Obtaining Spheroplasts of Armored Dinoflagellates and First Single-Channel Recordings of Their Ion Channels Using Patch-Clamping

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction

2. Results and Discussion
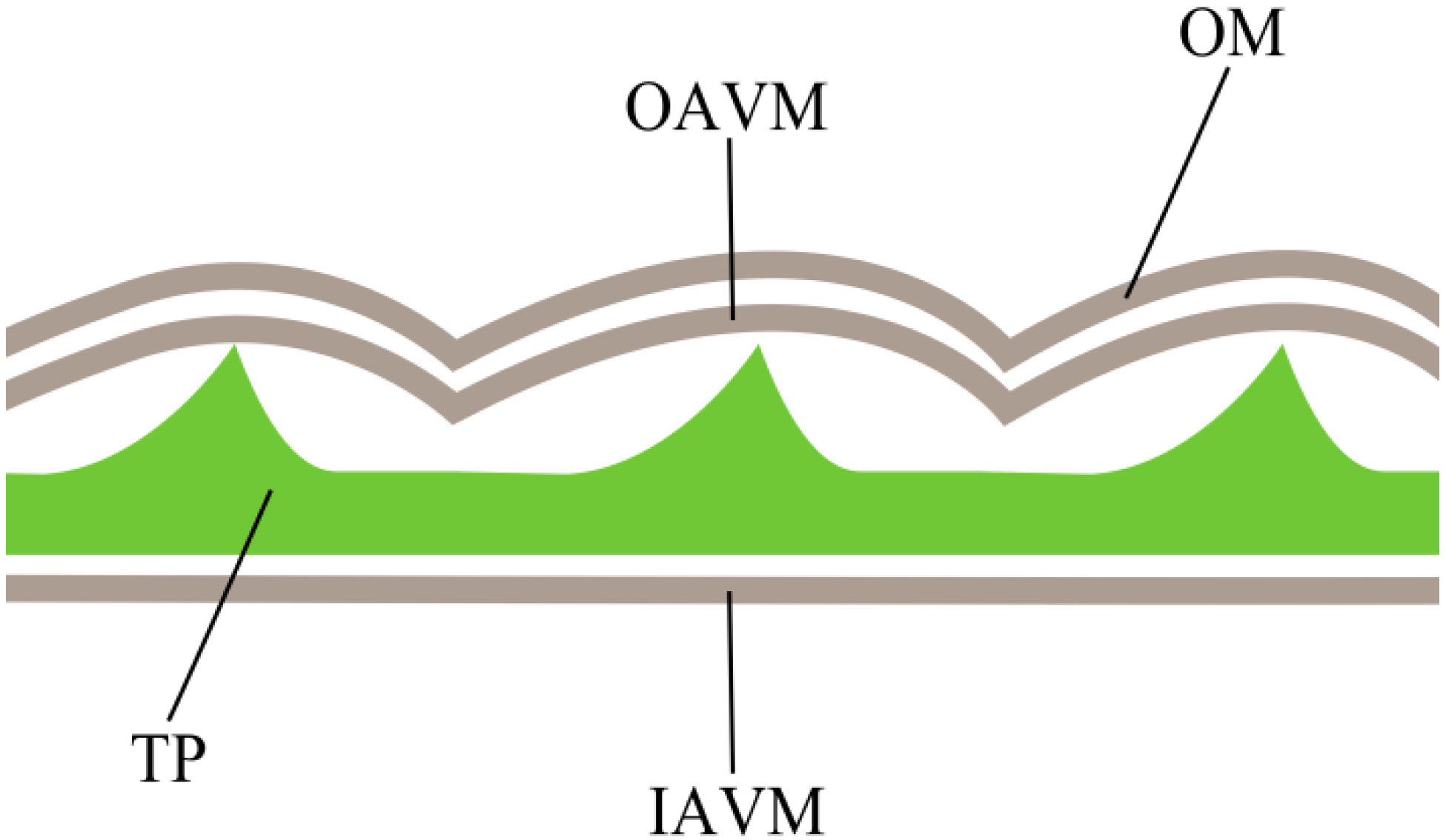
2.1. Ecdysed Cells
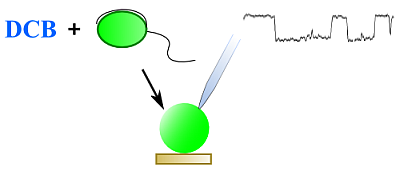
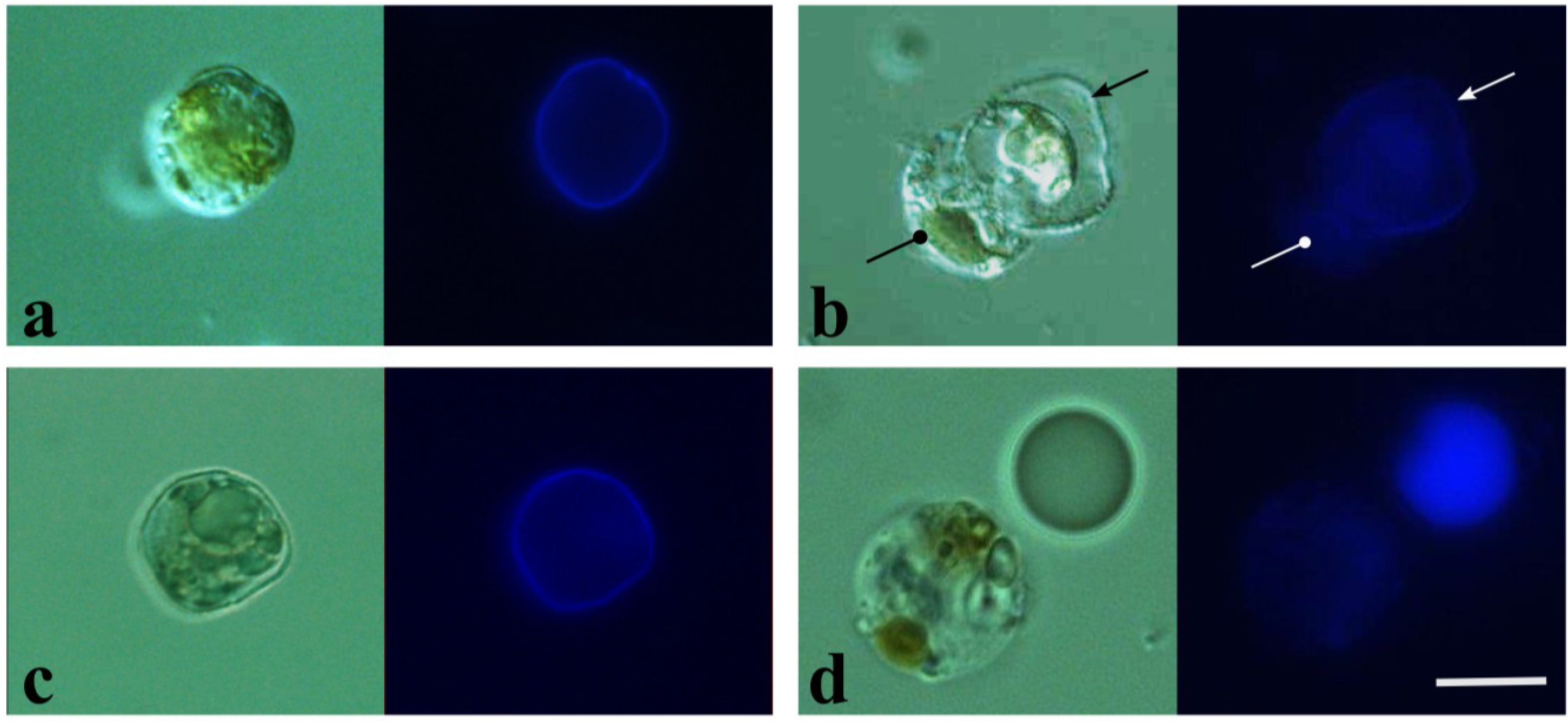
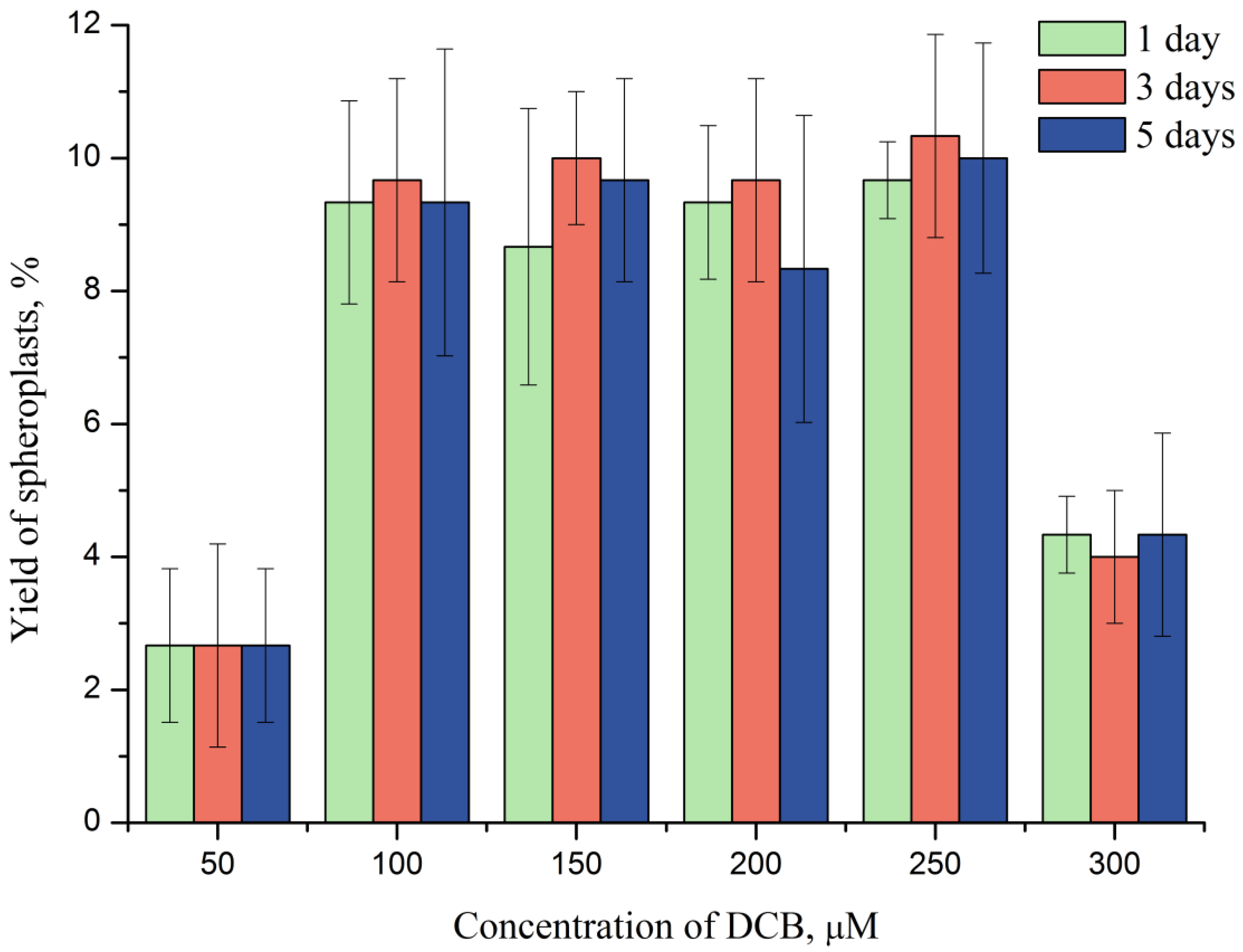
2.2. DCB-Induced Spheroplasts


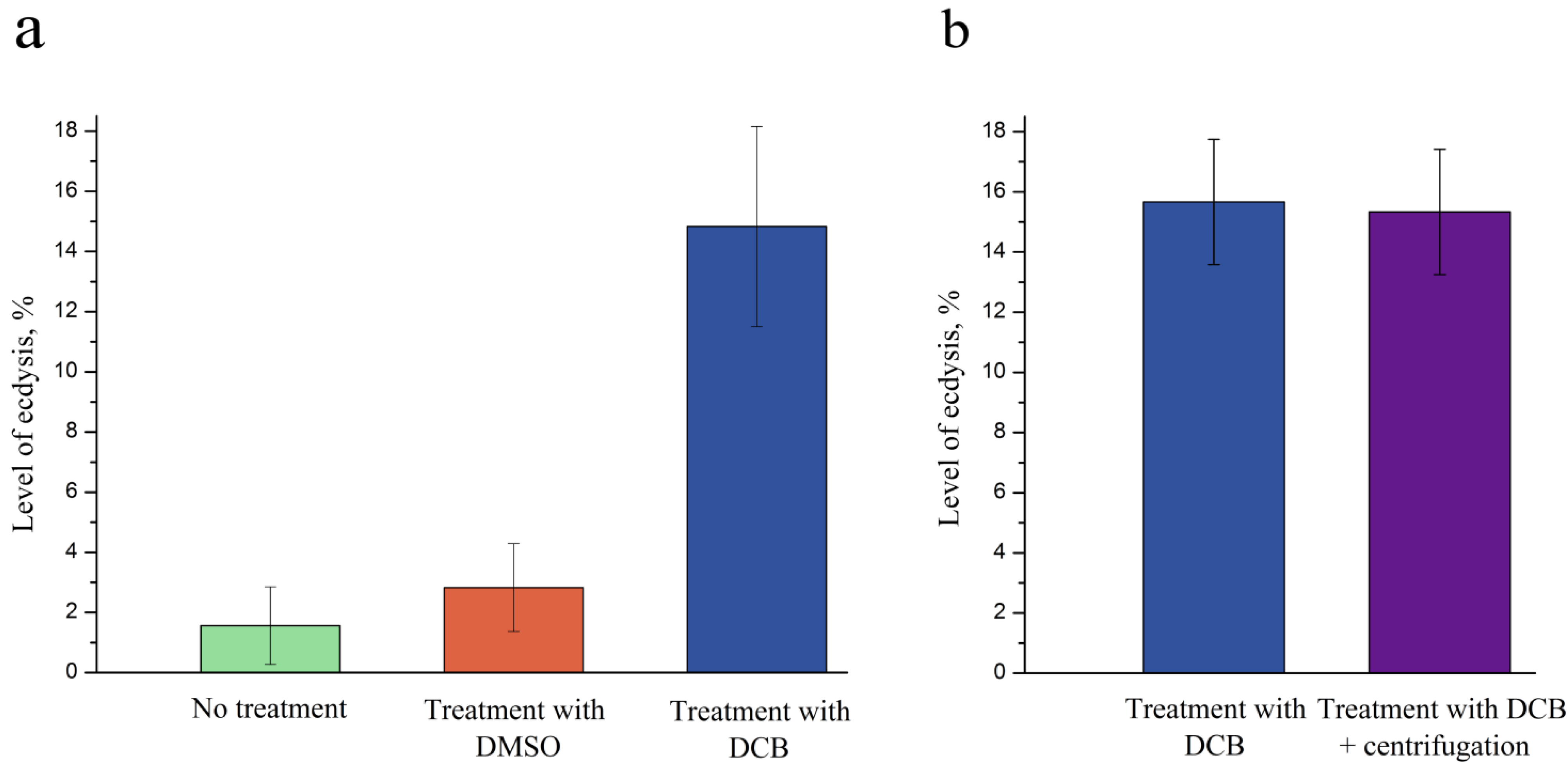
2.3. DCB as an Inducer of Ecdysis in P. minimum

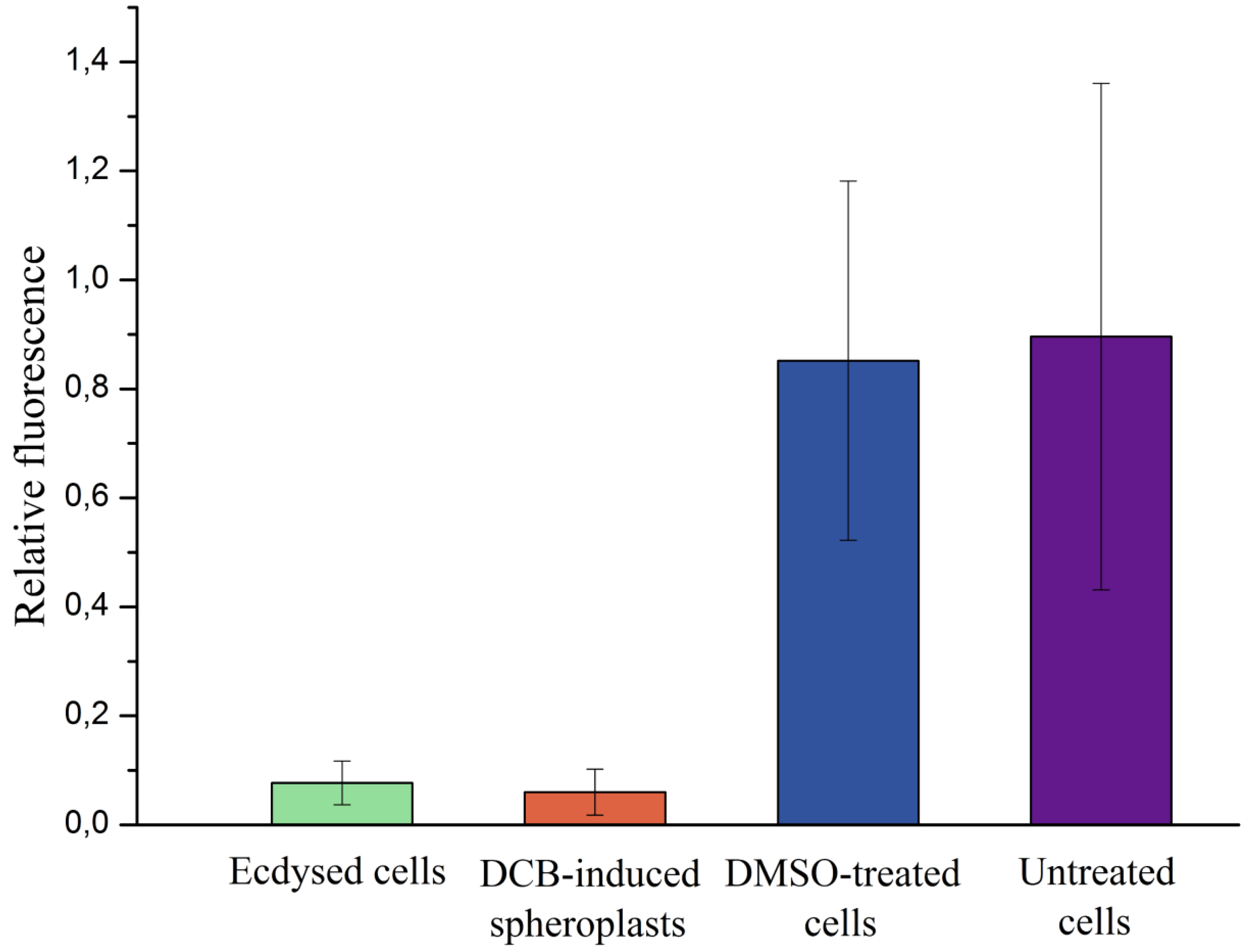
2.4. Ecdysed Cells vs. DCB-Induced Spheroplasts

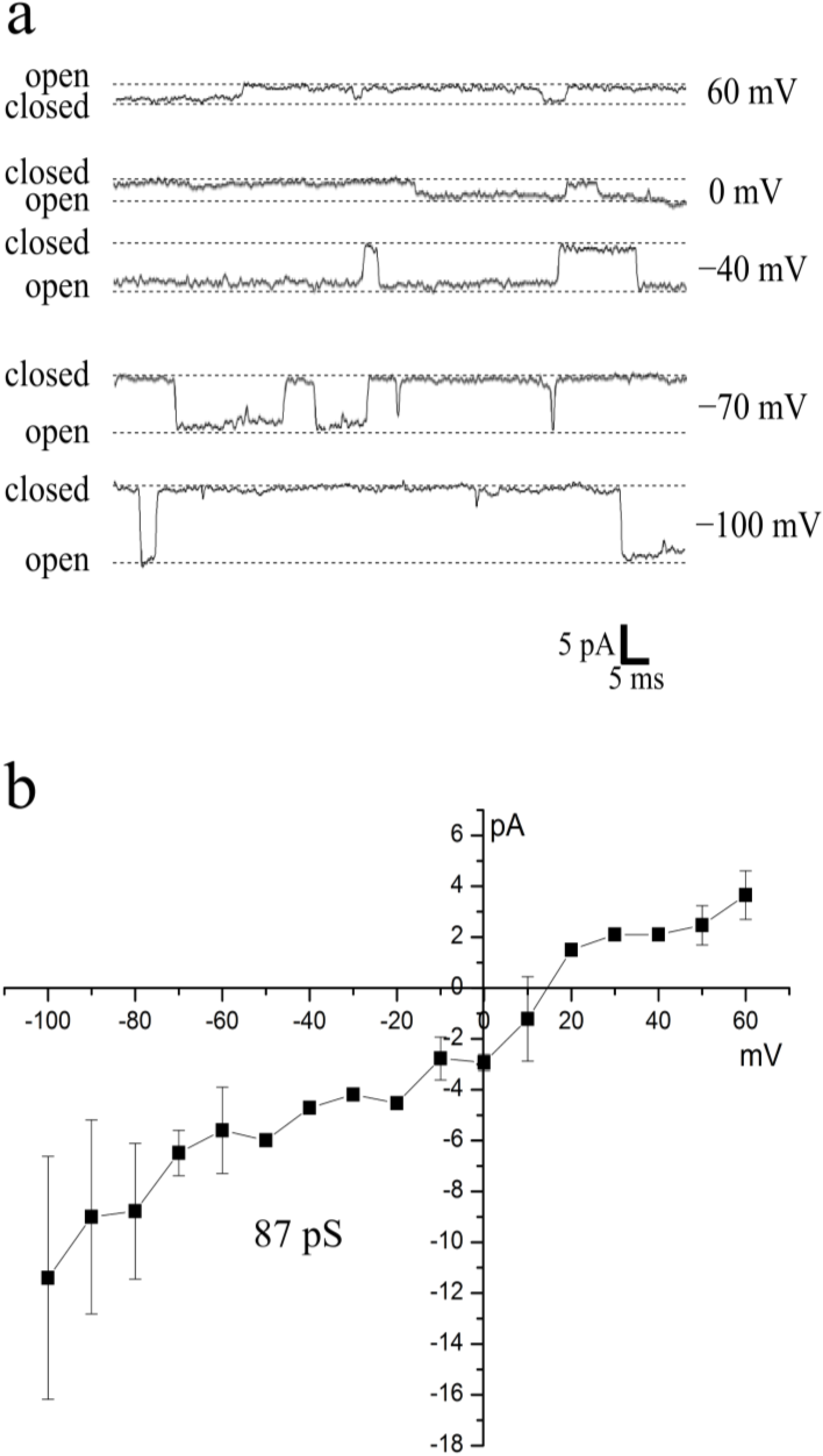
2.5. First Single-Channel Recordings in Armored Dinoflagellates

3. Experimental Section
3.1. Culture Conditions
3.2. Preparation of Spheroplasts
3.2.1. Physical Treatment
3.2.2. Chemical Treatment
3.3. Patch-Clamp Procedures
3.4. Cell Counting and Calculation of the Spheroplasts Yield and Ecdysis Level
3.5. Cellulose Staining and Microscopy
