Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex

 ,
,  ,
, 
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
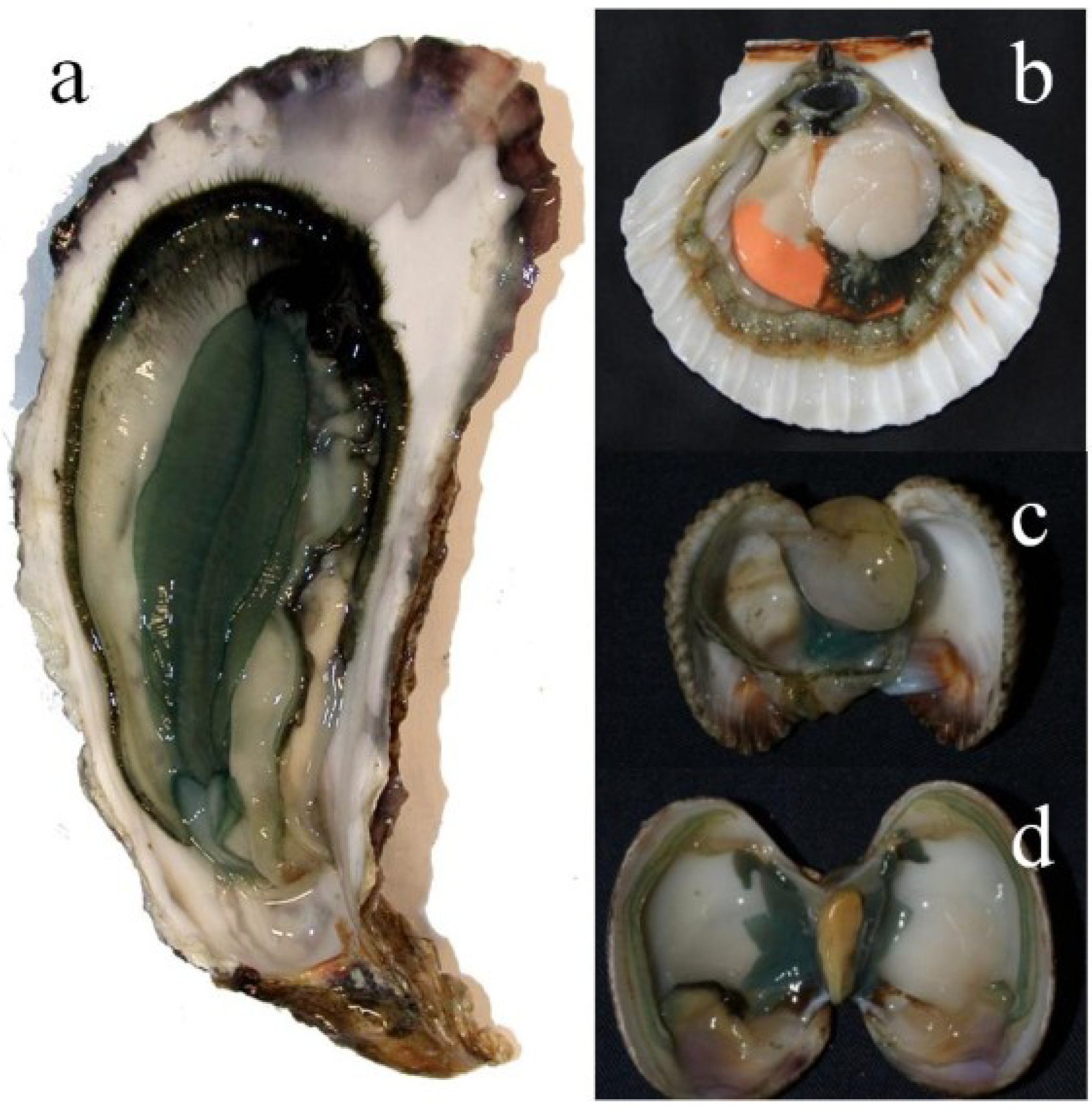
2. The Marennine–Oyster Connection

2.1. A Brief History of the Greening of Oysters, Signature of the Diatom Haslea ostrearia
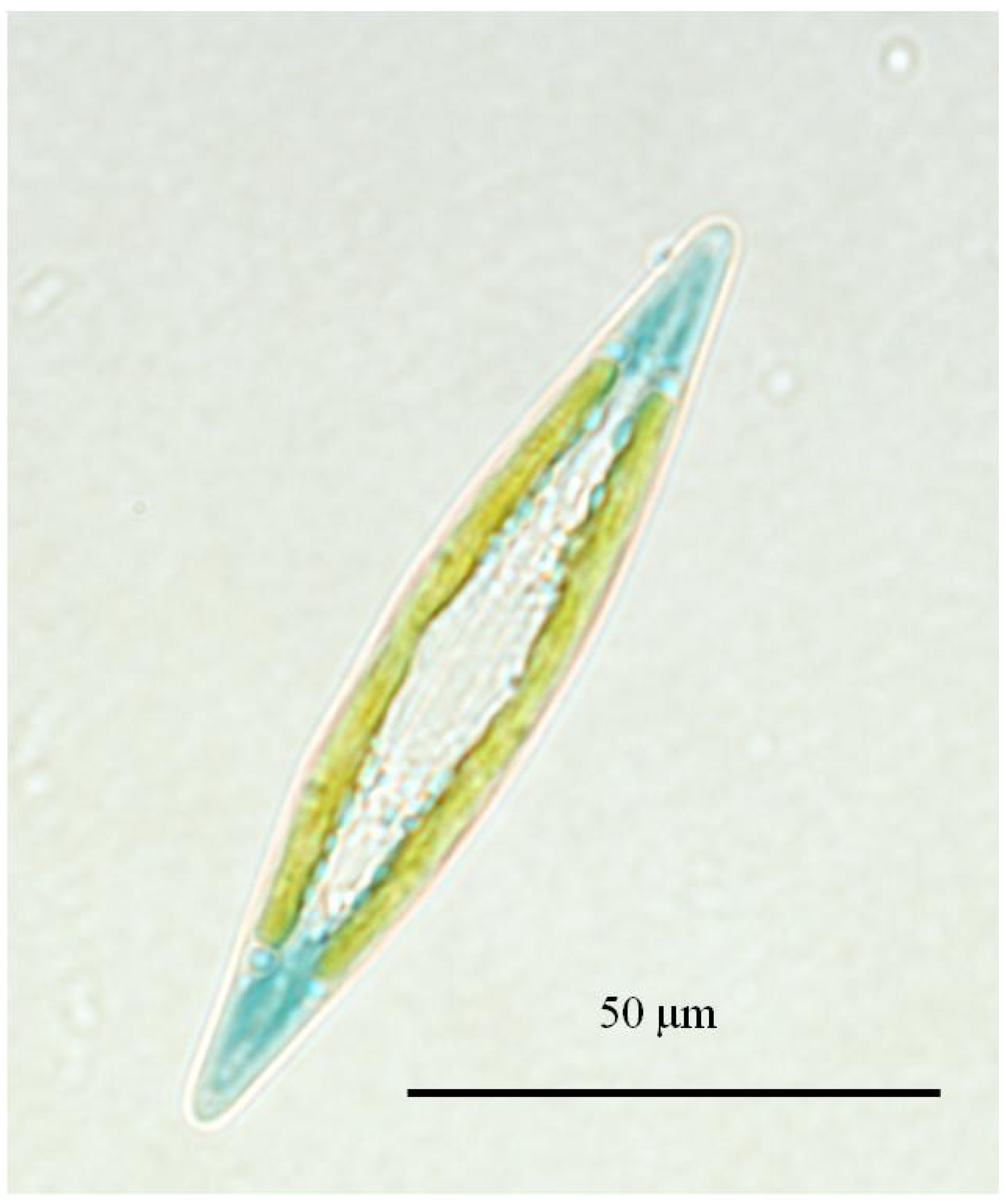
2.2. Haslea ostrearia, a Very Uncommon Diatom

2.3.An Expanding Species—Complex of Blue Diatoms from the Genus Haslea



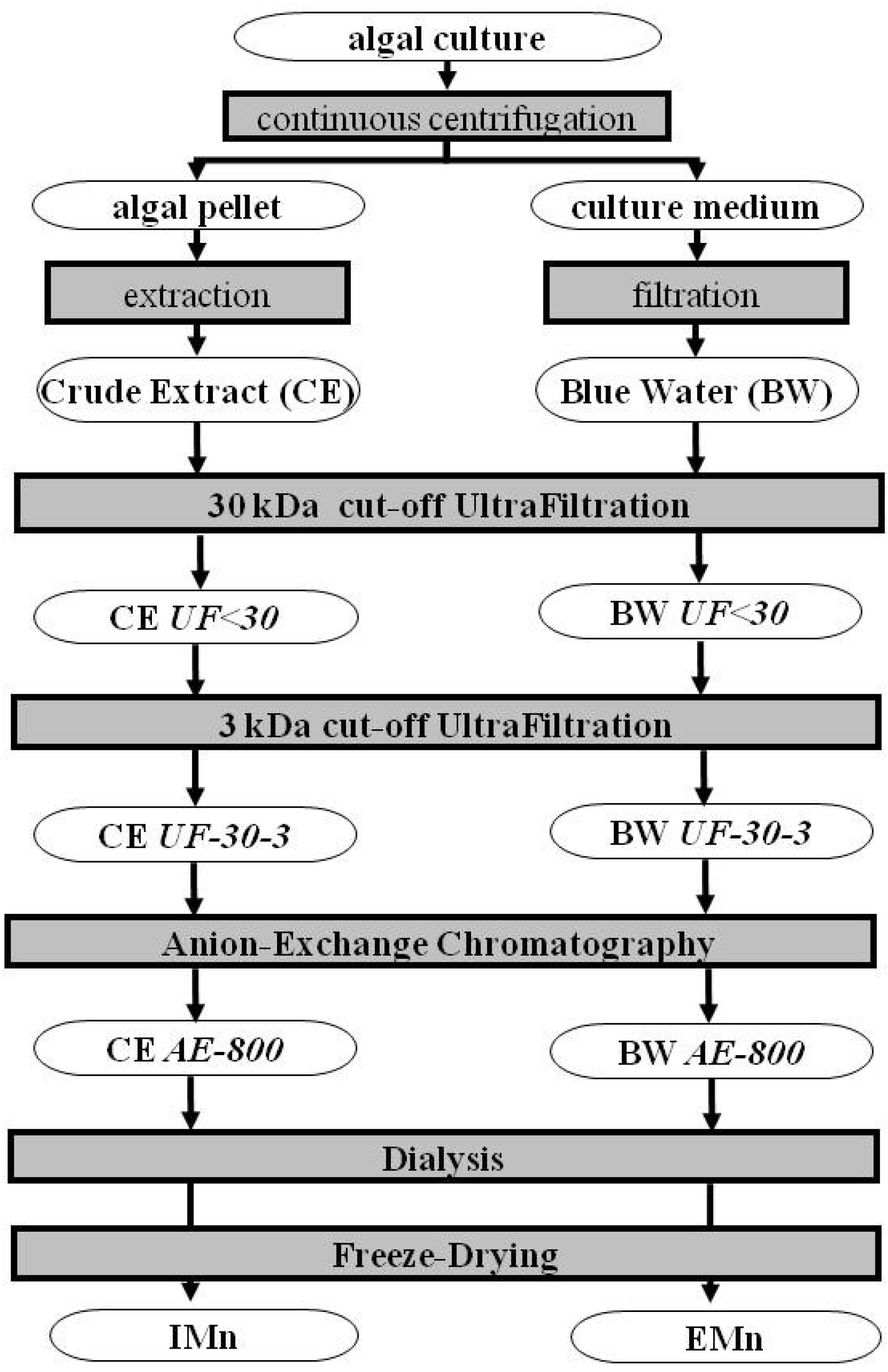
3. Mass Production and Methods of Purification
3.1. Overview of Former Attempts and State of the Art

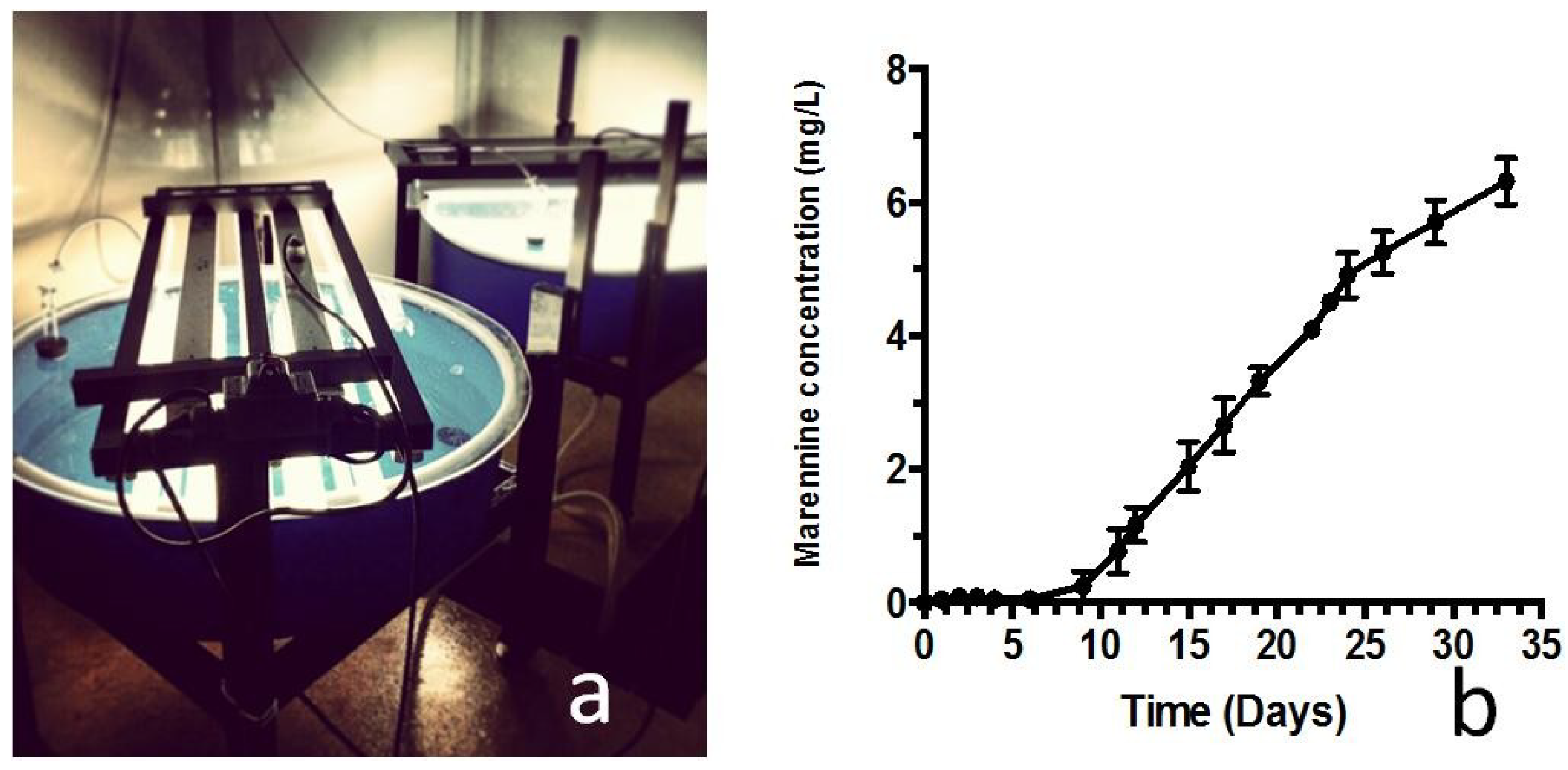
3.2. Recent Transfers to a Pre-Industrial Scale

4. Historical Perspective on Marennine Studies
4.1. Biosynthesis of Marennine: When, Where, Why, and How It Happens
4.2. The Various Hypotheses Regarding the Chemical Nature of Marennine
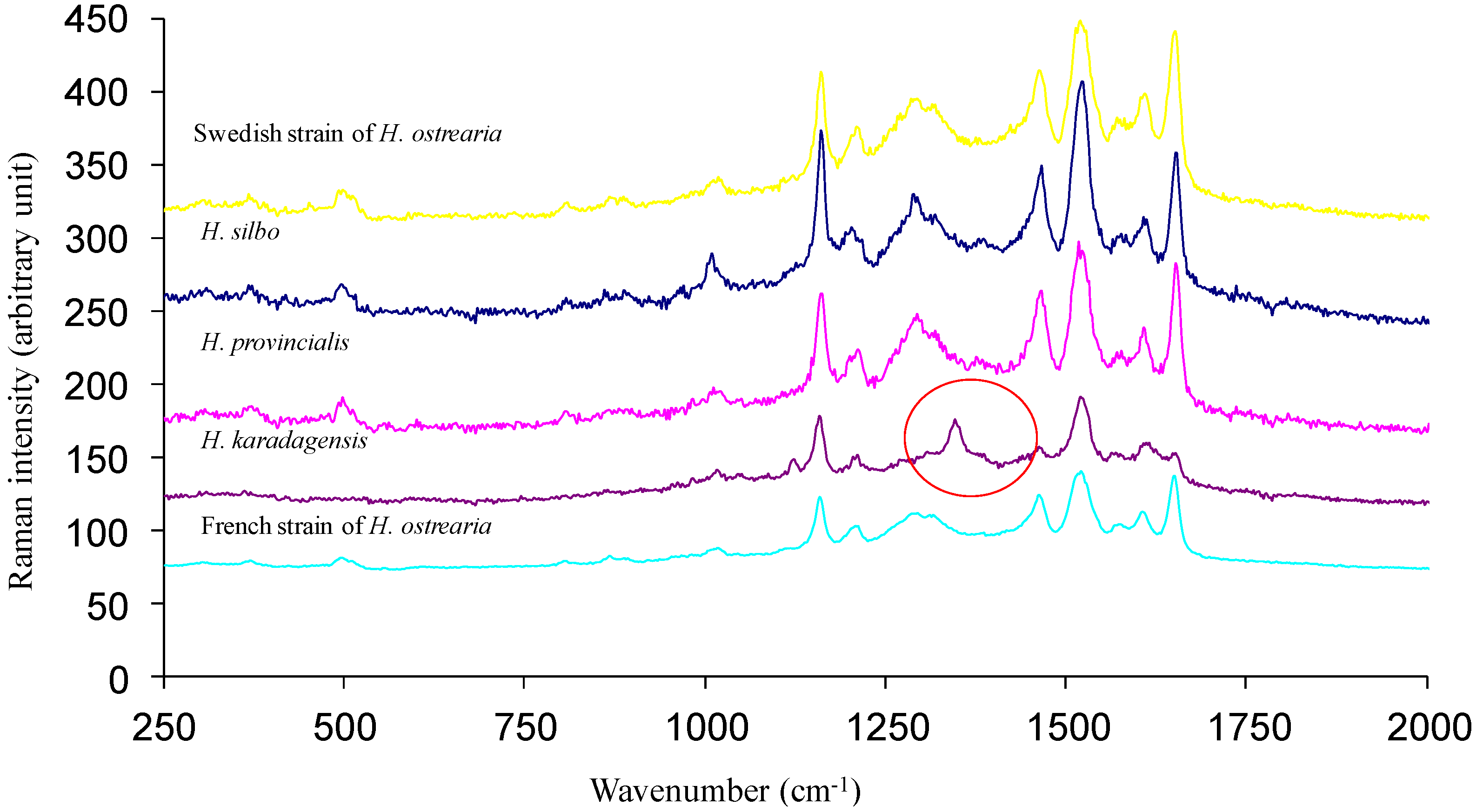
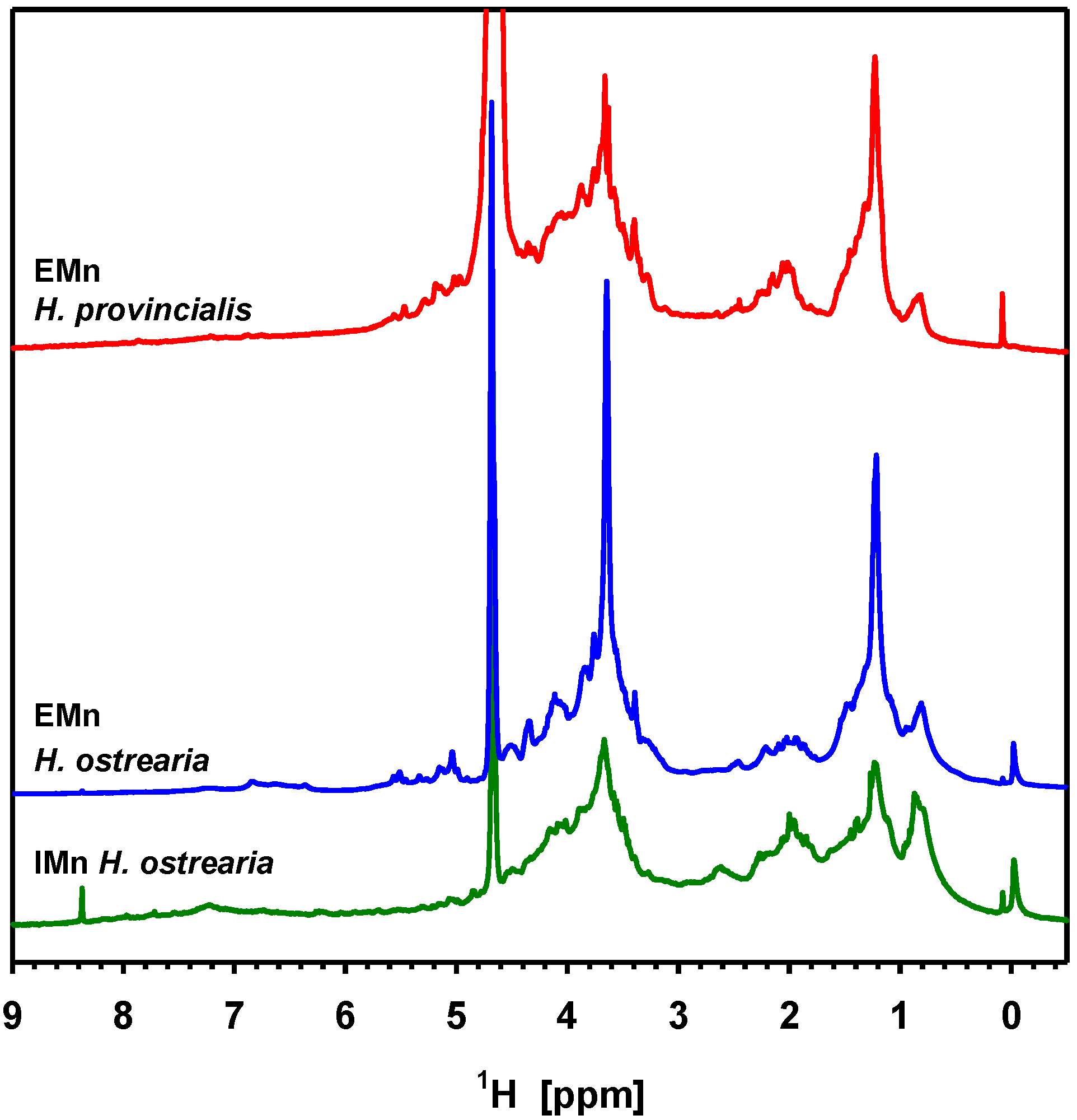
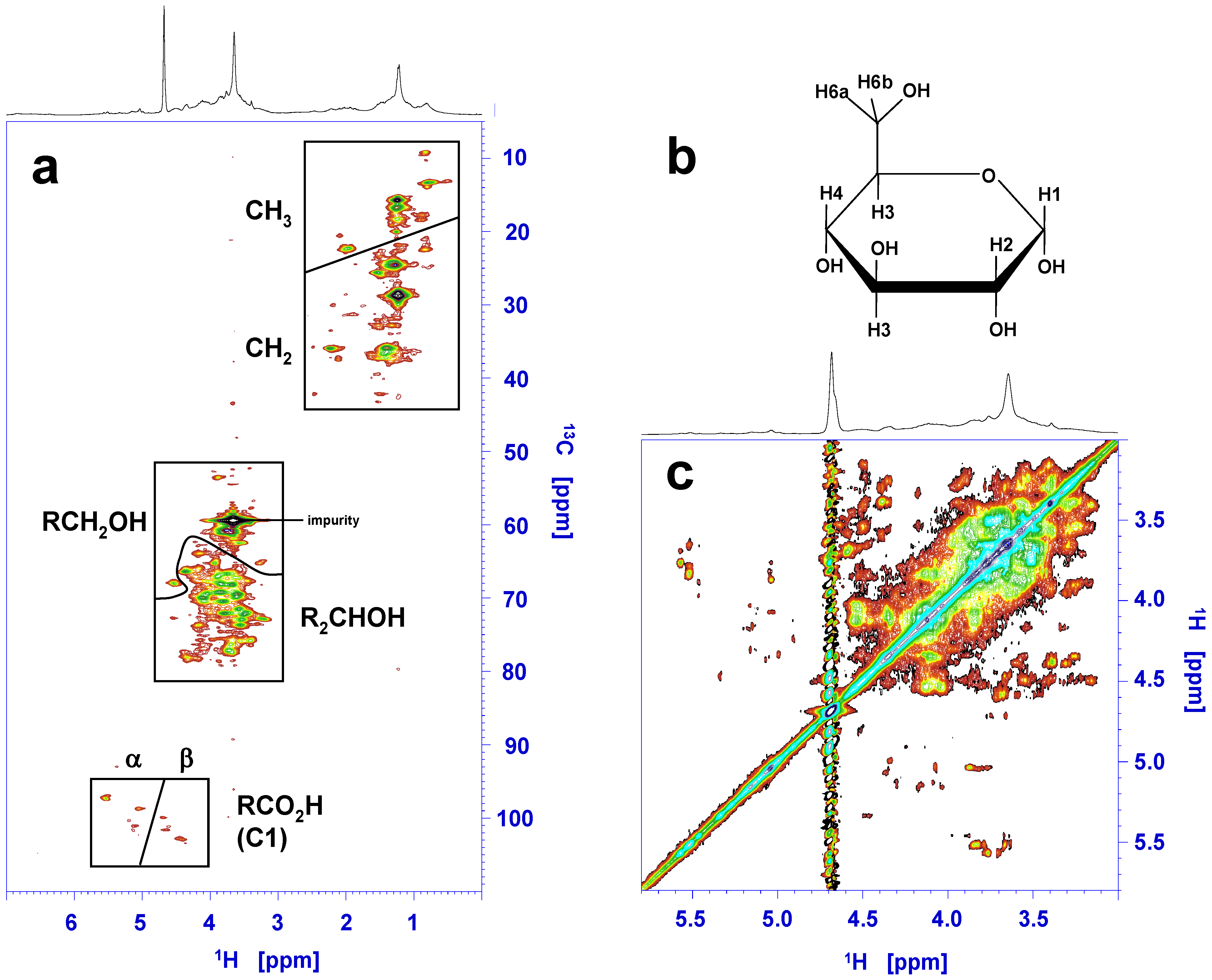
4.3. Recent Discoveries on Marennine’s Structure

5. Marennine Possible Functions and Biological Activities

6. The Putative Commercial Potential of Marennine-Like Pigments
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Wrede, F.; Strack, E. Über das Pyocyanin, den blauen Farbstoff des Bacillus pyocyaneus. Z. Physiol. Chem. 1929, 181, 58–76. [Google Scholar] [CrossRef]
- Frank, L.H.; DeMoss, R.D. On the biosynthesis of pyocyanine. J. Bacteriol. 1959, 77, 776–782. [Google Scholar]
- Hassan, H.M.; Fridovich, I. Mechanism of the antibiotic action of pyocyanine. J. Bacteriol. 1980, 141, 156–163. [Google Scholar]
- Fujikawa, H.; Akimoto, R. New Blue Pigment Produced by Pantoea agglomerans and Its Production Characteristics at Various Temperatures. Appl. Environ. Microbiol. 2011, 77, 172–178. [Google Scholar] [CrossRef]
- Fenical, W. Chemical Studies of Marine Bacteria: Developing a New Resource. Chem. Rev. 1999, 93, 1673–1683. [Google Scholar] [CrossRef]
- Brettar, I.; Christen, R.; Höfle, M.G. Rheinheimera baltica gen. nov., sp. nov., a blue coloured bacterium isolated from the central Baltic Sea. Int. J. Syst. Evol. Microbiol. 2002, 52, 1851–1857. [Google Scholar] [CrossRef]
- Grossart, H.P.; Thorwest, M.; Plitzko, I.; Brinkhoff, T.; Simon, M.; Zeeck, A. Production of a Blue Pigment (Glaukothalin) by Marine Rheinheimera spp. Int. J. Microbiol. 2009, 2009. [Google Scholar] [CrossRef]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the Antimicrobial Secondary Metabolite Indigoidine Contributes to Competitive Surface Colonization by the Marine Roseobacter Phaeobacter sp. Strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef]
- Starr, M.P.; Cosens, G.; Knackmuss, H.J. Formation of the Blue Pigment Indigoidine by Phytopathogenic Erwinia. Appl. Microbiol. 1966, 14, 870–872. [Google Scholar]
- Reverchon, S.; Rouanet, C.; Expert, D.; Nasser, W. Characterization of Indigoidine Biosynthetic Genes in Erwinia chrysanthemi and Role of This Blue Pigment in Pathogenicity. J. Bacteriol. 2002, 184, 654–665. [Google Scholar]
- Tandeau de Marsac, N. Phycobiliproteins and phycobilisomes: The early observations. Photosynth. Res. 2003, 76, 197–205. [Google Scholar] [CrossRef]
- Esenbeck, N. Ueber einen blau-rothen Farbstoff, der sich bei der Zersetzung von Oscillatorien bildet. Liebigs Ann. Chem. (Ann. Pharm.) 1836, 18, 75–82. [Google Scholar]
- Sorby, H.C. On the characteristic colouring-matters of the red groups of algae. J. Linnean Soc. Bot. 1877, 15, 34–40. [Google Scholar] [CrossRef]
