2.1. Differentiation of Pigment Cells in a Blastula-Derived Cell Culture
The growth patterns and morphology of cultured cells is often determined by peculiarities of the culture medium. To evaluate the effect of different culture media on the development of pigment differentiation in the cell cultures of both sea urchin species, we tested three types of media (
Figure 2 and
Figure 3): seawater, the coelomic fluid preparations of control sea urchins and injured sea urchins. Injured sea urchins were obtained by needle pricks in the area of Aristotle’s lantern (see the
Experimental Section). The first distinctions in the appearance of pigment cells and their pigmentation became obvious after 3–7 days of cultivation. The cells cultivated in seawater were faintly pigmented and not numerous, whereas the pigment cells were more abundant in the coelomic fluid-cultivated cells. This picture was observed both in a short-time culture of the sea urchin
S. mirabilis and in a long-time culture of the sea urchin
S. intermedius.
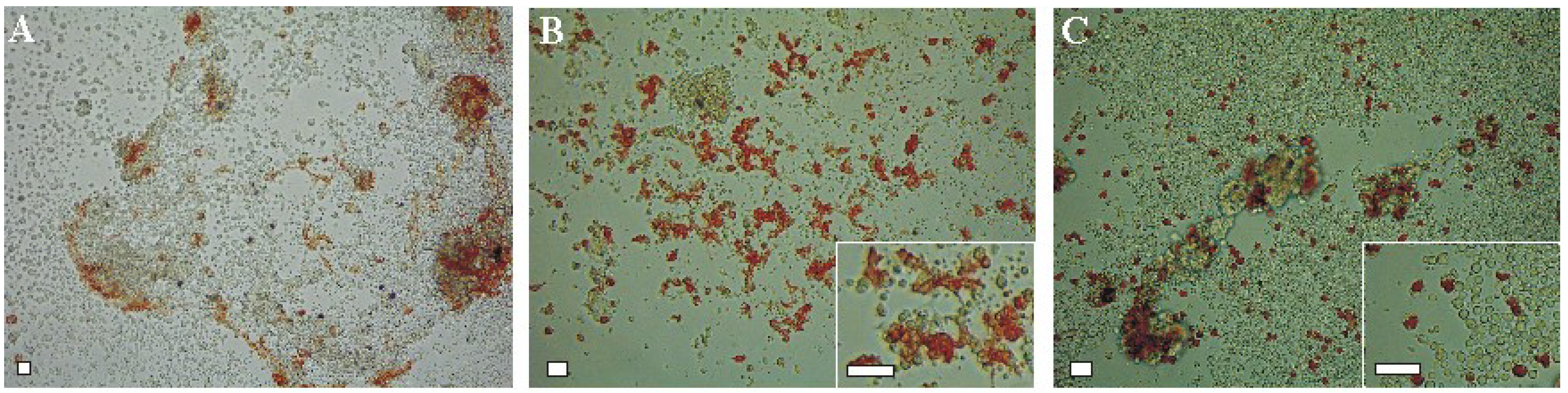
Figure 2.
Embryonic pigment cells in a blastula-derived cell culture of the sea urchin Scaphechinus mirabilis cultivated for 3 days. The cells were cultivated in seawater (A); the coelomic fluid of intact sea urchins (B); or the coelomic fluid of injured sea urchins (C). All culture media were supplemented with 2% fetal bovine serum. Insets in B and C: higher magnifications. Note the change in morphology of the pigment cells depending on the medium tested. Scale bar, 10 μm.
Figure 2.
Embryonic pigment cells in a blastula-derived cell culture of the sea urchin Scaphechinus mirabilis cultivated for 3 days. The cells were cultivated in seawater (A); the coelomic fluid of intact sea urchins (B); or the coelomic fluid of injured sea urchins (C). All culture media were supplemented with 2% fetal bovine serum. Insets in B and C: higher magnifications. Note the change in morphology of the pigment cells depending on the medium tested. Scale bar, 10 μm.
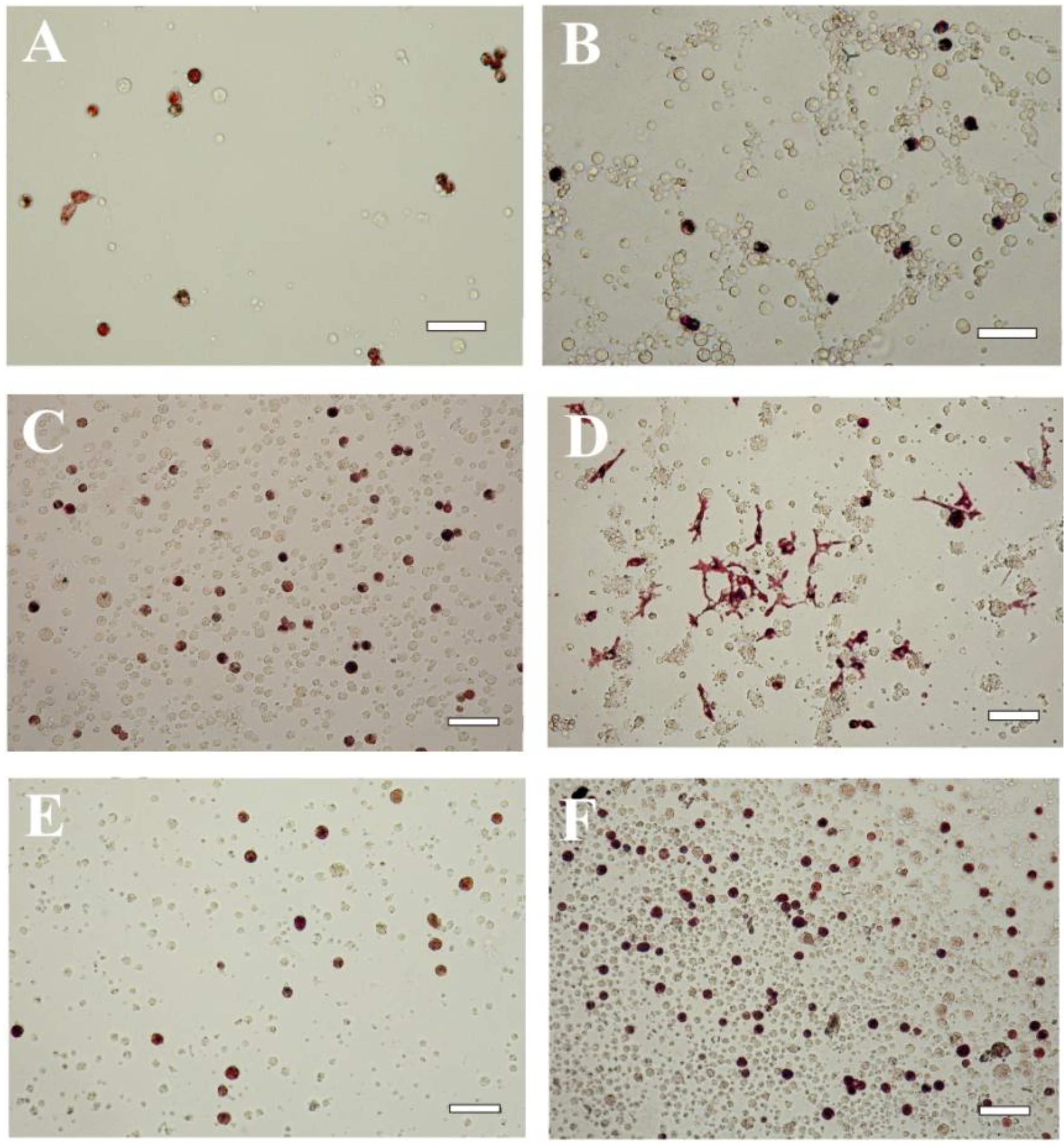
Figure 3.