3.6. Image Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Martinac, B.; Saimi, Y.; Kung, C. Ion channels in microbes. Physiol. Rev. 2008, 88, 1449–1490. [Google Scholar] [CrossRef]
- Beck, C.; Uhl, R. On the localization of voltage-sensitive calcium channels in the flagella of Chlamydomonas reinhardtii. J. Cell Biol. 1994, 125, 1119–1125. [Google Scholar] [CrossRef]
- Saimi, Y.; Hinrichsen, R.D.; Forte, M.; Kung, C. Mutant analysis shows that Ca2+-induced K+ current shuts off one type of excitation in Paramecium. Proc. Natl. Acad. Sci. USA 1983, 80, 5112–5116. [Google Scholar] [CrossRef]
- Kink, J.A.; Maley, M.E.; Preston, R.R.; Ling, K.Y.; Wallen-Friedman, M.A.; Saimi, Y; Kung, C. Mutation in paramecium calmodulin indicate functional differences between the C-terminal and N-terminal lobes in vivo. Cell 1990, 62, 165–174. [Google Scholar]
- Eckert, R.; Sibaoka, T. The flash-triggering action potential of luminescent dinoflagellate Noctiluca. J. Gen. Physiol. 1968, 52, 258–282. [Google Scholar] [CrossRef]
