also observed in the 508-Da MAA. These NMR analyses and molecular simulation support the substitution of hexose at C7 position.
Figure 4 shows the predicted structure of the 508-Da MAA from
N. commune.
Table 6.
Comparison of 13C and 1H chemical shifts at the hexose binding site of the 508-Da MAA, porphyra-334, neocedumoside and its hydrolysate.
Table 6.
Comparison of 13C and 1H chemical shifts at the hexose binding site of the 508-Da MAA, porphyra-334, neocedumoside and its hydrolysate.
| | 508-Da MAA | Porphyra-334 | Neocedumoside | Neocedumoside-hydrolysate |
|---|
| 13C | 76.1 | 67.1 | 79.0 | 70.8 |
| 1H | 3.47 (d, 10.1)
3.76 (d, 10.1) | 3.61 (s) | 3.40 (d, 10.1)
3.83 (d, 10.1) | 3.51 (d, 11.0)
3.54 (d, 11.0) |
2.3. 612-Da MAA with an Absorption Maximum at 322 nm
An MAA with an absorption maximum at 322 nm (
Figure 1) was purified from the same
N. commune colonies (
Table 1,
Figure S1). The UV absorption spectrum of the purified MAA showed a single absorption peak at 322 nm (
Figure 2b); similar to the 508-Da MAA, the absorption maximum reversibly shifted to 320 nm in a highly acidic solution. The molecular mass was 612 Da, according to MALDI-TOF MS (
Table 7). The accurate molecular mass was determined by FAB MS to predict the elemental composition. A molecular ion fragment with
m/
z 613.2462 was detected and its predicted molecular formula was C
24H
41N
2O
16 within 1 ppm error. The absorption coefficient of this MAA in water was 46.07 L·g
−1·cm
−1 at 322 nm, and its calculated molar absorption coefficient at 322 nm was 2.82 × 10
4 M
−1·cm
−1. Because no MAA with a molecular mass of 612 Da had been previously reported, the chemical structure of the 612 Da MAA was further characterized.
Table 7.
Summary of MALDI-TOF MS analysis of the purified 612-Da MAA with absorption maximum at 322 nm.
Table 7.
Summary of MALDI-TOF MS analysis of the purified 612-Da MAA with absorption maximum at 322 nm.
| | Mass of fragment | Relative abundance | Neutral loss | Deleted fragment |
|---|
| MS of purified MAA | 613.17 | 18 | | |
| 569.19 | 95 | 44 | CO2 |
| 451.12 | 4 | 162 | Hexose |
| 407.13 | 17 | 206 | CO2 + Hexose |
| 389.12 | 100 | 224 | CO2 + Hexose + H2O |
| 349.09 | 21 | 264 | Hexose + C4H6O3 |
| 227.02 | 16 | 386 | CO2 + 2Hexose + H2O |
| MS2 of
m/z 613 | 613.17 | | | |
| 451.21 | 100 | 162 | Hexose |
| 407.21 | 1 | 206 | Hexose + CO2 |
| 389.23 | 8 | 224 | Hexose + CO2 + H2O |
| 317.20 | 6 | 296 | - |
| 289.18 | 14 | 324 | 2Hexose |
| 241.17 | 10 | 372 | - |
| 185.14 | 6 | 428 | - |
| 91.10 | 6 | 522 | - |
| MS2 of
m/z 569 | 569.19 | | | |
| 407.37 | 100 | 162 | Hexose |
| 392.32 | 13 | 177 | Hexose + CH3 |
| 389.33 | 37 | 180 | Hexose + H2O |
| 375.31 | 10 | 194 | Hexose + H2O + CH2 |
| 245.25 | 26 | 324 | 2Hexose |
| 227.24 | 16 | 342 | 2Hexose + H2O |
| 209.22 | 21 | 360 | 2Hexose + 2H2O |
| 191.21 | 8 | 378 | 2Hexose + 3H2O |
| 177.19 | 18 | 392 | 2Hexose + 3H2O + CH2 |
| 91.10 | 12 | 478 | - |
| MS2 of
m/z 451 | 451.21 | | | |
| 407.00 | 2 | 44 | CO2 |
| 289.20 | 100 | 162 | Hexose |
| 245.17 | 6 | 206 | Hexose + CO2 |
| 227.19 | 2 | 224 | Hexose + CO2 + H2O |
| 170.13 | 1 | 281 | Hexose + CO2 + H2O + C3H5O |
| 151.14 | 1 | 300 | Hexose + CO2 + 2H2O + C3H6O |
| 91.10 | 2 | 360 | - |
| MS2 of
m/z 407 | 407.13 | | | |
| 245.28 | 100 | 162 | Hexose |
| 230.24 | 3 | 177 | Hexose + CH3 |
| 227.25 | 15 | 180 | Hexose + H2O |
| 209.23 | 5 | 198 | Hexose + 2H2O |
| 199.21 | 4 | 208 | Hexose + C2H6O |
| 177.21 | 6 | 230 | Hexose + 3H2O + CH2 |
| 91.11 | 3 | 316 | - |
| MS2 of
m/z 389 | 389.12 | | | |
| 227.29 | 100 | 162 | Hexose |
| 212.24 | 9 | 177 | Hexose + CH3 |
| 209.26 | 36 | 180 | Hexose + H2O |
| 191.24 | 54 | 198 | Hexose + 2H2O |
| 181.21 | 17 | 208 | Hexose + C2H6O |
| 170.19 | 8 | 219 | Hexose + C3H5O |
| 159.20 | 9 | 230 | - |
| 91.11 | 12 | 298 | - |
| MS2 of
m/z 349 | 349.09 | | | |
| 187.19 | 100 | 162 | Hexose |
| 169.18 | 8 | 180 | Hexose + H2O |
| 151.16 | 25 | 198 | Hexose + 2H2O |
Similar IR absorption peaks were observed when the 612-Da MAA IR spectrum (
Figure 3b) was compared with those from the known MAA palythine triacetate [
41] and the 1050-Da glycosylated MAA from
N. commune [
29] (
Table 8), which suggested that the 612-Da MAA had a similar structure to the known MAAs. The characteristic absorption peak at 3370 cm
−1, but not that at 3310 cm
−1, in the 612-Da MAA suggested the presence of a large number of hydroxyl groups, and the absorption peaks in the range of 1200–1300 cm
−1 suggested the presence of sugars, which was consistent with the presence of two hexose rings as described below. The absorption peak at 1542 cm
−1 suggested the presence of the conjugated imine in the 612-Da MAA.