Embryonic pigment cells in a blastula-derived cell culture of the sea urchin Strongylocentrotus intermedius cultivated for 5 (A,B); 17 (C,D) or 41 days (E,F). A, C, E—cells cultivated in the coelomic fluid of intact sea urchins; B, D, F—cells cultivated in the coelomic fluid of injured sea urchins. The coelomic fluid was supplemented with 2% fetal bovine serum. Note the change in morphology of the pigment cells during cultivation. Scale bar, 10 μm.
Figure 3.
Embryonic pigment cells in a blastula-derived cell culture of the sea urchin Strongylocentrotus intermedius cultivated for 5 (A,B); 17 (C,D) or 41 days (E,F). A, C, E—cells cultivated in the coelomic fluid of intact sea urchins; B, D, F—cells cultivated in the coelomic fluid of injured sea urchins. The coelomic fluid was supplemented with 2% fetal bovine serum. Note the change in morphology of the pigment cells during cultivation. Scale bar, 10 μm.
If the coelomic fluid of intact sea urchins was used as the medium, the pigment cells of
S. mirabilis were well attached and spread during three days in culture (
Figure 2B, insert). However, when the injured sea urchin coelomic fluid was used, most of the pigment cells were rounded and failed to spread (
Figure 2C, insert).
The medium-dependent differences in cell morphology were also seen in the sea urchin
S. intermedius. The pigment differentiation of the cells in the seawater occurred slowly and less extensively (data not shown) than that of the cells in the coelomic fluids. Furthermore, the pigment cells in this species changed their morphology during prolonged cultivation for 20 days in the same medium (
Figure 3A–D). When the cells were cultivated in injured sea urchin coelomic fluid, the maximal spreading of the pigment cells was observed on Day 17 of cultivation (
Figure 3D). Upon further culture, the cells became rounded (
Figure 3E,F) and increased in number up to Day 41 of cultivation (
Figure 4). Signs of cell degeneration were still detectable after two months of culturing.
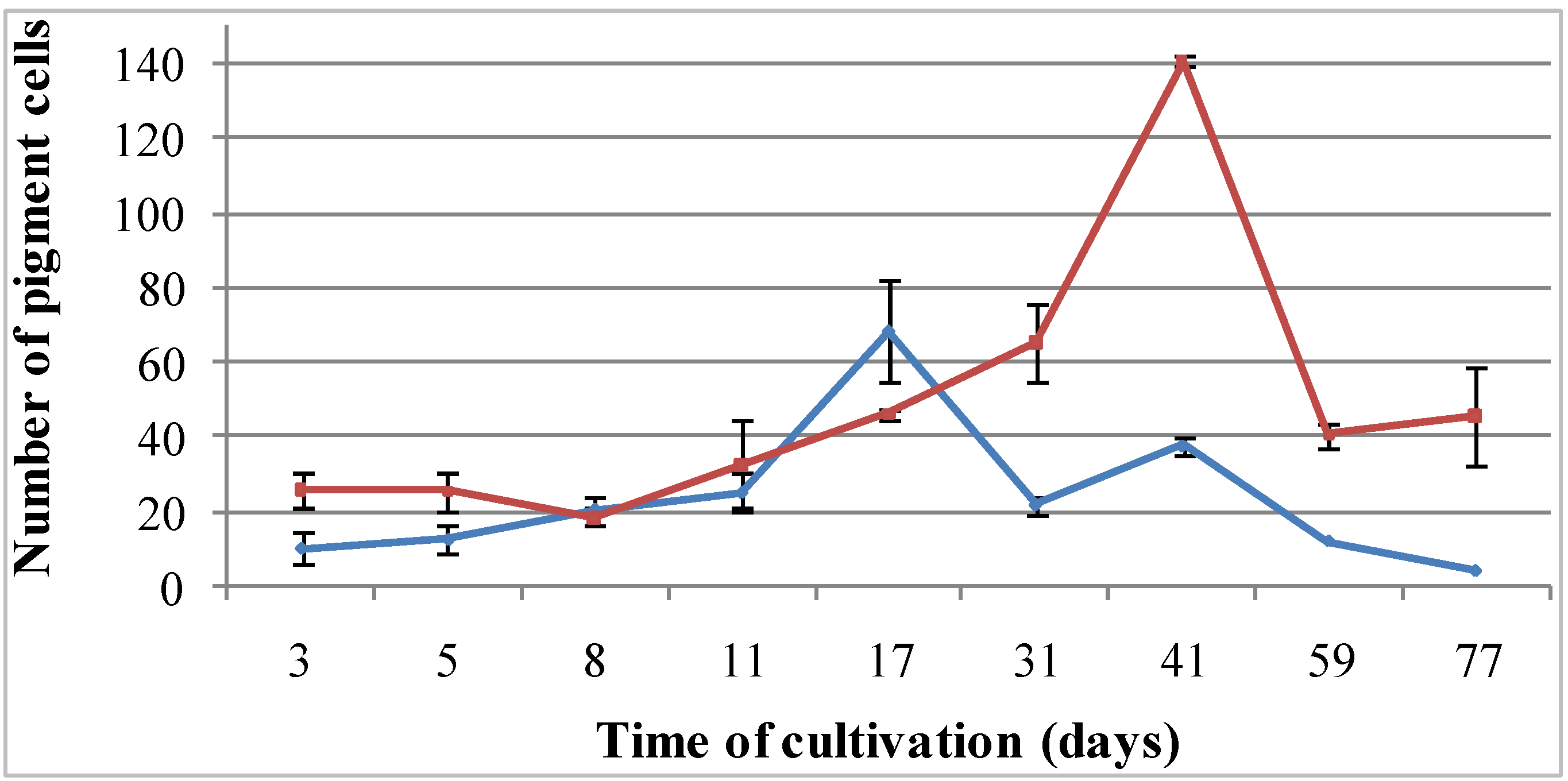
Figure 4.