- Smith, S.; Morgan, D.; Musset, B.; Cherny, V.; Place, A.; Woodland Hastings, J.; DeCoursey, T. Voltage-gated proton channel in a dinoflagellate. Proc. Natl. Acad. Sci. USA 2011, 108, 18162–18167. [Google Scholar] [CrossRef]
- Taylor, A.R.; Chrachri, A.; Wheeler, G.; Goddard, H.; Brownlee, C. A voltage-gated H+ channel underlying pH homeostasis in calcifying coccolithophores. PLoS Biol. 2011, 9, e1001085:1–e1001085:14. [Google Scholar] [CrossRef]
- Hackett, J.; Anderson, D.; Erdner, D.; Bhattacharya, D. Dinoflagellates: A remarkable evolutionary experiment. Am. J. Bot. 2004, 9, 1523–1534. [Google Scholar] [CrossRef]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stühmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Phil. Trans. R. Soc. B 2014, 369, 20130094:1–20130094:9. [Google Scholar] [CrossRef]
- Bortner, C.D.; Cidlowsky, J.A. Ion channels and apoptosis in cancer. Phil. Trans. R. Soc. B 2014, 369, 20130104:1–20130104:9. [Google Scholar] [CrossRef]
- Molleman, A. Patch Clamping: An Introductory Guide to Patch Clamp Electrophysiology; John Willey and Sons, Ltd.: Chichester, West Sussex, UK, 2003. [Google Scholar]
