
Acetylated Chitosan Oligosaccharides Act as Antagonists against Glutamate-Induced PC12 Cell Death via Bcl-2/Bax Signal Pathway



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
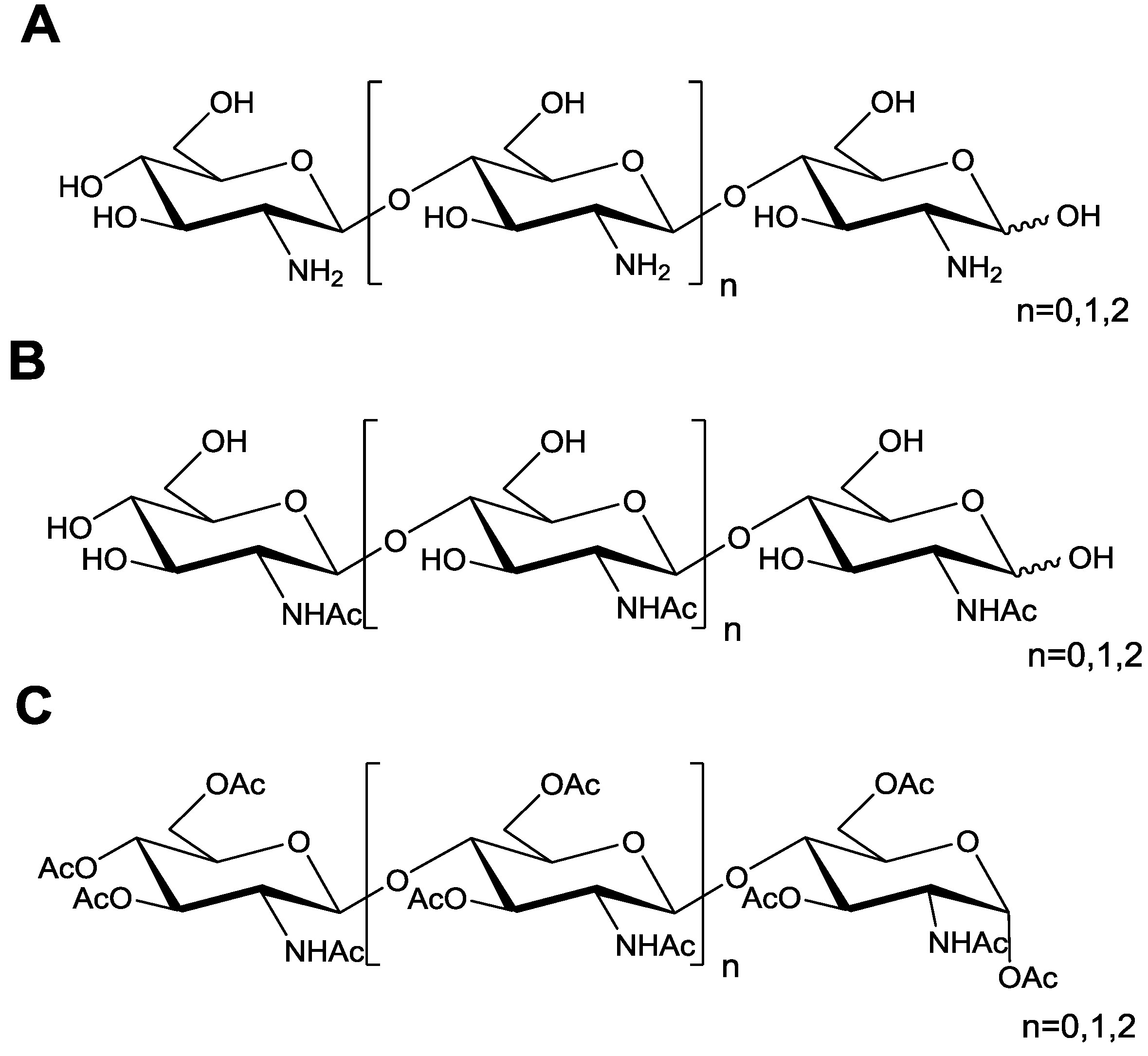
2.1. Characterization of Chitosan Oligosaccharides and Its Acetylated Derivatives

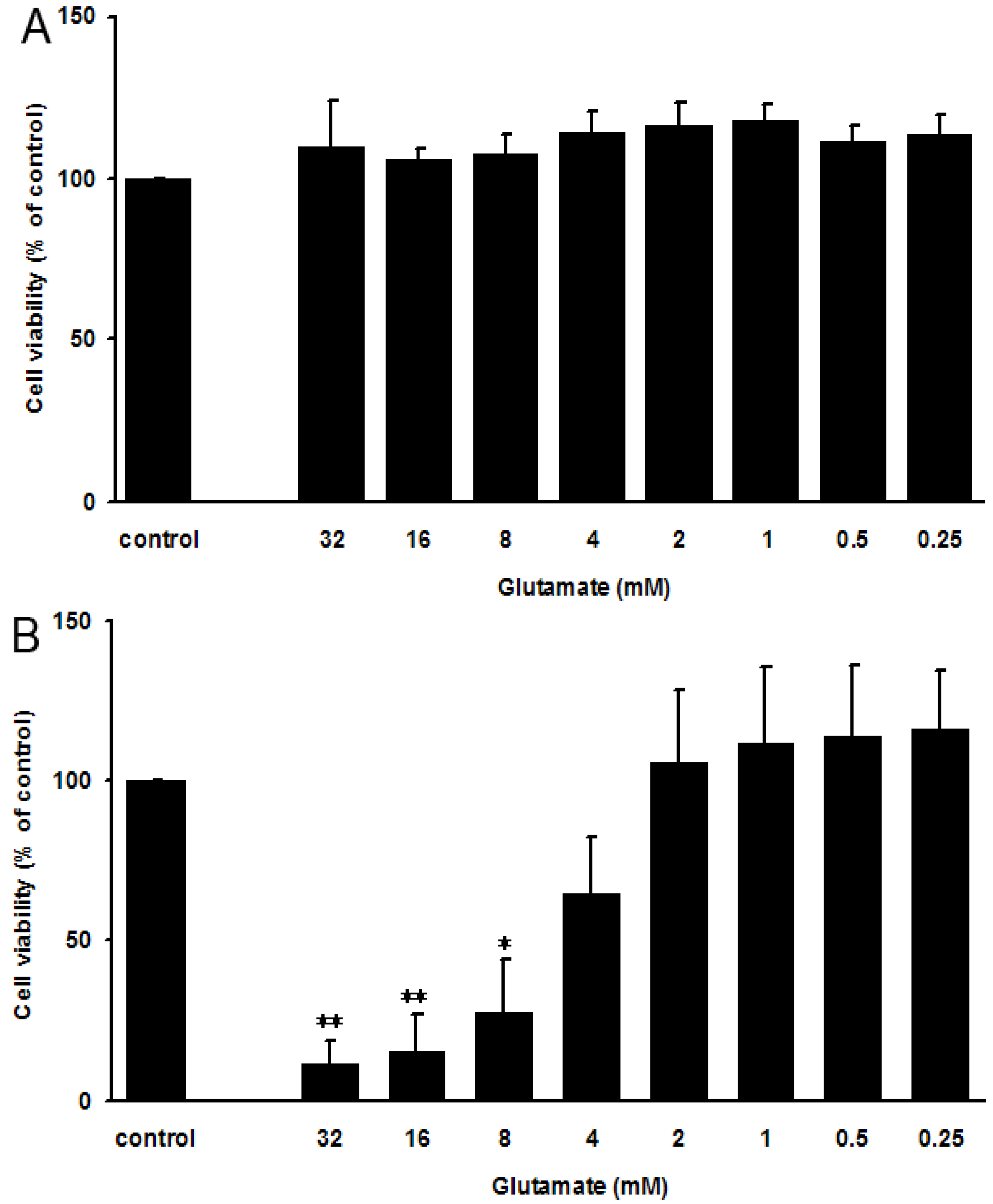