MALDI-TOF MS/MS analysis was performed on the parent molecular ion fragment with
m/
z 613 and on the other main molecular ion fragments with
m/
z 569, 451, 407, 389 and 349 (
Table 7). A plausible fragmentation pattern could be suggested with regard to the mass spectra (
Figure 5). The ion fragment with
m/
z 569 indicated the deletion of CO
2 from the parent molecular ion fragment with
m/
z 613, suggesting the presence of a carboxyl group. Subsequent deletions of 162 and 18 Da from the fragment with
m/
z 569 to produce the fragments with
m/
z 407 and
m/
z 389 indicated the deletions of hexose (C
6H
10O
5) and H
2O, respectively. MS/MS analysis of the fragments with
m/
z 407 and 389 led to the fragment at
m/
z 227, which indicated the deletion of another hexose (
Table 7 and
Figure 5).
Table 8.
Comparison of IR spectra of the 612-Da MAA with known MAAs.
Table 8.
Comparison of IR spectra of the 612-Da MAA with known MAAs.
| 612-Da MAA with Amax at 322 nm | Palythine triacetate | 1050-Da MAA with Amax at 312 and 340 nm |
|---|
| Wavenumber (cm−1) |
|---|
| 3370 | | 3399 |
| | 3310 | |
| 2932 | | 2929 |
| | 1725, 1740 | |
| | 1660 | |
| 1607 | 1590 | 1617 |
| 1542 | 1535–1555 | 1541 |
| 1388 | | 1400 |
| 1346 | | |
| 1277, 1216 | | 1275 |
| 1150 | | |
| 1067, 1036 | | 1076, 1046 |
| 972 | | |
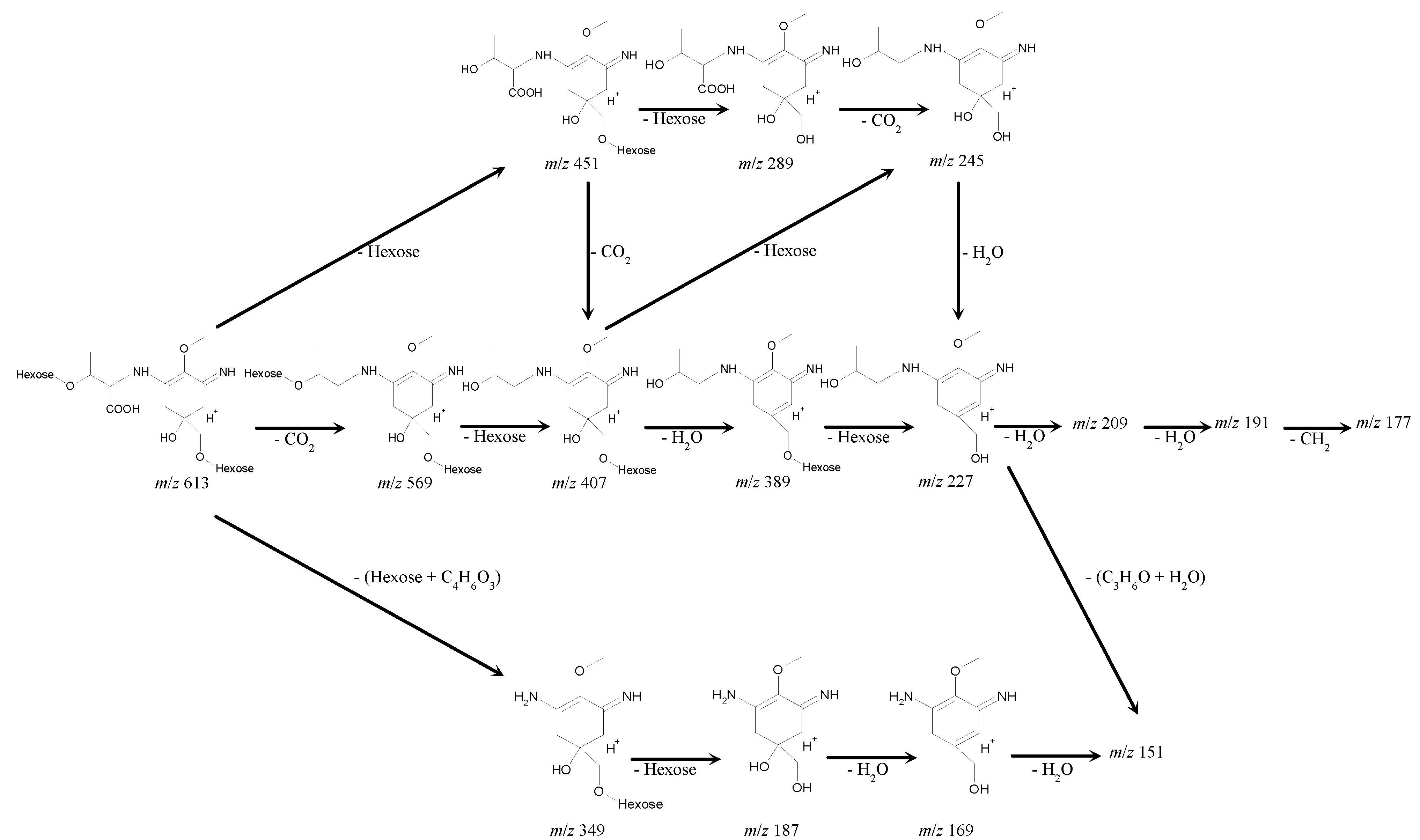
Figure 5.