Cellular dynamics of pigment cells cultivated in coelomic fluids of intact (blue line) and injured (red line) sea urchins for 2.5 months. At least 500 cells were counted in each tested culture medium.
Figure 4.
Cellular dynamics of pigment cells cultivated in coelomic fluids of intact (blue line) and injured (red line) sea urchins for 2.5 months. At least 500 cells were counted in each tested culture medium.
The maximal number of pigmented cells of
S. intermedius was detected on Day 17, when the cells were cultivated in the coelomic fluid of intact sea urchins (
Figure 4, blue line). When the cells were cultivated in the injured sea urchin coelomic fluid, that maximal number was recorded on Day 41 (
Figure 4, red line).
The number of pigment cells was dependent on the coelomic fluid used: twice as many pigment cells were detected in the cells cultivated in the injured sea urchin coelomic fluid as in that from intact sea urchins. In contrast, the number of pigment cells was significantly lower among cells cultured in in seawater (data not shown).
2.2. Cell Proliferation in Culture
It is well known that cell proliferation and differentiation are interrelated processes. To compare the contribution of these processes—pigment differentiation (as described above) and cell proliferation—in sea urchin culture, we detected dividing cells with antibodies to phospho-H3-histone, a well-known cell proliferation marker for many organisms [
17,
18,
19]. At least 300–500 DAPI-stained cells were examined for each experiment with the culture media tested, and the proportion of cells positive for phospho-H3-histone relative to the total number of cells was determined. The number of dividing cells was higher in cells cultured in seawater than in those cultured in the coelomic fluids, where cell division was weak (
Figure 5).
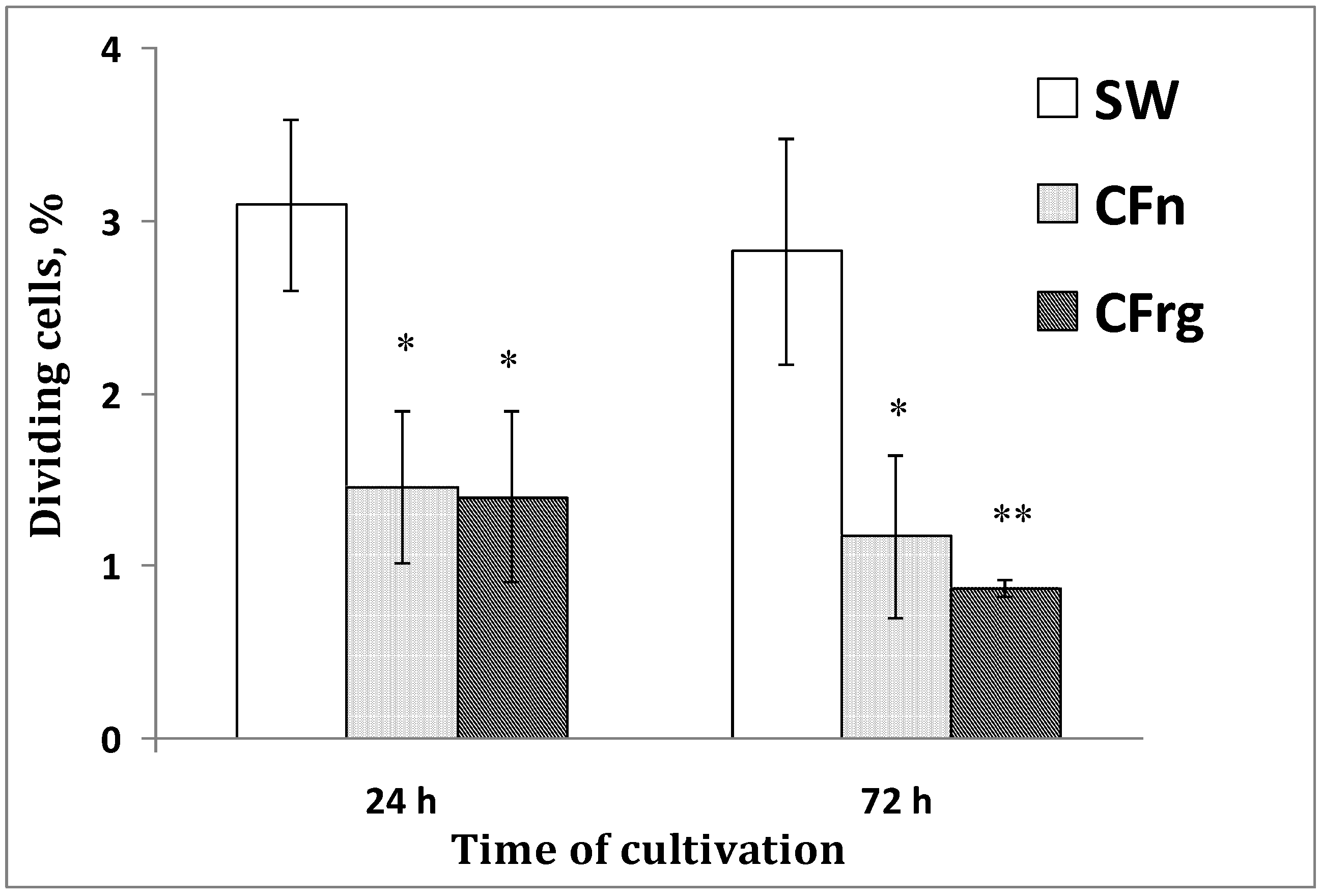
Figure 5.
The number of dividing cells in a blastula-derived culture of the sea urchin S. intermedius depending upon culture medium tested. The cells were cultivated for 24–72 h. The proportion of phospho-H3-histone-positive cells among the total number of examined cells (determined by DAPI staining of nuclei) was counted. SW—seawater; CFn—coelomic fluid obtained from intact sea urchins; CFreg—coelomic fluid obtained from injured sea urchins. * p < 0.05; ** p < 0.01 versus values of dividing cells cultivated in the sea water (24 h).
Figure 5.