- Mölisch, H. Das Phycocyan, ein krystallisirbarer Eiweisskörper. Bot. Z. 1895, 53, 131–135. [Google Scholar]
- Mölisch, H. Das phycoerythrin, seine Krystallisirbarkeit und chemische. Natur. Bot. Z. 1894, 52, 177–189. [Google Scholar]
- Kylin, H. Über die roten und blauen Farbstoffe der Algen. Z. Physiol. Chem. 1912, 76, 396–425. [Google Scholar] [CrossRef]
- Apt, K.E.; Collier, J.L.; Grossman, A.R. Evolution of the Phycobiliproteins. J. Mol. Biol. 1995, 248, 79–96. [Google Scholar] [CrossRef]
- Kai, A.; Yoshii, Y.; Nakayama, T.; Inouye, I. Aurearenophyceae classis nova, a New Class of Heterokontophyta Based on a New Marine Unicellular Alga Aurearenacruciata gen. et sp. nov. Inhabiting Sandy Beaches. Protist 2008, 159, 435–457. [Google Scholar] [CrossRef]
- Robert, J.M.; Morançais, M.; Pradier, E.; Mouget, J.L.; Tremblin, G. Extraction and quantitative analysis of the blue-green pigment “marennine” synthesized by the diatom Haslea ostrearia. J. Appl. Phycol. 2002, 14, 299–305. [Google Scholar] [CrossRef]
- Brill, T.B. The origin of color in organic molecules. In Light: Its Interaction with Art and Antiquities; Plenum Press: New York, NY, USA, 1980; pp. 119–154. [Google Scholar]
- Gastineau, R. Biodiversité, Reproduction et Phylogénie des Diatomées Bleues du Genre Haslea et Valorisation de Leurs Pigments de Type Marennine. Ph.D. Thesis, University of Le Mans, Le Mans, France, September 2011. [Google Scholar]
- Lankester, E.R. On green oysters. Q. J. Microsc. Sci. 1886, 26, 71–94. [Google Scholar]
- Sprat, T. History of the Royal Society; Martyn: London, UK, 1667. [Google Scholar]
- Goubeau de la Bilennerie, J.F. Dissertation sur les huîtres vertes de Marennes, avec des observations critiques de plusieurs naturalistes touchant la reproduction des huîtres en général, et des causes de la couleur verte que ces animaux peuvent acquérir; Goulard: Rochefort, France, 1821. [Google Scholar]
- Pasquier, A. Essai Médical sur les Huîtres; Didot le Jeune: Paris, France, 1818. [Google Scholar]
- Valenciennes, A. Sur les causes de la coloration en vert de certaines huîtres. Comptes Rendus Hebdomadaires des sEances de l'Académie des Sciences 1841, 12, 345–346. [Google Scholar]
- Valmont de Bomare, J.C. Huître in Dictionnaire Raisonné d’Histoire Naturelle; Lacombe: Paris, France, 1768. [Google Scholar]
- Cadet de Gassicourt, C.-L. Cours Gastronomique, ou les Diners de Manant-Ville; Capelle et Renand: Paris, France, 1809. [Google Scholar]
- Bory de Saint-Vincent, J.-B. Dictionnaire Classique d’Histoire Naturelle; Tome Dixième, Rey et Gravier/Baudouin: Paris, France, 1826. [Google Scholar]
- Bosc, G.; Lair, P.A. Huître. In Nouveau Dictionnaire d’Histoire Naturelle Appliquée aux Arts, à l’agriculture, à l’économie Rurale et Domestique, à la Médecine, etc. Tome XV; Deterville: Paris, France, 1817. [Google Scholar]
- Coste, V. Voyage d’Exploration sur le Littoral de la France et de l’Italie; Imprimerie Impériale: Paris, France, 1861. [Google Scholar]
- Bachrach, E. Le bleuissement des Diatomées et le verdissement des huîtres. Revue critique. Revue des Travaux de l'Institut des Pêches Maritimes 1935, 8, 112–123. [Google Scholar]
- Bizio, B. Ricerche sopra il coloramento in verde delle branchie delle ostriche. In Memorie dell’Imperial Regio Instituto Veneto di Scienze Lettere ed Arti; Antonelli: Venice, Italy, 1845; Volume 2. [Google Scholar]
- Boon, D.D. Coloration in bivalves: A Review. J. Food Sci. 1977, 42, 1008–1012. [Google Scholar] [CrossRef]
- Boyce, R.W.; Herdman, W.A. The possible infectivity of the oyster, and upon the green disease in oysters. Comm. Br. Assoc. Adv. Sci. 1896, 66, 663–669. [Google Scholar]