- Elzenga, J.T.M.; Keller, C.P.; van Volkenburgh, E. Patch clamping protoplasts from vascular plants. Plant Physiol. 1991, 97, 1573–1575. [Google Scholar] [CrossRef]
- Garrill, A.; Davies, J.M. Patch clamping fungal membranes: A new perspective on ion transport. Mycol. Res. 1994, 98, 257–263. [Google Scholar] [CrossRef]
- Morrill, L.C.; Loeblich, A.R., III. Ultrastructure of the dinoflagellate amphiesma. Int. Rev. Cytol. 1983, 82, 151–180. [Google Scholar]
- Pozdnyakov, I.; Skarlato, S. Dinoflagellate amphiesma at different stages of the life cycle. Protistology 2012, 7, 108–115. [Google Scholar]
- Dodge, J.D. Thecal fine-structure in the dinoflagellate genera Prorocentrum and Excuvialle. J. Mar. Biol. Ass. UK 1965, 45, 607–614. [Google Scholar] [CrossRef]
- Adamich, M.; Sweeney, B.M. The preparation and characterization of Gonyaulax spheroplasts. Planta 1976, 130, 1–6. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Binder, B.J. Sexuality and cyst formation in the dinoflagellate Gonyaulax tamarensis: Cyst yield in batch cultures. J. Phycol. 1984, 20, 418–425. [Google Scholar] [CrossRef]