The number of dividing cells in a blastula-derived culture of the sea urchin S. intermedius depending upon culture medium tested. The cells were cultivated for 24–72 h. The proportion of phospho-H3-histone-positive cells among the total number of examined cells (determined by DAPI staining of nuclei) was counted. SW—seawater; CFn—coelomic fluid obtained from intact sea urchins; CFreg—coelomic fluid obtained from injured sea urchins. * p < 0.05; ** p < 0.01 versus values of dividing cells cultivated in the sea water (24 h).
A significant difference in the number of dividing cells in the different media after one day of cultivation was found, and this difference increased for three days, when a general decrease in mitotic activity was then detected. In most cases, the cultures survived well, but their proliferation rate was low.
Dividing (phospho-H3-histone+) cells were detected from 6 to 72 h in all media tested, but the maximal number of dividing cells was found in seawater. The decrease in cell proliferation in the coelomic fluids was accompanied by a simultaneous intensification of the pigment cell differentiation. As already noted, the composition of the culture medium affected the rate of appearance of the pigment cells during cultivation.
Figure 6A,B arrows, shows dividing cells in both of the coelomic fluid and seawater after 12 h of cultivation (see details of the immunocytochemical manipulations in the
Experimental Section).
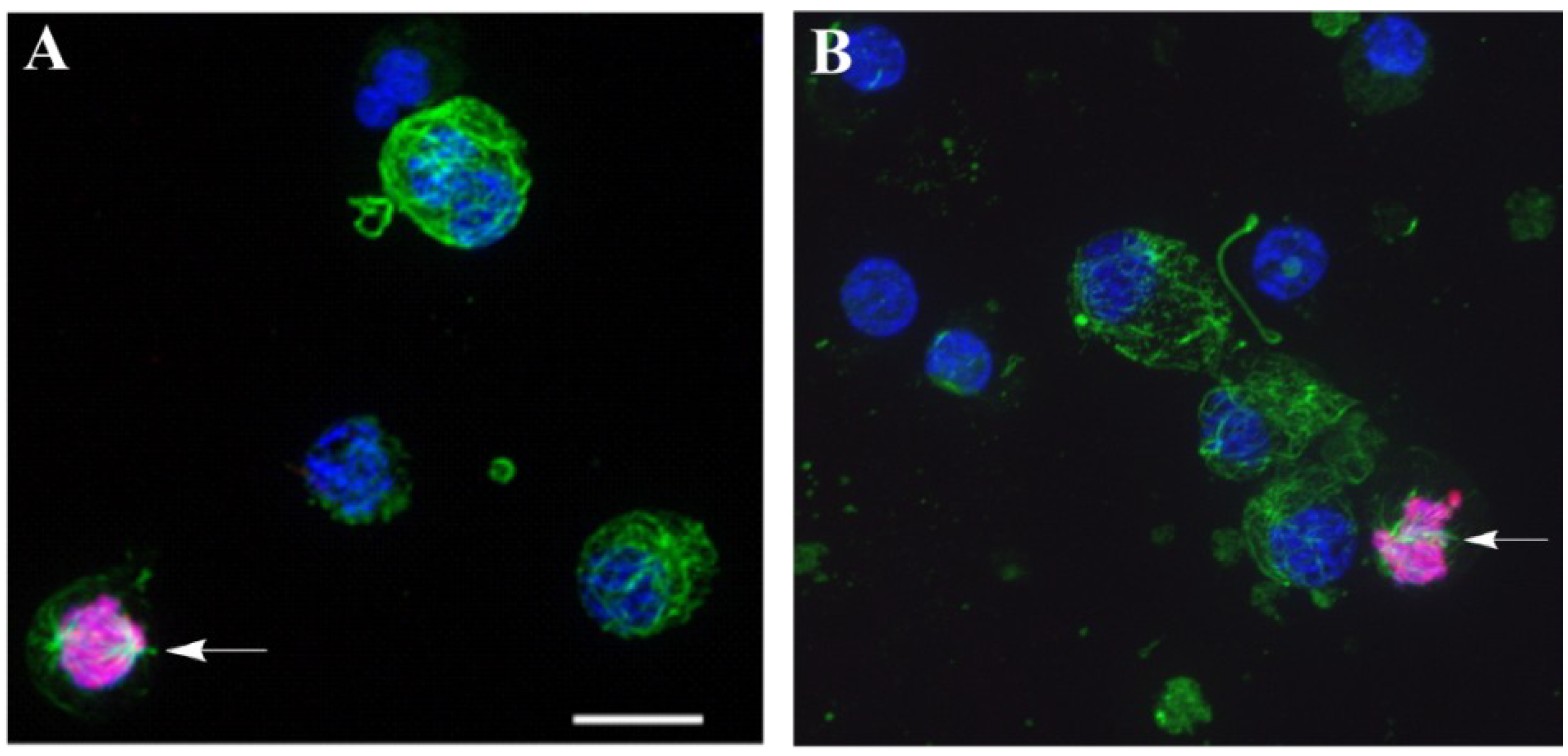
Figure 6.
Immunofluorescence detection of dividing (phospho-H3-histone-positive) cells in a blastula-derived culture. The preparations were imaged via confocal microscopy. The cells were cultivated on fibronectin-coated coverslips in coelomic fluid obtained from injured sea urchins (A) and in seawater (B) for 12 h and then labeled with Abs against phospho-H3-histone Abs for the detection of dividing cells (red) and tubulin (green) for the detection of microtubules. The nuclei were stained with DAPI (blue). Arrows show ph-H3-histone-positive cells. Scale bar, 10 µm.
Figure 6.
Immunofluorescence detection of dividing (phospho-H3-histone-positive) cells in a blastula-derived culture. The preparations were imaged via confocal microscopy. The cells were cultivated on fibronectin-coated coverslips in coelomic fluid obtained from injured sea urchins (A) and in seawater (B) for 12 h and then labeled with Abs against phospho-H3-histone Abs for the detection of dividing cells (red) and tubulin (green) for the detection of microtubules. The nuclei were stained with DAPI (blue). Arrows show ph-H3-histone-positive cells. Scale bar, 10 µm.