A proposed fragmentation pattern for the 612-Da MAA, based on the MALDI-TOF MS/MS analysis. MALDI-TOF MS/MS analysis was performed on the parent molecular ion fragment with m/z 613 and the other main molecular ion peaks with m/z 569, 451, 407, 389 and 349. With regard to the mass spectra, a plausible fragmentation pattern shows the presence of a two-hexose and a threonine chain linked by a cyclohexenimine ring.
Figure 5.
A proposed fragmentation pattern for the 612-Da MAA, based on the MALDI-TOF MS/MS analysis. MALDI-TOF MS/MS analysis was performed on the parent molecular ion fragment with m/z 613 and the other main molecular ion peaks with m/z 569, 451, 407, 389 and 349. With regard to the mass spectra, a plausible fragmentation pattern shows the presence of a two-hexose and a threonine chain linked by a cyclohexenimine ring.
The successive deletions of 162 Da from the parental molecular fragment with
m/
z 613 to produce
m/
z 451 and
m/
z 289 indicates the presence of two hexoses (C
6H
10O
5) in the parent molecular ion with
m/
z 613; the resultant fragment with
m/
z 289 can be assumed to be a palythine-threonine (C
12H
20N
2O
6 + H
+). Consistent with this assumption, the deletion of 44 Da from
m/
z 289 to produce
m/
z 245 indicated the deletion of CO
2 and suggested the presence of a carboxyl group in the molecular fragment with
m/
z 289. The fragment with
m/
z 227 could lose two molecules of H
2O and CH
3 to produce the fragments with
m/
z 209, 191 and 177, respectively. The fragment with
m/
z 227 might also lose a part of the threonine chain (C
3H
6O) and H
2O to produce the fragment with
m/
z 170 (or
m/
z 169) and 151 (
Table 7 and
Figure 5).
In a MS/MS analysis of the molecular ion fragment with
m/
z 349, the neutral loss of 162 Da indicated the presence of hexose in the fragment with
m/
z 349, and the fragments with
m/
z 169 and
m/
z 151 suggested the subsequent deletions of two H
2O molecules (
Table 7 and
Figure 5).
These fragmentation patterns were similar to the results from MS analyses of the known MAAs [
42,
43] and confirmed the proposed structure of the 612-Da MAA with the presence of two hexoses and a carboxyl group.
1H,
13C and 2D NMR experiments were performed in D
2O. The
13C and
1H NMR spectra of the 612-Da MAA were compared to those of the known MAAs mycosporine-glycine and palythine-threonine sulfate as well as β-
d-glucose and methyl β-
d-glucose (
Table 9). Signals with identical chemical shifts to 3-aminocyclohexenimine (C1, 2, 3, 4, 5, 6) and threonine (C9, 10, 11, 12) were observed in the 612-Da MAA, similar to those of palythine-threonine sulfate. In mycosporine-glycine the chemical shifts assignable to carbonyl and imine carbon are obviously distinguishable (159.7 ppm and 187.2 ppm in
Table 9, [
44]). However, the chemical shifts corresponding to imine carbon (C3) and amino alkene carbon (C1) in the 612-Da MAA overlapped each other because of the conjugation between them. In palythine-threonine sulfate these chemical shifts assigned to C1 and C3 carbons also overlapped each other (
Table 9, [
45]). Therefore, high-resolution
13C NMR measurement focusing on the X-range at around 160 ppm was performed to separate two distinct signals assignable to imine and amino alkene carbons at C1 and C3 positions in the 612-Da MAA (NMR data shown in
Supporting Information). These two separated signals demonstrated the presence of the 3-aminocyclohexenimine ring in the 612-Da MAA. In the 2D NMR spectra, characteristic correlations assignable to the MAA backbone were detected. The correlations in COSY and HMBC spectra are summarized in
Table 10 and shown in
Figure 6 as red arrows and blue dashed lines.
Table 9.
Summary of the NMR analysis of the 612-Da MAA with an absorption maximum at 322 nm.
Table 9.
Summary of the NMR analysis of the 612-Da MAA with an absorption maximum at 322 nm.
| C | 612-Da MAA with Amax at 322 nm | Palythine-threonine sulfate | Mycosporine-glycine |
|---|
| | 13C | 1H | 13C | 1H | 13C | 1H |
|---|
| 1 a | 163.6 | - | 160.6 | - | 159.7 | - |
| 2 | 127.7 | - | 125.0 | - | 130.4 | - |
| 3 a′ | 163.6 | - | 160.6 | - | 187.2 | - |
| 4 b | 37.0 | 2.85, 3.10 (ABq, 17.6) | 34.0 | 2.81 (ABq, 17.0) | 45.4 | 2.50, 2.73 (ABq, 17.0) |
| 5 | 73.3 | - | 69.8 | - | 72.9 | |
| 6 b′ | 38.9 | 2.76, 3.03 (ABq, 17.0) | 36.0 | 2.77 (ABq, 17.0) | 33.8 | 2.72, 2.83 (ABq, 17.0) |
| 7 | 76.0 | 3.49 (d, 10.0), 3.78 (d, 10.0) | 72.5 | 3.90 (s) | 68.4 | 3.57 (s) |
| 8 | 62.1 | 3.69 (s) | 59.3 | 3.55 (s) | 60.2 | 3.64 (s) |
| 9 | 66.4 | 4.17 (d, 4.6) | 64.5 | 3.98 (d, 4.5) | 43.7 | 4.24 (s) |
| 10 | 177.3 | - | 174.9 | - | 174.5 | - |
| 11 | 75.7 | 4.34 (dq, 4.6, 6.4) | 68.2 | 4.20 (dq, 4.5, 6.0) | | |
| 12 | 18.9 | 1.28 (d, 6.4) | 19.5 | 1.12 (d, 6.0) | | |
| | | | Methyl β-d-Glc | β-d-Glc |
| | | | 13C | 1H | 13C | 1H |
| 1′ | 107.4 | 5.09 (s) | 104.0 | 4.27 | 96.8 | 4.64 |
| 1″ | 101.7 | 5.00 (d, 3.7) | | | | |
Table 10.