- Mitchell, P.H.; Barney, R.L. The occurrence in Virginia of green-gilled oysters similar to those of Marennes. Bull. U. S. Bur. Fish. 1918, 35, 135–149. [Google Scholar]
- Wang, W.X.; Yang, Y.B.; Guo, X.Y.; He, M.; Guo, F.; Ke, C.H. Copper and zinc contamination in oysters: Subcellular distribution and detoxification. Environ. Toxicol. Chem. 2011, 30, 1767–1774. [Google Scholar] [CrossRef]
- Gayot de Pitaval, F. Bibliothèque des gens de cours, ou Mélange Curieux de bons mots d’Henri IV, de Louis XIV, de Plusieurs Princes et Seigneurs de la Cour, et autres Personnes Illustres Tome V; Le Gras: Paris, France, 1732. [Google Scholar]
- Compain de Saint-Martin, A. Nouvelle Méthode Géographique; Mérigot: Paris, France, 1771; Volume 2. [Google Scholar]
- Prudhomme, L.M. Dictionnaire Géographique Universel Tome IV; Kilian/Picquet: Paris, France, 1828. [Google Scholar]
- Grelon, M. Saintonge, Pays des Huîtres Vertes; Rupella: La Rochelle, France, 1978. [Google Scholar]
- Gaillon, B. Des huîtres vertes, et des causes de cette coloration. J. de Physique, de Chimie, d’histoire Naturelle et des Arts 1820, 91, 222–225. [Google Scholar]
- Bruguière, J.G. Encyclopédie Méthodique: Histoire Naturelle des vers; Chez la veuve Agasse: Paris, France, 1792; Volume 1. [Google Scholar]
- Briée, C. Le verdissement des huîtres: Deux siècles de transformation d’un problème biologique. Ph.D. Thesis, University of Nantes, Nantes, France, October 2010. [Google Scholar]
- Bory de Saint-Vincent, J.B. Navicules. In Dictionnaire Classique d'Histoire Naturelle; Rey et Gravier/Baudouin: Paris, France, 1824; pp. 562–565. [Google Scholar]
- Simonsen, R. The diatom plankton of the Indian Ocean expedition of RV “Meteor”. Hrsg. Dtsch. Forschungsgemeinschaft 1974, 19, 46–49. [Google Scholar]
- Robert, J.M. Fertilité des eaux des claires ostréicoles et verdissement: Utilisation de l’azote par les diatomées dominantes. Ph.D. Thesis, University of Nantes, Nantes, France, April 1983. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms. Biology AndMorphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Neuville, D.; Daste, P. Recherches sur le déterminisme de la production de marennine par la Diatomée marine Navicula ostrearia (Gaillon) Bory en culture in vitro. Rev. Gen. Bot. 1978, 85, 255–303. [Google Scholar]
- Wraige, E.J.; Belt, S.T.; Lewis, C.A.; Cooke, D.A.; Robert, J.M.; Massé, G.; Rowland, S.J. Variations in structures and distributions of C25 highly branched isoprenoid (HBI) alkenes in cultures of the diatom, Haslea ostrearia (Simonsen). Org. Geochem. 1997, 27, 497–505. [Google Scholar] [CrossRef]
- Mouget, J.L.; Tremblin, G.; Morant-Manceau, A.; Morançais, M.; Robert, J.M. Long-term photoacclimation of Haslea ostrearia (Bacillariophyta): Effect of irradiance ongrowth rates, pigment content and photosynthesis. Eur. J. Phycol. 1999, 34, 109–115. [Google Scholar] [CrossRef]
- Maestrini, S.Y.; Robert, J.-M. La production micro-algale des claires à huîtres: Particularités nutritionnelles; importance de l’azote organique dissous. Production et relations trophiques dans les écosystèmes marins. IFREMER Act. Coll. 1984, 5, 185–214. [Google Scholar]
- Petersen, C.G.J. “Grüne Austern” in Dänemark in 1911–1912. Int. Rev. Hydrobiol. Hydrogr. 1916, 7, 39–41. [Google Scholar] [CrossRef]
- Ryder, J.A. On the cause of the greening of oysters. With a supplementary note on the coloration of the blood corpuscles of the oysters. In Report of Commission of Fish and Fisheries for 1882; Governement Printing Office: Washington, DC, USA, 1884. [Google Scholar]