2.3. MALDI MS Analysis of Coelomic Fluids Obtained from Intact and Injured Sea Urchins
To estimate the contribution of specific components in intact and injured sea urchin coelomic fluids to the cell pigment differentiation process during cultivation and to understand its origin, we conducted MALDI MS analyses of both coelomic fluids of
S. intermedius.
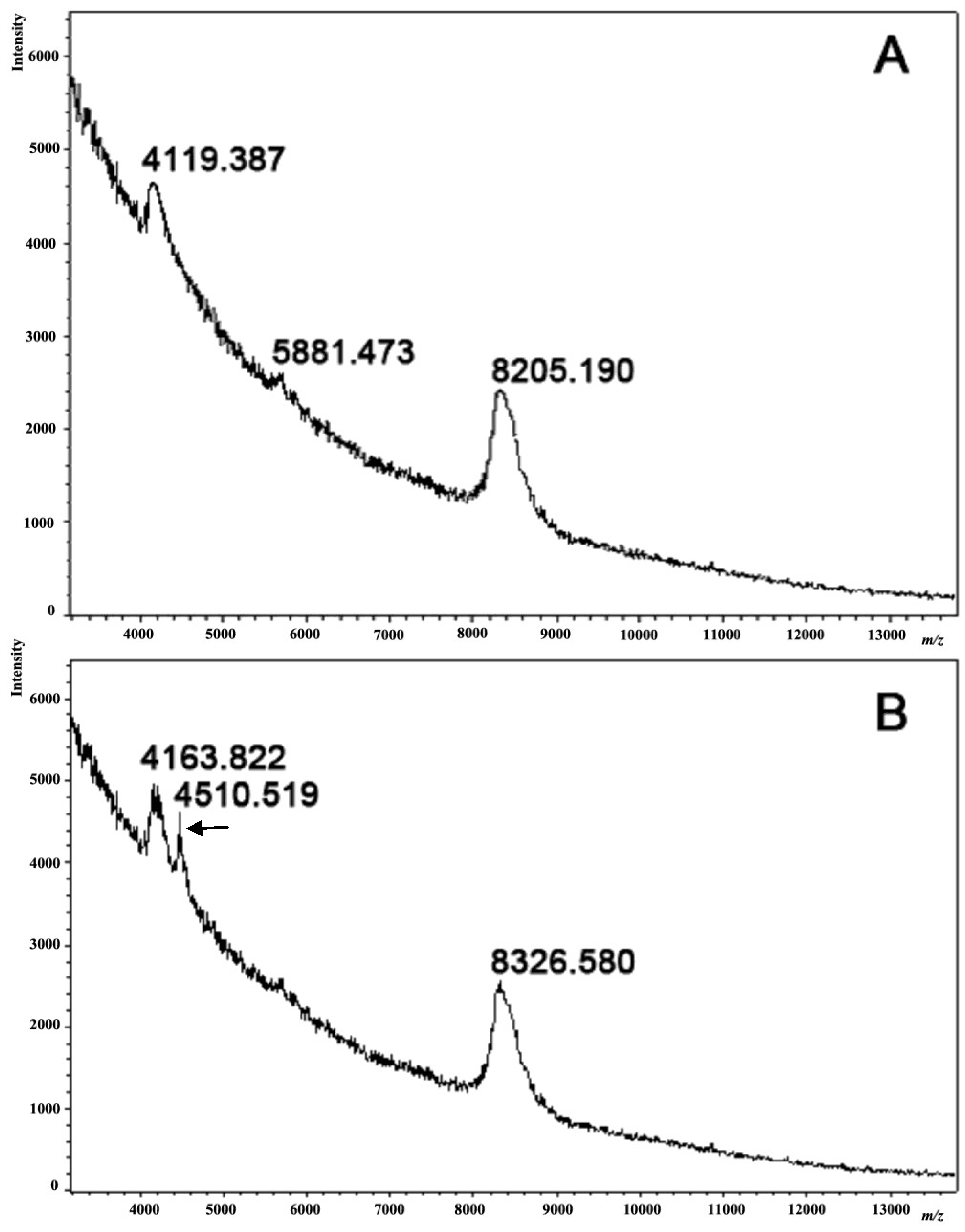
Figure 7 illustrates the MALDI mass spectra obtained for proteins with a molecular mass of 4000–9000 Da. Two well-defined peaks corresponding to proteins of 8205 Da and 4119 Da were detected in the spectrum of the intact sea urchin coelomic fluid (
Figure 7A). The spectrum of coelomic fluid obtained from the injured sea urchins exhibited three peaks: 8327 Da, 4511 Da, and 4164 Da (
Figure 7B). A comparison of the coelomic fluid profiles revealed a new peak in the coelomic fluid of the injured sea urchins corresponding to a protein with a molecular weight of around 4500 Da and a shift of approximately 40–120 Da from the basic components in the intact sea urchin coelomic fluid, which may be due to a protein modification such as phosphorylation, sulfation,
etc. The molecular weights of the proteins were measured using MALDI MS with standard precision (0.1%) based on the external calibration programs included (Protein Calibration Mixture II, Bruker Daltonik, Germany). The new peak in the spectrum (4511 Da) indicates a change in the composition of the proteins in the coelomic fluid after injury in comparison to the norm. Although we could not identify the molecular nature of the coelomic fluid proteins, our data clearly demonstrated a qualitative difference in the composition of normal coelomic-fluid proteins and that of coelomic-fluid proteins following injury.
Figure 7.
The mass spectra of coelomic fluids obtained from intact (A) and injured (B) sea urchins by MALDI TOF MS using a mass spectrometer (Ultraflex-III TOF/TOF, Bruker Daltonics, Germany). Note a new peak in the coelomic fluid of the injured sea urchins, corresponding to a protein with MW near 4500 Da (marked by an arrow), as well as the shift of the basic components of the intact coelomic fluid to approximately 40–120 Da. The coelomic fluids from three independent experiments were analyzed.
Figure 7.
The mass spectra of coelomic fluids obtained from intact (A) and injured (B) sea urchins by MALDI TOF MS using a mass spectrometer (Ultraflex-III TOF/TOF, Bruker Daltonics, Germany). Note a new peak in the coelomic fluid of the injured sea urchins, corresponding to a protein with MW near 4500 Da (marked by an arrow), as well as the shift of the basic components of the intact coelomic fluid to approximately 40–120 Da. The coelomic fluids from three independent experiments were analyzed.