Characteristic correlations in COSY and HMBC spectra assignable to the 612-Da MAA backbone.
Table 10.
Characteristic correlations in COSY and HMBC spectra assignable to the 612-Da MAA backbone.
| | COSY | HMBC |
|---|
| H4 | H4 | C2, C3, C5, C6 |
| H6 | H6 | C1, C2, C4, C5 |
| H7 | H7 | C4, C5, C6 |
| H8 | | C2 |
| H9 | H11 | C10 |
| H11 | H9, H12 | C9 |
| H12 | H11 | C9, C11 |
| H1′ | | C11 |
Figure 6.
A predicted structure for the 612-Da MAA with an absorption maximum at 322 nm and a molecular formula of C24H40N2O16. Two hexose sugars are bound to palythine-threonine at C7 and C11 positions. Red arrows and blue dashed lines represent the apparent HMBC and COSY correlations, respectively.
Figure 6.
A predicted structure for the 612-Da MAA with an absorption maximum at 322 nm and a molecular formula of C24H40N2O16. Two hexose sugars are bound to palythine-threonine at C7 and C11 positions. Red arrows and blue dashed lines represent the apparent HMBC and COSY correlations, respectively.
Two separable signals that are characteristic for the anomer positions (C1′ and C1″) of distinguishable carbohydrates were observed (
Table 9), demonstrating the presence of two carbohydrates. The downfield
13C chemical shifts of the carbohydrate anomer positions suggested hexose sugars bound to the palythine-threonine scaffold of the 612-Da MAA. As described in the section on the structure determination of the 508-Da MAA, the downfield
13C shifts of C7 and C11 were thought to be due to hexose binding via
O-glycoside bonds in the 612-Da MAA. According to the HMBC spectrum, a correlation between the C11 carbon in the MAA backbone and the proton bound to the anomer carbon (C1′) of hexose was observed, suggesting the presence of hexose bound at the C11 position. The correlation between the C7 carbon and the anomer proton of the other hexose did not appear; however, geminal coupling at the C7 methylene protons due to the high rotational barrier was observed, suggesting the presence of a large functional group such as a hexose at the C7 position. The difference of
13C chemical shift at C7 position between the 612-Da MAA and palythine-threonine sulfate was only 3.5 ppm, which is smaller than that expected from the difference between the 508-Da MAA and porphyra-334. Because a sulfate group is a strong electron-withdrawing substituent, the
1H-chemical shift at the C7 position in palythine-threonine sulfate shifts downfield from that in palythine-threonine.
These results are consistent with the MS analysis of the 612-Da MAA. After combining the data from the MS and NMR analysis, the predicted structure of the 612-Da MAA was generated and is shown in
Figure 6.
2.5. Radical Scavenging Activity in Glycosylated MAAs
Table 12 shows the radical scavenging activity found in the purified 508-Da and 612-Da MAAs from
N. commune. Both the 508-Da MAA and the 612-Da MAA showed ABTS radical scavenging activity as determined by the decolorization of ABTS radicals. During the time course experiments, the decolorization of the ABTS radicals increased as the incubation time extended from 10 min to 2 h, which suggested that these MAAs were slow-acting radical scavengers. Trolox and ascorbic acid, which were used as standards, are known to be fast-acting scavengers that bring the decolorization reactions to completion within 10 min. Based on these results, the assay incubation time was fixed at 1 h as described by Matsui
et al. [
29]. When ESR was used to directly monitor the decrease in ABTS radical concentrations, the activity of the 612-Da MAA was comparable to that of Trolox and ascorbic acid, while the 508-Da MAA showed weak activity (
Table 12).
Table 12.
Radical scavenging activity in MAAs a.
Table 12.
Radical scavenging activity in MAAs a.