- Hardeland, R. Induction of cyst formation by low temperature in the dinoflagellate Gonyaulax polyedra Stein: Dependence on circadian phase and requirement of light. Experientia 1994, 50, 60–62. [Google Scholar] [CrossRef]
- Wong, J.T.Y.; Wong, Y.H. Indoleamine-induced encystment in dinoflagellates. J. Mar. Biol. Ass. UK 1994, 74, 467–469. [Google Scholar] [CrossRef]
- Bravo, I.; Figueroa, R.I. Towards an ecological understanding of dinoflagellate cyst functions. Microorganisms 2014, 2, 11–32. [Google Scholar]
- Kwok, A.C.M.; Mak, C.C.M.; Wong, F.T.W.; Wong, J.T.Y. Novel method for preparing spheroplasts from cells with an internal cellulosic cell wall. Eukaryot. Cell 2007, 6, 563–567. [Google Scholar] [CrossRef]
- Mayer, Y.; Herth, W. Chemical inhibition of cell wall formation and cytokinesis, but not of nuclear division, in protoplasts of Nicotinia tabacum L. cultured in vitro. Planta 1978, 142, 253–262. [Google Scholar]
- Morrill, L.C. Ecdysis and the location of the plasma membrane in the dinoflagellate Heterocapsa niei. Protoplasma 1984, 119, 8–20. [Google Scholar] [CrossRef]
- Kwok, A.C.M.; Wong, J.T.Y. Cellulose synthesis is coupled to cell cycle progression at G1 in the dinoflagellate Crypthecodinium cohnii. Plant Physiol. 2003, 131, 1681–1691. [Google Scholar] [CrossRef]
- Sheldletzky, E.; Shmuel, M.; Trainin, T.; Kalman, S.; Delmer, D. Cell wall structure in cells adapted to growth on the cellulose-synthesis inhibitor 2,6-dichlorobenzonitrile. Plant Physiol. 1992, 100, 120–130. [Google Scholar] [CrossRef]
- Mizuta, S.; Brown, R.M. Effects of 2,6-dichlorobenzonitrile and tinopal LPW on the structure of the cellulose synthesizing complexes of Vaucheria hamata. Protoplasma 1992, 166, 200–207. [Google Scholar] [CrossRef]
- Dickson, D.M.; Kirst, G.O. Osmotic adjustment in marine eukaryotic algae: The role of inorganic ions, quaternary ammonium, tertiary sulphonium and carbohydrate solutes. I. Diatoms and rhodophyte. New Phytol. 1987, 106, 645–655. [Google Scholar] [CrossRef]