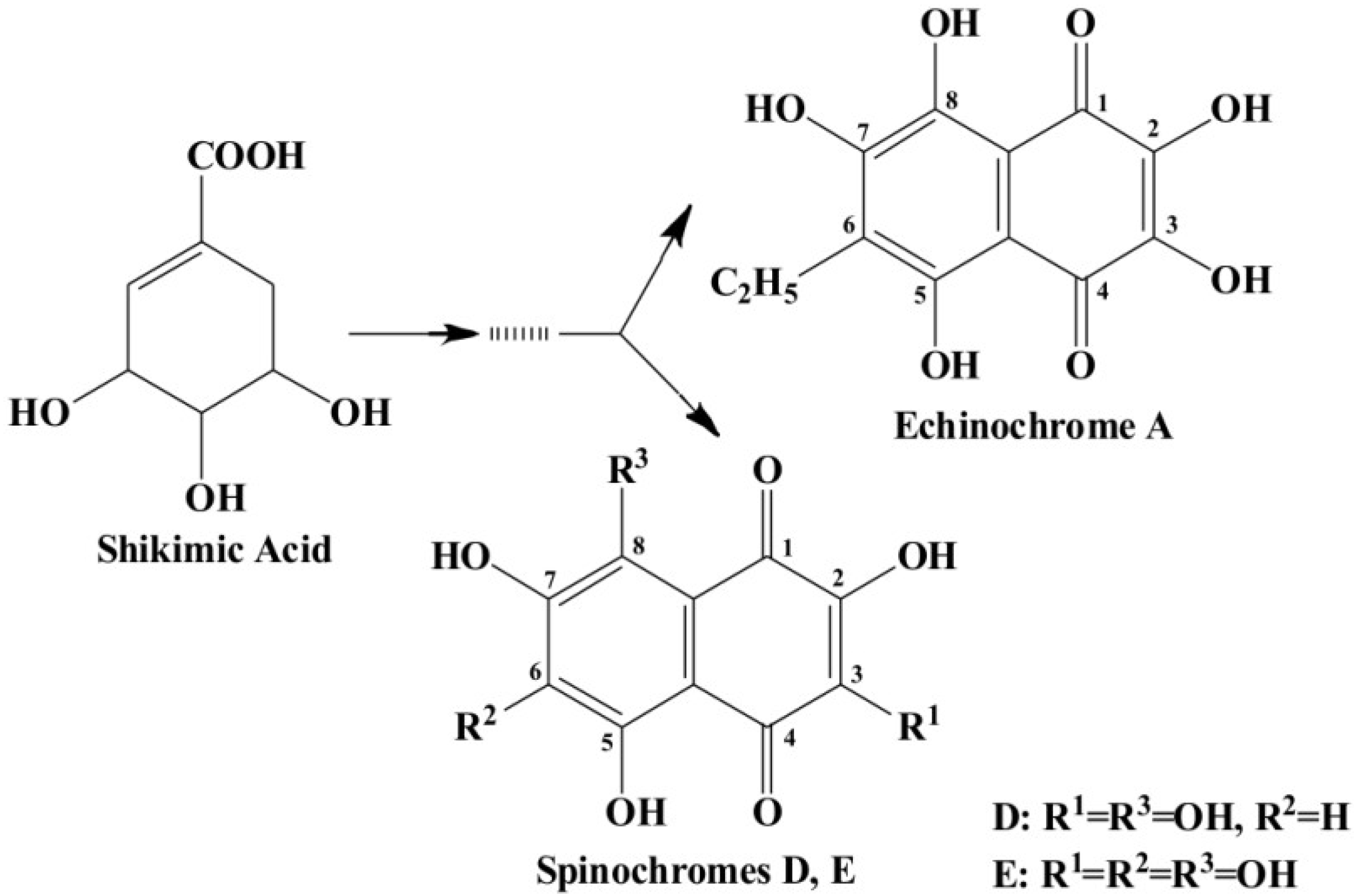
2.4. Naphthoquinone Pigment Production in Cultivated Sea Urchin Cells
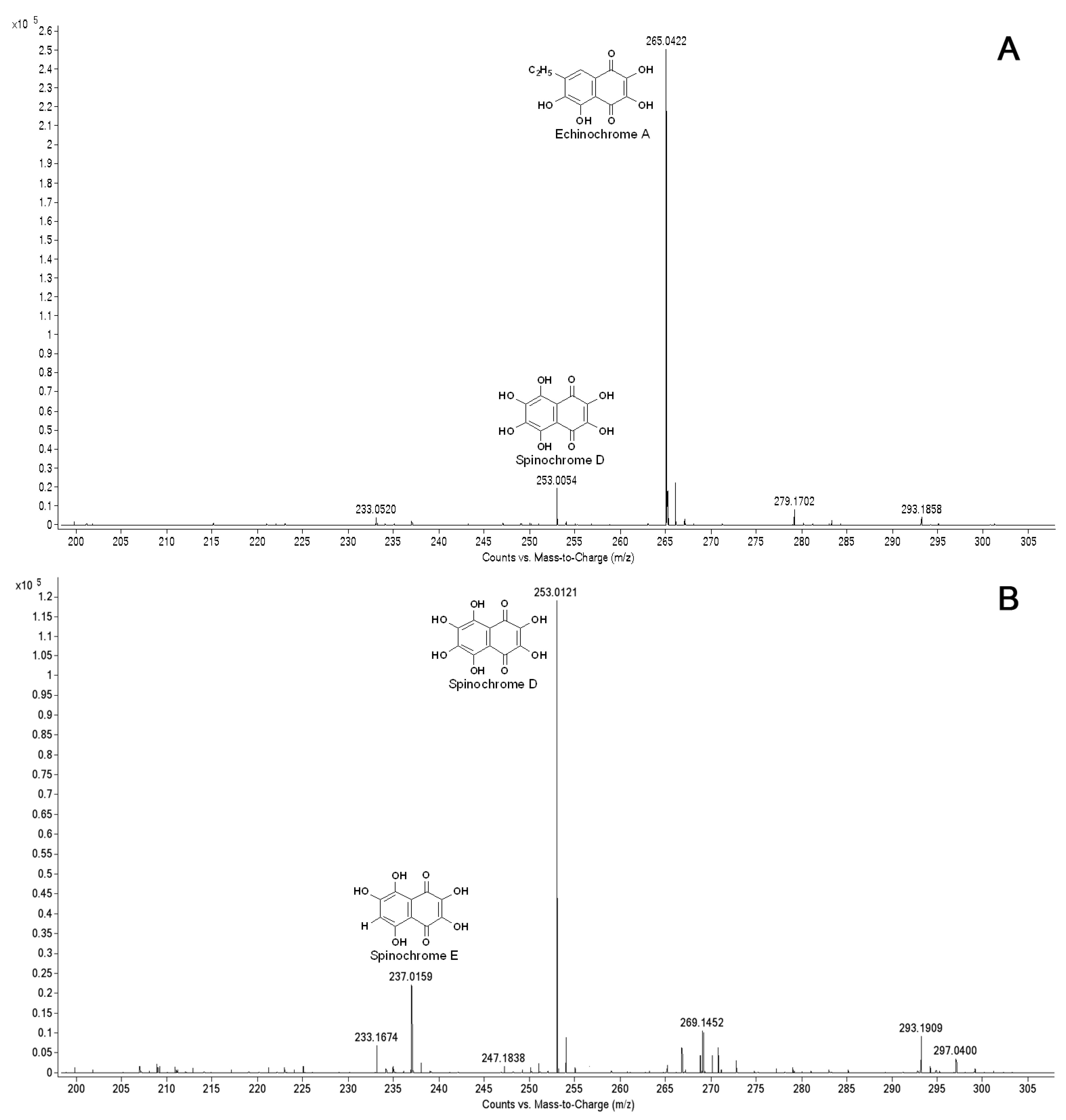
To elucidate the naphthoquinone pigment profile in the cultivated sea urchin cells and the production of pigments during cultivation, we performed ESI MS. As shown in
Figure 8A, a significant peak of echinochrome A and a small peak corresponding to spinochrome E were detected in the pigment extracts from the sand dollar
S. mirabilis, while only the peaks corresponding to spinochromes D and E were recorded in the pigment cell extracts from
S. intermedius (
Figure 8B). Thus, a distinct difference in pigment expression was demonstrated in the cells of the two sea urchin species.