| Assay | Colorimetry b | ESR c |
|---|
| IC50 (mM) |
|---|
| Ascorbic acid d | 0.28 | 0.16 |
| Trolox d | 0.25 | 0.16 |
| 508-Da MAA with Amax at 334 nm | 58 | 29 |
| 612-Da MAA with Amax at 322 nm | 16 | 0.25 |
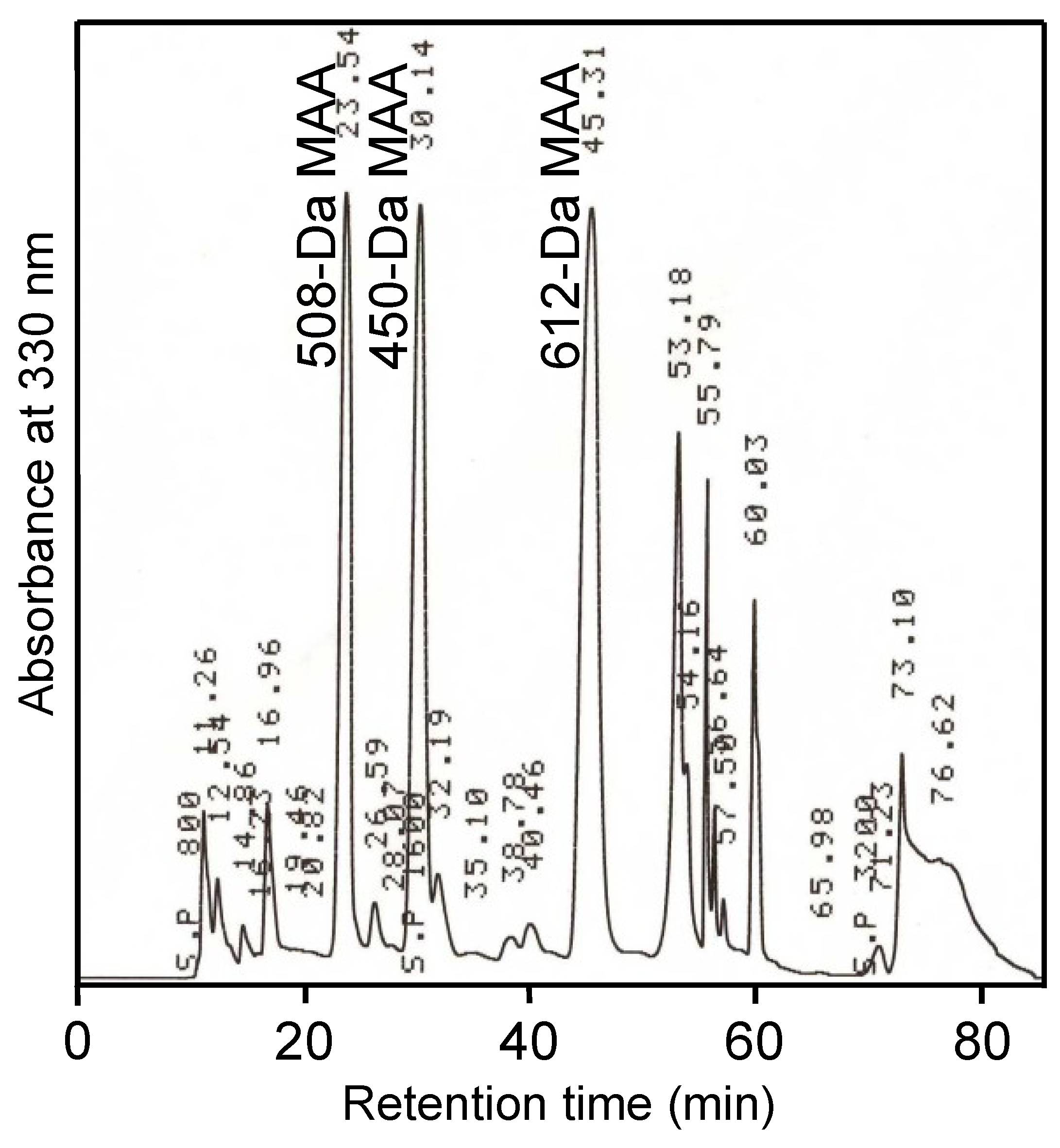
The MAA-associated radical scavenging activity was examined in the
N. commune water extract by HPLC fractionation (
Figure 7a). The fraction with MAA-associated radical scavenging activity accounted for approximately 45% of the total recovered activity (
Figure 7a). Interestingly, high radical scavenging activity, which was not MAA-associated, was detected and determined to account for approximately 40% of the total recovered activity (
Figure 7a). This result suggested that additional water-soluble antioxidants contributed highly to the total radical scavenging capacity of genotype D
N. commune. This non-MAA radical scavenger will be characterized in future studies.
Figure 7.
The elution profile of a water extract from N. commune that was fractionated by HPLC. MAAs were detected at A330 (solid line). The radical scavenging activity (TEAC) was measured by the ABTS decolorization assay (dashed line). (a) The water extract was separated on a reverse phase column as described in the Experimental Section. The MAAs were eluted together with their associated radical scavenging activities in the fraction at 6 min, and additional radical scavenging activity was detected in the fraction at 30 min; (b) The MAA-containing fraction collected at 6 min was additionally separated on a gel filtration column as described in Experimental Section. The 612-Da MAA was eluted together with its associated radical scavenging activity from 17 to 21.5 min in a volume of 2.25 mL.
Figure 7.
The elution profile of a water extract from N. commune that was fractionated by HPLC. MAAs were detected at A330 (solid line). The radical scavenging activity (TEAC) was measured by the ABTS decolorization assay (dashed line). (a) The water extract was separated on a reverse phase column as described in the Experimental Section. The MAAs were eluted together with their associated radical scavenging activities in the fraction at 6 min, and additional radical scavenging activity was detected in the fraction at 30 min; (b) The MAA-containing fraction collected at 6 min was additionally separated on a gel filtration column as described in Experimental Section. The 612-Da MAA was eluted together with its associated radical scavenging activity from 17 to 21.5 min in a volume of 2.25 mL.
The recovered fractions with MAAs were further examined on an HPLC system with a gel filtration column (
Figure 7b). The 612-Da MAA accounted for approximately 60% of the total MAAs, and the radical scavenging activity eluted with the 612-Da MAA accounted for approximately 45% of the recovered activity (
Figure 7b). These results suggest that the 612-Da MAA is a major MAA in genotype D
N. commune and is also a main component of the water-soluble radical scavengers, as it contributed approximately 20% of the total water-soluble radical scavenging activity in the water extract.