- Ranson, G. L’absorption de matières organiques dissoutes par la surface extérieure du corps chez les animaux aquatiques. Annales de l'Institut Océanographique 1927, 4, 174. [Google Scholar]
- Gastineau, R.; Davidovich, N.A.; Bardeau, J.F.; Caruso, A.; Leignel, V.; Hardivillier, Y.; Jacquette, B.; Davidovich, O.I.; Rincé, Y.; Gaudin, P.; et al. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the Black Sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar]
- Gastineau, R.; Hardivillier, H.; Leignel, V.; Tekaya, N.; Morançais, M.; Fleurence, J.; Davidovich, N.A.; Jacquette, B.; Gaudin, P.; Hellio, C.; et al. Greening effect on oysters and biological activities of the blue pigments produced by the diatom Haslea karadagensis (Naviculaceae). Aquaculture 2013, 368–369, 61–67. [Google Scholar]
- Grünow, A. New diatoms from Honduras. Month. Microscop. J. 1877, 18, 165–186. [Google Scholar] [CrossRef]
- Davidovich, N.A.; Gastineau, R.; Gaudin, P.; Davidovich, O.I.; Mouget, J.L. Sexual reproduction in the second described blue diatom, Haslea karadagensis. Fottea 2012, 12, 219–229. [Google Scholar]
- Gastineau, R.; Pouvreau, J.B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Bourgougnon, N.; Mouget, J.L. Biological activities of purified marennine, the blue pigment produced by the diatom Haslea ostrearia and responsible for the greening of oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef]
- Vandanjon, L. Etude d’un procédé de valorisation d’une microalgue marine: Concentration et purification par techniques à membranes d’un pigment naturel produit par la diatomée Haslea ostrearia. Ph.D. Thesis, University of Nantes, Nantes, France, June 1997. [Google Scholar]
- Rossignol, N. Procédés d’extraction et de séparation par membranes appliqués à la production du pigment endo- et exocellulaire synthétisé par la diatomée Haslea ostrearia Simonsen. Mise en oeuvre d’un photobioréacteur à membrane à fonctionnement continu. Ph.D. Thesis, University of Nantes, Nantes, France, November 1999. [Google Scholar]
- Vandanjon, L.; Jaouen, P.; Rossignol, N.; Quéméneur, F.; Robert, J.-M. Concentration and desalting by membrane processes of a natural pigment produced by the marine diatom Haslea ostrearia Simonsen. J. Biotechnol. 1999, 70, 393–402. [Google Scholar] [CrossRef]
- Rossignol, N.; Moan, R.; Jaouen, P.; Robert, J.-M.; Quéméneur, F. Continuous high-pressure disruption of marine diatom Haslea ostrearia. Assessment by laser diffraction particle sizer. Biotechnol. Technol. 1999, 13, 909–913. [Google Scholar] [CrossRef]
- Rossignol, N.; Jaouen, P.; Robert, J.-M.; Quéméneur, F. Production of exocellular pigment by the marine diatom Haslea ostrearia Simonsen in a photobioreactor equipped with immersed ultrafiltration membranes. Bioresour. Technol. 2000, 73, 197–200. [Google Scholar] [CrossRef]
- Rossignol, N.; Lebeau, T.; Jaouen, P.; Robert, J.-M. Comparison of two membrane-photobioreactors, with free or immobilized cells, for the production of pigments by a marine diatom. Bioprocess Eng. 2000, 23, 495–501. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Morançais, M.; Massé, G.; Rosa, P.; Robert, J.-M.; Florence, J.; Pondaven, P. Purification of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 769–781. [Google Scholar] [CrossRef]