Figure 8.
ESI MS profiles of naphthoquinone pigments from sea urchin cell cultures: A—the pigment extracts from the cells of the sea urchin S. mirabilis after 3-day cultivation in seawater; B—the pigment extracts from the cells of the sea urchin S. intermedius after 6-day cultivation in seawater.
Figure 8.
ESI MS profiles of naphthoquinone pigments from sea urchin cell cultures: A—the pigment extracts from the cells of the sea urchin S. mirabilis after 3-day cultivation in seawater; B—the pigment extracts from the cells of the sea urchin S. intermedius after 6-day cultivation in seawater.
The results of a semi-quantitative measurement (using ESI MS) of the echinochrome and spinochrome production in cells cultivated in the different media are presented in
Table 1. Echinochrome A was produced by cultivated
S. mirabilis cells in all of the tested media, and it was well defined in all of the spectra. The highest level of echinochrome A was detected in the cells cultured in the coelomic fluids as compared to those cultured in seawater. In contrast, no significant differences were seen in the spinochrome D and E levels in the tested media for the cultivated cells of the other sea urchin,
S. intermedius (
Table 1).
Table 1.
Naphthoquinone pigment production in sea urchin cultivated cells. Data are presented as the mean ± standard error from two independent experiments (ESI MS). SW—seawater; CFn—coelomic fluid obtained from intact sea urchins; CFreg—coelomic fluid obtained from injured sea urchins.
2.5. Expression of the pks Genes in Cultivated Sea Urchin Cells. Experiments with Shikimic Acid, a Precursor of Naphthoquinone Pigments
To understand the molecular mechanisms underlying sea urchin pigment specialization in culture, we evaluated the gene expression associated with the induction of pigment differentiation, particularly the
pks genes. The expression level of these genes (estimated by quantitative real-time PCR) in sea urchin cells cultivated in different culture media is presented in relative units in
Figure 9 (see the
Experimental Section). The gene expression in sea urchin cells cultivated in the coelomic fluid of injured sea urchins increased more than 1.2-fold in comparison to the cells cultivated in seawater, whereas the
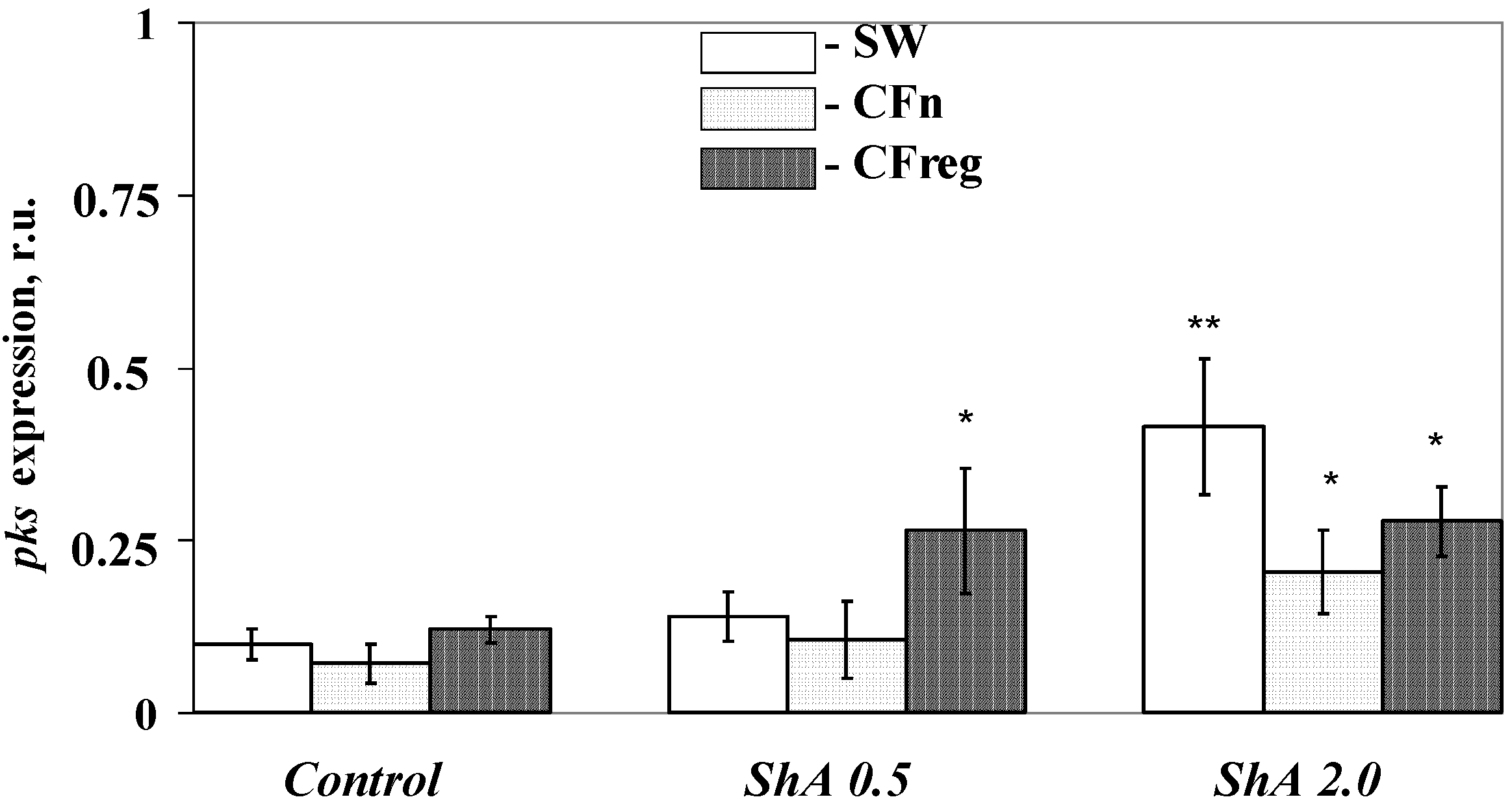
pks expression in the cells cultivated in the coelomic fluid from intact sea urchins decreased by almost 1.4-fold; however, these differences were insignificant. The addition of a high concentration of ShА, a precursor of naphthoquinone pigments, (2 mM) to the sea urchin cells resulted in a marked intensification of the gene expression in all media tested (2.3–4.2-fold) compared to the control cells (
Figure 9), but the effect was especially significant in seawater. In contrast, adding ShА of a lower concentration (0.5 mM) increased
pks expression only in the cells cultivated in injured sea urchin coelomic fluid (2.2-fold). No apparent effect on
pks expression was detected in the cells cultivated with 0.5 mM ShA in seawater or intact sea urchin coelomic fluid. These data can be explained by the presence of some specific components in the sea urchin coelomic fluids. The marked intensification of the
pks expression (2.2-fold) in the presence of 0.5 mM ShА only in the cells cultivated in injured sea urchin coelomic fluid is additional evidence of a cellular reaction in response to injury signals. Our findings showing a significant difference between the number of sea urchin pigment cells cultivated in the coelomic fluids of injured and intact sea urchins support this suggestion.