2.6. Specificity of the Genotypes and MAAs
N. commune is known to be genetically diverse, and four major genotypes of
N. commune have been reported in Japan; however, the morphological features of the macroscopic colonies and microscopic trichomes are almost identical, and the genotypes are indistinguishable without a determination of molecular taxonomical markers [
33]. During an investigation of MAA contents in field-isolated
N. commune colonies, we observed different UV-absorption spectra in the water extracts from colonies at different sampling locations. We have reported two types of
N. commune, those that specifically produce a 478-Da MAA with an absorption maximum at 335 nm or those that produce a 1050-Da MAA with double absorption maxima at 312 and 340 nm, although biological or physiological differences that would allow us to separate the different MAA producers could not be determined [
29]. In addition to these two types of
N. commune colonies, another type of
N. commune colony, the water extract of which showed a characteristic UV-absorbing spectrum with an absorption maximum at 325 nm was found. According to the unique UV-absorbing spectrum and HPLC chromatogram of the water extract, we thought that this particular
N. commune contained a structurally different MAA. Thus, in this study, the novel glycosylated MAAs were purified and characterized from organisms that did not produce either the 478-Da MAA or the 1050-Da MAA. Eight samples of
N. commune from different sampling locations that had an absorption maximum at 325 nm in their water extracts were identified as genotype D, according to their 16S rRNA nucleotide sequences [
46]; these data confirmed that genotype D is the 612-Da MAA producer
. The glycosylated MAA patterns could be a feasible chemotaxonomic marker with which to characterize
N. commune; it might be classified into three groups, namely, the 478-Da MAA producer, the 1050-Da MAA producer and the 612-Da MAA producer, as these groups are genetically different chemical races of
N. commune. Further investigations into the genotypes and specificities of the types of the glycosylated MAAs are ongoing, and the results will be published in the future.
2.8. Porphyra-334 and Its Derivatives
Porphyra-334, a UV-protective compound with absorption maximum at 334 nm and molecular mass of 346 Da, was first identified in the marine red alga
Porphyra tenera [
51] and was reported to be one of the most common MAAs in marine algal species [
52]. To date, a limited number of studies have been published on the occurrence of porphyra-334 in cyanobacteria. Porphyra-334 has been reported in the marine cyanobacterial species of
Nodularia [
53], in the freshwater bloom-forming cyanobacterium
Microcystis aeruginosa [
54], in the aquatic cyanobacterium
Aphanizomenon flos-aquae [
35] and in the rice-field cyanobacterium
Anabaena doliolum [
55]. We have reported a 478-Da MAA pentose-bound porphyra-334 derivative in the terrestrial cyanobacterium
N. commune [
29], and in this study, we have identified a 508-Da MAA to be a hexose-bound porphyra-334 derivative. These studies have indicated the existence of glycosylated porphyra-334 in
N. commune; however, the non-glycosylated form of porphyra-334 was not observed and the glycosylation of porphyra-334 in
N. commune suggests a unique adaptation for terrestrial environments.
The absorption maximum of the hexose-bound porphyra-334 derivative from
N. commune reversibly shifted to 332 nm in a highly acidic solution, similar to porphyra-334 [
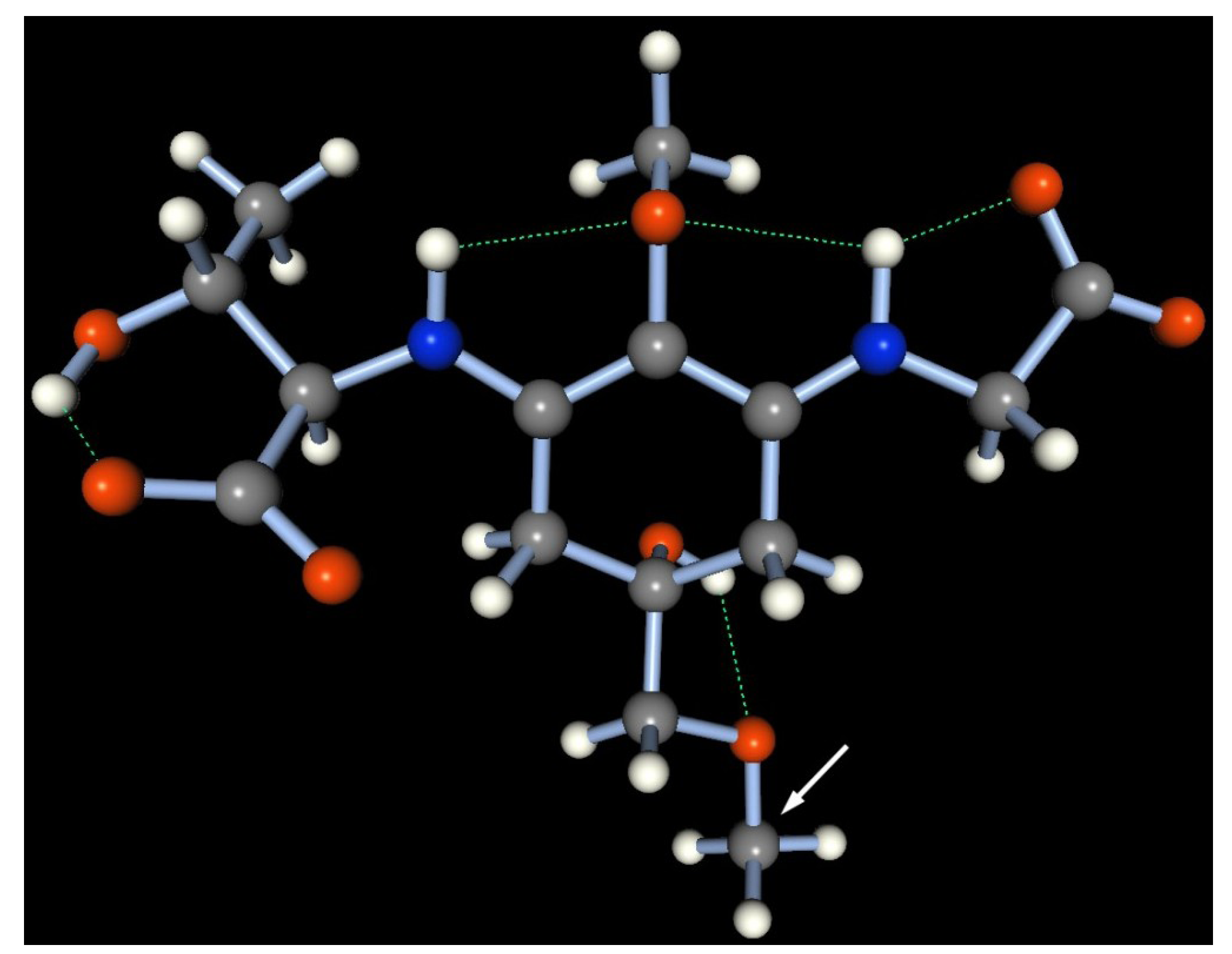
56], which suggested that glycosylated porphyra-334 is a zwitterion in aqueous solution and thus is stable across a wide pH range. To confirm the stable conformation, a semi-empirical molecular simulation of the 508-Da MAA in neutral water at 25 °C was performed by molecular orbital package (MOPAC) 2011 with the PM6 Hamiltonian function (