- Dickson, D.M.; Kirst, G.O. Osmotic adjustment in marine eukaryotic algae: The role of inorganic ions, quaternary ammonium, tertiary sulphonium and carbohydrate solutes. II. Prasinophytes and haptophytes. New Phytol. 1987, 106, 657–666. [Google Scholar] [CrossRef]
- Oami, K.; Sibaoka, T.; Naitoh, Y. Tentacle regulating potentials in Noctiluca miliaris: Their generation sites and ionic mechanisms. J. Comp. Physiol. A 1988, 162, 179–185. [Google Scholar] [CrossRef]
- Oami, K.; Naitoh, Y. Distribution of ion channels in the membrane of the dinoflagellate Noctiluca miliaris. J. Exp. Biol. 1990, 150, 473–478. [Google Scholar]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine diatoms I. Cyclistella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar]
- Rasband, W.S. ImageJ, 1.47v; U.S. National Institutes of Health: Bethesda, MD, USA, 2013. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pozdnyakov, I.; Matantseva, O.; Negulyaev, Y.; Skarlato, S. Obtaining Spheroplasts of Armored Dinoflagellates and First Single-Channel Recordings of Their Ion Channels Using Patch-Clamping. Mar. Drugs 2014, 12, 4743-4755. https://doi.org/10.3390/md12094743
Pozdnyakov I, Matantseva O, Negulyaev Y, Skarlato S. Obtaining Spheroplasts of Armored Dinoflagellates and First Single-Channel Recordings of Their Ion Channels Using Patch-Clamping. Marine Drugs. 2014; 12(9):4743-4755. https://doi.org/10.3390/md12094743
Chicago/Turabian StylePozdnyakov, Ilya, Olga Matantseva, Yuri Negulyaev, and Sergei Skarlato. 2014. "Obtaining Spheroplasts of Armored Dinoflagellates and First Single-Channel Recordings of Their Ion Channels Using Patch-Clamping" Marine Drugs 12, no. 9: 4743-4755. https://doi.org/10.3390/md12094743
APA StylePozdnyakov, I., Matantseva, O., Negulyaev, Y., & Skarlato, S. (2014). Obtaining Spheroplasts of Armored Dinoflagellates and First Single-Channel Recordings of Their Ion Channels Using Patch-Clamping. Marine Drugs, 12(9), 4743-4755. https://doi.org/10.3390/md12094743