- Turpin, V.; Robert, J.; Goulletquer, P. Limiting nutrients of oyster pond seawaters in the Marennes-Oléron region for Haslea ostrearia: Applications to the mass production of the diatom in mesocosm experiments. Aquat. Living Resour. 1999, 12, 335–342. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press Inc.: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Pouvreau, J.-B.; Morançais, M.; Fleury, F.; Rosa, P.; Thion, L.; Cahingt, B.; Zal, F.; Fleurence, J.; Pondaven, P. Preliminary characterization of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 757–767. [Google Scholar] [CrossRef]
- Lebeau, T.; Junter, G.A.; Jouenne, T.; Robert, J.-M. Marennine production by agar-entrapped Haslea ostrearia Simonsen. Bioresour. Technol. 1999, 67, 13–17. [Google Scholar] [CrossRef]
- Lebeau, T.; Gaudin, P.; Junter, G.A.; Mignot, L.; Robert, J.-M. Continuous marennine production by agar-entrapped Haslea ostrearia using a tubular photobioreactor with internal illumination. Appl. Microbiol. Biotechnol. 2000, 54, 634–640. [Google Scholar] [CrossRef]
- Lebeau, T.; Gaudin, P.; Moan, R.; Robert, J.-M. A new photobioreactor for continuous marennine production with a marine diatom: Influence of the light intensity and the immobilised-cell matrix (alginate beads or agar layer). Appl. Microbiol. Biotechnol. 2002, 59, 153–159. [Google Scholar] [CrossRef]
- Rosa, P.; Turpin, V.; Robert, J.; Goulletquer, P.; Masse, G.; Charente, A.B.; Bay, A. Oyster greening by outdoor mass culture of the diatom Haslea ostrearia Simonsen in enriched seawater. Aquac. Res. 2001, 32, 801–809. [Google Scholar] [CrossRef]
- Robert, J.M.; Hallet, J.N. Absorption Spectrum in vivo of the blue pigment “marennine” of the pennate diatom Navicula ostrearia Bory. J. Exp. Bot. 1981, 32, 341–345. [Google Scholar] [CrossRef]
- Tremblin, G.; Cannuel, R.; Mouget, J.L.; Rech, M.; Robert, J.M. Change in light quality due to a blue-green pigment, marennine, released in oyster-ponds: Effect on growth and photosynthesis in two diatoms, Haslea ostrearia and Skeletonema costatum. J. Appl. Phycol. 2000, 12, 557–566. [Google Scholar] [CrossRef]
- Mouget, J.L.; Rosa, P.; Vachoux, C.; Tremblin, G. Enhancement of marennine production by blue light in the diatom Haslea ostrearia. J. Appl. Phycol. 2005, 17, 437–445. [Google Scholar] [CrossRef]
- Moreau, J. Contribution aux recherches écologiques sur les claires à huîtres du bassin de Marennes-Oléron. Rev. Trav. Inst. Pêches. Marit. 1970, 34, 380–462. [Google Scholar]
- Ranson, G. Le verdissement des huîtres. Sciences 1937, 8, 13–24. [Google Scholar]
- Bachrach, E.; Jouvent, A. Sur la pigmentation bleue de certaines diatomées. Bulletin Mensuel de la Société Linnéenne de Lyon 1933, 2, 36–37. [Google Scholar]
- Bachrach, E.; Simonet, M. Le phénomène du bleuissement chez les diatomées. Revue des Travaux de l’Institut des Pêches Maritimes 1936, 9, 113–116. [Google Scholar]
- Bornet, E.; Chatin, A.D. Le verdissement des huîtres. Bulletin des Séances de laSociété Royale et Centrale d’agriculture 1895, 55, 429–439. [Google Scholar]
- Nassiri, Y.; Robert, J.M.; Rincé, Y.; Ginsburger-Vogel, T. The cytoplasmic finestructure of the diatom Haslea ostrearia (Bacillariophyceae) in relation to marennine production. Phycologia 1998, 37, 84–91. [Google Scholar] [CrossRef]
- Moreau, J. Recherches préliminaires sur le verdissement en claires: L’évolution de leurs divers pigments liée au complexe pigmentaire de Navicula ostrearia Bory. Rev. Trav. Inst. Pêches Marit. 1967, 31, 372–382. [Google Scholar]
- Genevès, L.; Choussy, M.; Barbier, M.; Neuville, D.; Daste, P. Ultrastructure et composition pigmentaire comparées des chromatophores de la Diatomée Navicula ostrearia (Gaillon) Bory normale et bleue. C. R. Acad. Sci. Sér. D Sci. Nat. 1976, 282, 449–452. [Google Scholar]