Figure 8). A methyl group was substituted to mimic the hexose group in the 508-Da MAA and simplify the simulation (indicated by a white arrow in
Figure 8). The most stable form of the methyl-porphyra-334 derivative occupied a near-planar conformation, which allowed for a stable conformation with three hydrogen bonds in the molecule (depicted as green dashed bonds; heat of formation = −485.75 kcal/mol). In acidic conditions, the protonation on the carboxylate anion could cleave the one of the hydrogen bonds and thus would slightly prevent the resonance delocalization of π-electrons within the molecule.
Figure 8.
A stable geometry of methyl porphyra-334 obtained by a molecular simulation of MOPAC 2011, based on the PM6 Hamiltonian function. The hydrogen bond is represented as a green dashed line. Gray: carbon, white: hydrogen, blue: nitrogen, red: oxygen atom. A methyl group (used instead of a hexose group) is indicated by a white arrow.
Figure 8.
A stable geometry of methyl porphyra-334 obtained by a molecular simulation of MOPAC 2011, based on the PM6 Hamiltonian function. The hydrogen bond is represented as a green dashed line. Gray: carbon, white: hydrogen, blue: nitrogen, red: oxygen atom. A methyl group (used instead of a hexose group) is indicated by a white arrow.
The biosynthesis of porphyra-334 is thought to be genetically controlled, and porphyra-334 producers can be classified as a taxonomical group in cyanobacteria and also in marine algae. Biochemical and molecular biological studies of the biosynthesis of the porphyra-334 scaffold based 478-Da and 508-Da MAAs in
N. commune will yield further understanding of the diversity and function of porphyra-334, which is produced and accumulated in taxonomically diverse marine, freshwater and terrestrial organisms [
18,
19,
20].
2.10. Antioxidative Role of MAAs
In terrestrial environments,
N. commune colonies are subjected to desiccation and UV-irradiation. Protective compounds that are involved in reactive oxygen-scavenging mechanisms must have an important role in increasing cell tolerance to the oxidative stresses associated with desiccation and UV irradiation [
12,
59]. MAAs have been suggested to be protective against UV-induced oxidative stress in algae [
60] and in cyanobacteria [
61]. In
N. commune, the 478-Da and 1050-Da MAAs were reported to show radical scavenging activity
in vitro, and the 1050-Da MAA, which has potent radical scavenging activity, contributes approximately 27% of the total water-soluble radical scavenging activity [
29]. The glycosylated MAAs identified in this study also showed radical scavenging activities
in vitro (
Table 12). Although porphyra-334 was reported to have photoprotective but not antioxidant functions [
62,
63,
64], both glycosylated derivatives of porphyra-334, the 508-Da MAA (
Table 12) and the 478-Da MAA [
29], had slow-acting radical scavenging activities
in vitro. The glycosylation of porphyra-334 has been suggested to provide the antioxidant activities of these glycosylated porphyra-334 derivatives, although the molecular mechanism is unknown. The 612-Da MAA was the main water-soluble radical scavenger in the water extract, as it provided approximately 20% of the water-soluble radical scavenging activities in
N. commune. The 612-Da and 1050-Da MAAs were the major MAAs in their respective colonies and demonstrated very similar water-soluble radical scavenging capacities [
29]. The high
in vitro radical scavenging activity of the 1050-Da MAA consisting of cyclohexenone chromophores is consistent with the antioxidative functions of the oxocarbonyl-type MAAs, such as mycosporine-glycine [
64,
65,
66] and mycosporine-taurine [
61]. Because the 612-Da MAA is not an oxocarbonyl-type MAA, the high radical scavenging activity may be due to its glycosylation, similar to the glycosylated porphyra-334 derivatives. These findings suggest the importance of these glycosylated MAAs to the oxidative stress response in