Figure 9.
Effect of Shikimic Acid (ShA) on the pks gene expression in a blastula-derived cell culture of the sea urchin Strongylocentrotus intermedius. ShA concentrations tested: 0.5 mM and 2.0 mM. Y-axis—relative units (r.u.). The cells were cultivated with ShA for 4 days in: seawater (SW), coelomic fluid of intact sea urchins (CFn), and coelomic fluid of injured sea urchins (CFreg). After this period, total RNA was isolated from cultivated cells for the following real-time PCR. Сontrol cells are the cells cultivated without ShA. * p < 0.05; ** p < 0.01 versus values of pks expression in the appropriate control conditions.
Figure 9.
Effect of Shikimic Acid (ShA) on the pks gene expression in a blastula-derived cell culture of the sea urchin Strongylocentrotus intermedius. ShA concentrations tested: 0.5 mM and 2.0 mM. Y-axis—relative units (r.u.). The cells were cultivated with ShA for 4 days in: seawater (SW), coelomic fluid of intact sea urchins (CFn), and coelomic fluid of injured sea urchins (CFreg). After this period, total RNA was isolated from cultivated cells for the following real-time PCR. Сontrol cells are the cells cultivated without ShA. * p < 0.05; ** p < 0.01 versus values of pks expression in the appropriate control conditions.
2.6. Discussion
Marine biotechnology progress is associated with the search, characterization, and development of optimal methods for obtaining new substances from marine organisms with pharmaceutically interesting biological activity, and many are of enormous scientific interest. On the one hand, marine organisms are a source of unique marine secondary metabolites that are the basis for developing new and improved natural products for commercial purposes [
2]. On the other hand, they include a variety of phylogenetic groups that are important for understanding the evolutionary history of life. Some, such as the echinoderms, are members of a phylum of deuterostomate metazoans and occupy a key position in the early steps in chordate evolution [
20,
21]. As shown by Evans-Illidge
et al. [
1], the ancestral early-metazoans (Porifera), early eumetazoans (Cnidaria) and some deuterostomes (Echinodermata) have a very high concentration of bioactive compounds.
Here, we have shown that the culture medium composition affects the rate of appearance of pigment cells. In seawater, the number of dividing cells was higher than when the cells were cultured in the coelomic fluids, but the pigment cell differentiation occurred slowly and less extensively. An increase in cell pigmentation in the coelomic fluids probably leads to a decrease in the number of dividing cells in comparison to the cells incubated in seawater. In the early stages of cultivation, we detected dividing cells and a few pigment cells, but later during the pigment cell differentiation, we observed a decrease in the number of dividing cells and a corresponding increase in the number of pigment cells. This is not surprising: either proliferation or differentiation usually dominates in cell cultures. A significant decrease in the number of dividing cells can be explained by a general reduction of mitotic activity during cultivation that has been previously reported for many invertebrate cells after 7–10 days of cultivation [
22,
23].
We proposed that a significant difference in the proportion of pigment cells in the cultures grown in the coelomic fluids of intact and injured sea urchins in comparison to those cultivated in seawater was connected to the specific components of coelomic fluids that increase the adhesion of cultivated cells. Recently, approximately 30 proteins that may be involved in adhesion were uncovered by a large-scale proteomic analysis of sea urchin coelomic fluid [
24]. Thus, far, a proteomic analysis has only been performed on normal sea urchin coelomic fluid.
In this study, we analyzed the coelomic fluid of both normal and injured sea urchins, and detected qualitative differences in their composition. Our results from the overview of integral MALDI mass spectra of sea urchin coelomic fluids have shown that new peaks appear in the coelomic fluid obtained from injured sea urchins. These new peaks may correspond to stress proteins produced after injury. As recently reported, relatively abundant proteins involved in stress responses were identified in the coelomic fluid of the sea urchin
S. purpuratus [
24]. Homologues of these proteins have been found throughout the animal kingdom and are often used as markers for environmental stress [
25]. We have suggested that the new coelomic fluid proteins from the injured sea urchins may be related to stress-response proteins. The MALDI MS data together with the results for
pks gene expression suggest that the specific components of sea urchin coelomic fluids play a role in pigment differentiation through the regulation of the genes implicated in naphthoquinone synthesis. Analyzing cells cultivated in the presence of 0.5 mM ShА, we showed that
pks gene expression was two-fold higher in cells cultivated in the injured sea urchin coelomic fluid relative to cells cultivated in that from intact sea urchins. A higher proportion of cells containing naphthoquinones in response to different conditions of stress could be a consequence of a protective reaction and be important as a defense mechanism [
26,
27].
We found that echinochrome was produced only in the sand dollar cells and its content in cells grown in coelomic fluid was higher than that in cells grown in seawater. This finding suggests the existence of a specific regulatory factor that may induce pigmentation activity and subsequent naphthoquinone production in these coelomic fluids. Our morphological observations of the cultured cells correlated with the semi-quantitative data obtained using ESI MS. Unfortunately, not all of the experiments could be performed on the S. mirabilis cell cultures due to the difficulty in obtaining the sand dollar cells free of bacterial contamination.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