- Tremblin, G.; Robert, J.M. Comportement photosynthétique de Haslea ostrearia en relation avec sa pigmentation bleue. C. R. Acad. Sci. Ser. III Sci. de la Vie 1996, 319, 939–944. [Google Scholar]
- Tremblin, G.; Robert, J.M. Carbon fixation by the peculiar marine diatom Haslea ostrearia. Photosynthetica 2001, 39, 215–220. [Google Scholar] [CrossRef]
- Sauvageau, C. A propos de la présence de la diatomée bleue dans la Méditerranée. Travaux des Lab. (Arcachon) 1906, 6, 46–59. [Google Scholar]
- Sauvageau, C. Le verdissement des huîtres par la diatomée bleue. Travaux des Lab. (Arcachon ) 1907, 2, 1–128. [Google Scholar]
- Neuville, D.; Daste, P. Observations concernant la production de pigment bleu par la Diatomée Navicula ostrearia (Gaillon) Bory, maintenue en culture unialgale. C. R. Acad. Sci. Sér. D Sci. Nat. 1970, 271, 2389–2391. [Google Scholar]
- Neuville, D.; Daste, P. Variations structurales présentées par la Diatomée Navicula ostrearia (Gaillon) Bory, soumise à diverses conditions de culture. Comptes Rendus de la Société de Biol. 1971, 165, 1723–1726. [Google Scholar]
- Robert, J.-M. Etude préliminaire des relations entre l’état de verdissement d’une claire ostréicole et l’examen biométrique de populations planctoniques et benthiques du Navicula ostrearia Bory, pigmenté; applications. C. R. Acad. Sci. Sér. D Sci. Nat. 1974, 278, 1405–1408. [Google Scholar]
- Robert, J.-M. Le verdissement des huîtres dans les claires de la baie de Bourgneuf. Bull. de la Soc. des Sci. Nat. de l’Ouest de la France 1975, 73, 123–129. [Google Scholar]
- Sullivan, W.K. Report on the composition of the soils of oyster grounds; and on qualities which exert most influence on oyster cultivation. In Appendix H, Report of the commission appointed to inquire into the methods of oyster culture in the United Kingdom and France, with a view to the introduction of improved methods of cultivation of oysters in Ireland; Alexander Thom: Dublin, Ireland, 1870. [Google Scholar]
- Bocat, L. Sur la marennine de la diatomée bleue; comparaison avec la phycocyanine. Comptes Rendus des Seances de la Société de Biologie et de ses Filiales 1907, 62, 1073–1075. [Google Scholar]
- Neuville, D.; Daste, P. Production de pigment bleu par la Diatomée Navicula ostrearia (Gaillon) Bory, maintenue en culture unialgale sur un milieu synthétique carencé enazote nitrique. C. R. Acad. Sci. Sér. D Sci. Nat. 1972, 274, 2030–2033. [Google Scholar]
- Hardouin, V.; Vandanjon, L.; Jaouen, P.; Robert, J.-M. Procédé combiné extraction-membranes pour l’isolement et la purification de pigments naturels. Actes Colloque Interfiltra Intermembr. 1994, 175–181. [Google Scholar]
- Bubb, W.A. NMR spectroscopy in the study of carbohydrates: Characterizing the structural complexity. Concept Magnetic Res. A 2003, 19, 1–19. [Google Scholar] [CrossRef]
- Størseth, T.R.; Kirkvold, S.; Skjermo, J.; Reitan, K.I. A branched β-d-(1→3,1→6)-glucan from the marine diatom Chaetoceros debilis (Bacillariophyceae) characterized by NMR. Carbohydr. Res. 2006, 341, 2108–2134. [Google Scholar] [CrossRef]
- Dyer, W.T.T. Greening of oysters. Nature 1877, 16, 397. [Google Scholar] [CrossRef]
- Puységur, M. Notice sur la cause du verdissement des huîtres. Revue Maritime et Coloniale 1880, 64, 248–256. [Google Scholar]
- Schubert, H.; Tremblin, G.; Robert, J.M.; Sagert, S.; Rincé, Y. In-vivo fluorescence measurement of photosynthesis of Haslea ostrearia Simonsen in relation to marennine content. Diatom Res. 1995, 10, 341–349. [Google Scholar] [CrossRef]