N. commune and support the idea of multifunctional MAAs as UV-protectants and antioxidants.
2.12. Biosynthesis of Glycosylated MAAs in N. commune
More than 30 structurally distinct MAAs, including the mycosporine-derivatives, have been characterized to date [
18,
19,
22,
57,
58]. These MAAs are formed from the precursor 4-deoxygadusol [
70] and are conjugated with a nitrogen substituent (e.g., amino acids) [
18,
19,
22,
71]. The gene product of Ava_3856 from
Anabaena variabilis can convert 4-deoxygadusol and glycine into mycosporine-glycine [
70], which is a metabolic precursor of the bi-substituted MAAs [
70,
72]. Previous reports indicate that condensation of the mono-substituted mycosporine-glycine with an amino acid would be a common reaction in the generation of bi-substituted MAAs such as shinorine and porphyra-334 [
70,
72]. Consistent with a previous report on the coral
P. capitata which suggested that palythine-threonine is formed by decarboxylation of porphyra-334 followed by demethylation of mycosporine-methylamine-threonine [
42], the simultaneous occurrence of palythine-threonine and porphyra-334 as scaffolds in
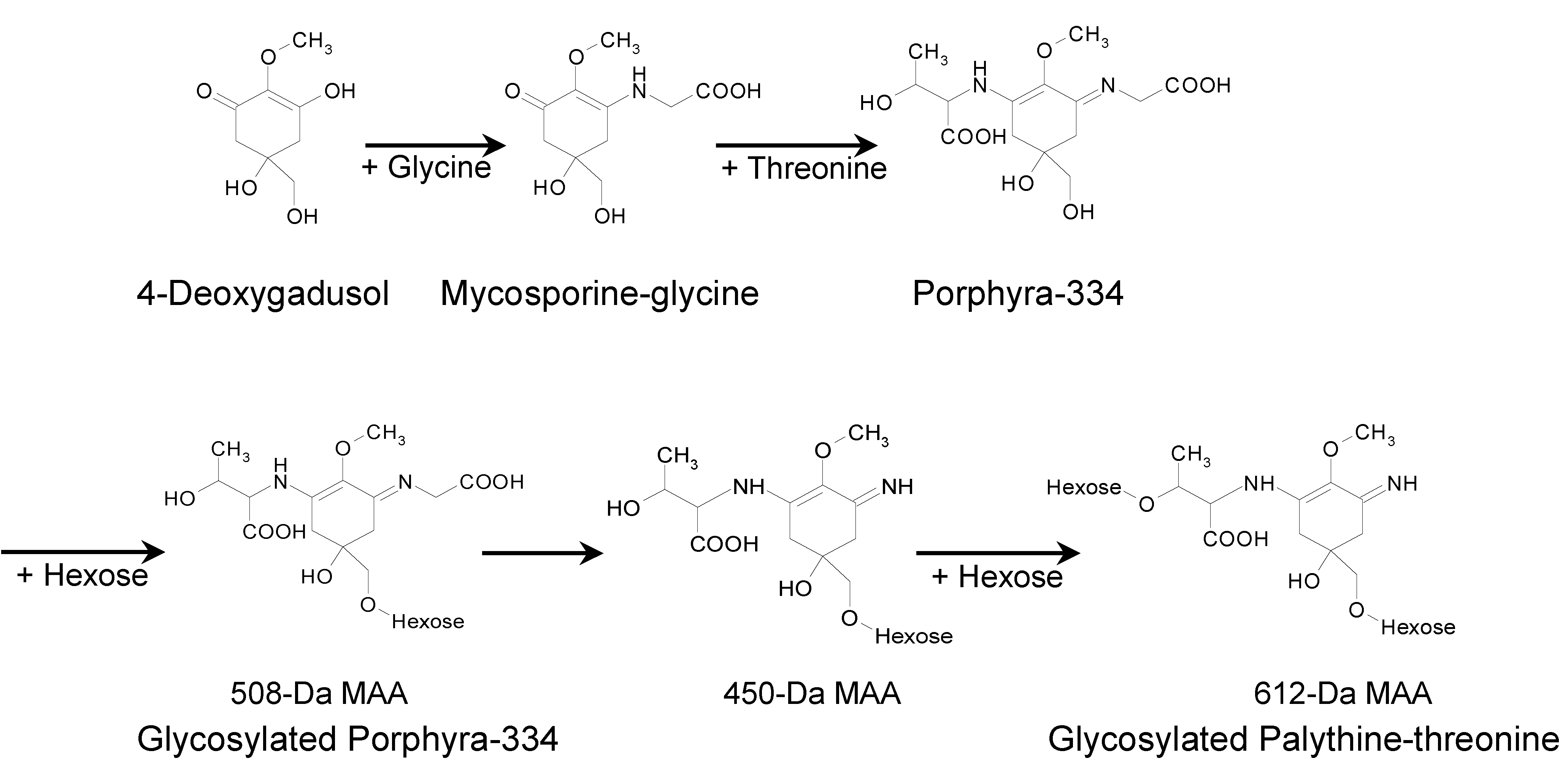
N. commune suggests that porphyra-334 could be a metabolic precursor of palythine-threonine (
Figure 9). Supporting this idea, the glycosylated palythine-threonine accounted for 60% of the total MAAs and was the most abundant one in
N. commune. According to the structures and fragmentation patterns of the glycosylated MAAs, a MAA biosynthetic pathway is predicted in
N. commune (
Figure 9). In our predicted pathway, porphyra-334 could be generated via the addition of threonine to the core ring of mycosporine-glycine and the glycosylation of porphyra-334 could produce the 508-Da MAA. The 450-Da MAA could be formed via the elimination of a portion of glycine from C3 of the 508-Da MAA. The addition of another hexose to the 450-Da MAA could produce the 612-Da MAA (
Figure 9). This biosynthetic pathway for the production of the glycosylated palythine-threonine and porphyra-334 derivatives could occur particularly in genotype D of
N. commune; however, the enzymes and genes involved in the biosynthesis of these glycosylated MAAs are unknown. Further molecular genetic studies are required to identify the enzymes and associated genes in the predicted biosynthetic pathway as well as the function of these glycosylated MAAs in adaptation to terrestrial environments.
Figure 9.
A plausible biosynthetic pathway for glycosylated MAAs in N. commune (genotype D).
Figure 9.
A plausible biosynthetic pathway for glycosylated MAAs in N. commune (genotype D).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