- Pouvreau, J.B.; Morançais, M.; Taran, F.; Rosa, P.; Dufossé, L.; Guérard, F.; Pin, S.; Fleurence, J.; Pondaven, P. Antioxidant and free radical scavenging properties of marennine, a blue-green polyphenolic pigment from the diatom Haslea ostrearia (Gaillon/Bory) Simonsen responsible for the natural greening of cultured oysters. J. Agric. Food Chem. 2008, 56, 6278–6286. [Google Scholar] [CrossRef]
- Minier, C.; Tutundjian, R.; Galgani, F.; Robert, J.M. Copper tolerance in Haslea ostrearia assessed by measurements of in vivo esterase activity. Mar. Environ. Res. 1998, 46, 579–582. [Google Scholar] [CrossRef]
- Pouvreau, J.B.; Housson, E.; le Tallec, L.; Morançais, M.; Rincé, Y.; Fleurence, J.; Pondaven, P. Growth inhibition of several marine diatom species induced by the shading effect and allelopathic activity of marennine, a blue-green polyphenolic pigment of the diatom Hasleaostrearia (Gaillon/Bory) Simonsen. J. Exp. Mar. Biol. Ecol. 2007, 352, 212–225. [Google Scholar] [CrossRef]
- Ianora, A.; Bentley, M.G.; Caldwell, G.S.; Casotti, R.; Cembella, A.D.; Engström-Öst, J.; Halsband, C.; Sonnenschein, E.; Legrand, C.; Llewellyn, C.A.; et al. The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field. Mar. Drugs 2011, 9, 1625–1648. [Google Scholar] [CrossRef] [Green Version]
- Bergé, J.P.; Bourgougnon, N.; Alban, S.; Pojer, F.; Billaudel, S.; Chermann, J.C.; Robert, J.M.; Franz, G. Antiviral and anticoagulant activities of a water-soluble fraction of the marine diatom Haslea ostrearia. Planta Medica 1999, 65, 604–609. [Google Scholar] [CrossRef]
- Carbonnelle, D.; Pondaven, P.; Morançais, M.; Massé, G.; Bosch, S.; Jacquot, C.; Briand, G.; Robert, J.M.; Roussakis, C. Antitumor and antiproliferative effects of an aqueous extract from the marine diatom Haslea ostrearia (Simonsen) against solid tumors: Lung carcinoma (NSCLC-N6), kidney carcinoma (E39) and melanoma (M96) cell lines. Anticancer Res. 1999, 19, 621–624. [Google Scholar]
- Garnier, M.; Labreuche, Y.; Nicolas, J.L. Molecular and phenotypiccharacterization of Vibrio aestuarianus subsp. francensis subsp. nov., a pathogen of the oyster Crassostrea gigas. Syst. Appl. Microbiol. 2008, 31, 358–365. [Google Scholar] [CrossRef]
- Denis, S.; de Decker, S.; Philippe, H.; Laetitia, C.; Maeva, R.; Garcia, C. A Large-Scale Epidemiological Study to Identify Bacteria Pathogenic to Pacific Oyster Crassostrea gigas and Correlation Between Virulence and Metalloprotease-like Activity. Microb. Ecol. 2010, 59, 787–798. [Google Scholar] [CrossRef]
- Segarra, A.; Pépin, J.F.; Arzul, I.; Morga, B.; Faury, N.; Renault, T. Detection and description of a particular ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010, 153, 92–99. [Google Scholar]
- Tardy-Laporte, C.; Arnold, A.A.; Génard, B.; Gastineau, R.; Morançais, M.; Mouget, J.L.; Tremblay, R.; Marcotte, I. A 2H solid-state NMR study of the effect of antimicrobials on intact non-mutated Escherichia coli. BBA-Biomembranes 2013, 1828, 614–622. [Google Scholar]
- Mayer, A.M.S.; Hamann, M.T. Marine pharmacology in 2001–2002 marine compounds with anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, antiinflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C 2005, 140, 265–286. [Google Scholar]
- Olicard, C.; Renault, T.; Torhy, C.; Benmansour, A.; Bourgougnon, N. Putative antiviral activity in haemolymph from adult Pacific oysters. Crassostrea gigas. Antivir. Res. 2005, 66, 147–152. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar]
- Shimizu, Y. Drugs from the Sea; Fusetani, N., Ed.; Karger: Basel, Switzerland, 2000; pp. 30–45. [Google Scholar]
- Scholz, B.; Liebezeit, G. Screening for biological activities and toxicological effects of 63 phytoplankton species isolated from freshwater, marine and brackish water habitats. Harmful Algae 2012, 20, 58–70. [Google Scholar] [CrossRef]
- Yasuhara-Bell, J.; Lu, Y. Marine compounds and their antiviral activities. Antivir. Res. 2010, 86, 231–240. [Google Scholar] [CrossRef]
- Newsome, A.G.; Murphy, B.T.; van Breemen, R.B. Isolation and Characterization of Natural Blue Pigments from Underexplored Sources Physical Methods in Food Analysis; American Chemical Society: Washington, DC, USA, 2013; pp. 105–125. [Google Scholar]
- Kuddus, M.; Singh, P.; Thomas, G.; Al-Hazimi, A. Recent Developments in Production and Biotechnological Applications of C-Phycocyanin. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Couteau, C.; Pommier, M.; Paparis, E.; Coiffard, L.J.M. Study of the efficacy of 18 sun filters authorized in European Union tested in vitro. Pharmazie 2007, 62, 449–452. [Google Scholar]
- Groves, G.A.; Agin, P.P.; Sayre, R.M. In vitro and in vivo methods to define sunscreen protection. Australas. J. Dermatol. 1979, 20, 112–119. [Google Scholar] [CrossRef]
- Couteau, C.; Chammas, R.; Alami, S.; Choquenet, B.; Paparis, E.; Coiffard, L.J.M. Combination of UVA-filters and UVB-filters or inorganic UV-filters—Influence on the Sun Protection Factor (SPF) and the PF-UVA determined by in vitro method. J. Dermatol. Sci. 2008, 50, 159–161. [Google Scholar]
- Brétéché, A.; Duflos, M.; Dassonville, A.; Nourrisson, M.R.; Brelet, J.; le Baut, G.; Grimaud, N.; Petit, J.Y. New N-pyridinyl(methyl)-indole-2- and 3-(Alkyl)carboxamides and Derivatives Acting as systemic and topical Inflammation Inhibitors. J. Enzym. Inhib. Med. Chem. 2002, 17, 415–424. [Google Scholar] [CrossRef]
- Carlson, R.P.; O’Neil-Davis, L.; Chang, J.; Lewis, A.J. Modulation of mouse ear oedema by clyclooxygenase and lipooxygenase inhibitors and other pharmacologic agents. Agents Actions 1985, 17, 197–204. [Google Scholar] [CrossRef]
- Collin, X.; Robert, J.M.; Duflos, M.; Wielgosz, G.; le Baut, G.; Bobin-Dubigeon, C.; Grimaud, N.; Lang, F.; Petit, J.Y. Synthesis of N-Pyridinyl(methyl)-1,2-dihydro-4-hydroxy-2-oxoquinoline-3-carboxamides and analogues and their anti-inflammatory activity in mice and rats. J. Pharm. Pharmacol. 2001, 53, 417–423. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.S.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Mar. Drugs 2014, 12, 3161-3189. https://doi.org/10.3390/md12063161
Gastineau R, Turcotte F, Pouvreau J-B, Morançais M, Fleurence J, Windarto E, Prasetiya FS, Arsad S, Jaouen P, Babin M, et al. Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Marine Drugs. 2014; 12(6):3161-3189. https://doi.org/10.3390/md12063161
Chicago/Turabian StyleGastineau, Romain, François Turcotte, Jean-Bernard Pouvreau, Michèle Morançais, Joël Fleurence, Eko Windarto, Fiddy Semba Prasetiya, Sulastri Arsad, Pascal Jaouen, Mathieu Babin, and et al. 2014. "Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex" Marine Drugs 12, no. 6: 3161-3189. https://doi.org/10.3390/md12063161
APA StyleGastineau, R., Turcotte, F., Pouvreau, J. -B., Morançais, M., Fleurence, J., Windarto, E., Prasetiya, F. S., Arsad, S., Jaouen, P., Babin, M., Coiffard, L., Couteau, C., Bardeau, J. -F., Jacquette, B., Leignel, V., Hardivillier, Y., Marcotte, I., Bourgougnon, N., Tremblay, R., ... Mouget, J. -L. (2014). Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Marine Drugs, 12(6), 3161-3189. https://doi.org/10.3390/md12063161