2.2. Glutamate-Induced PC12 Cell Death

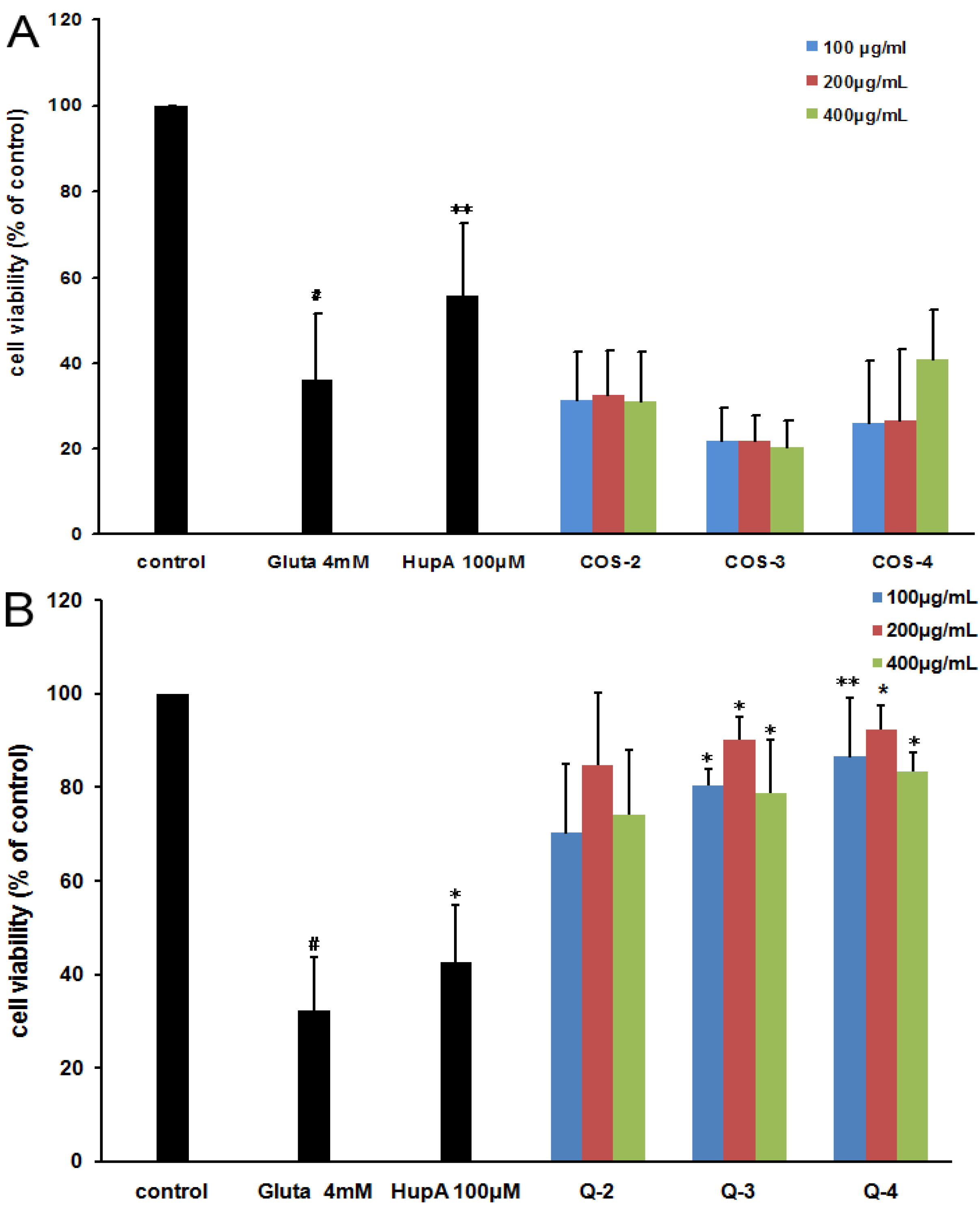
2.3. Oligosaccharides as Antagonists against Glutamate-Induced PC12 Cell Death


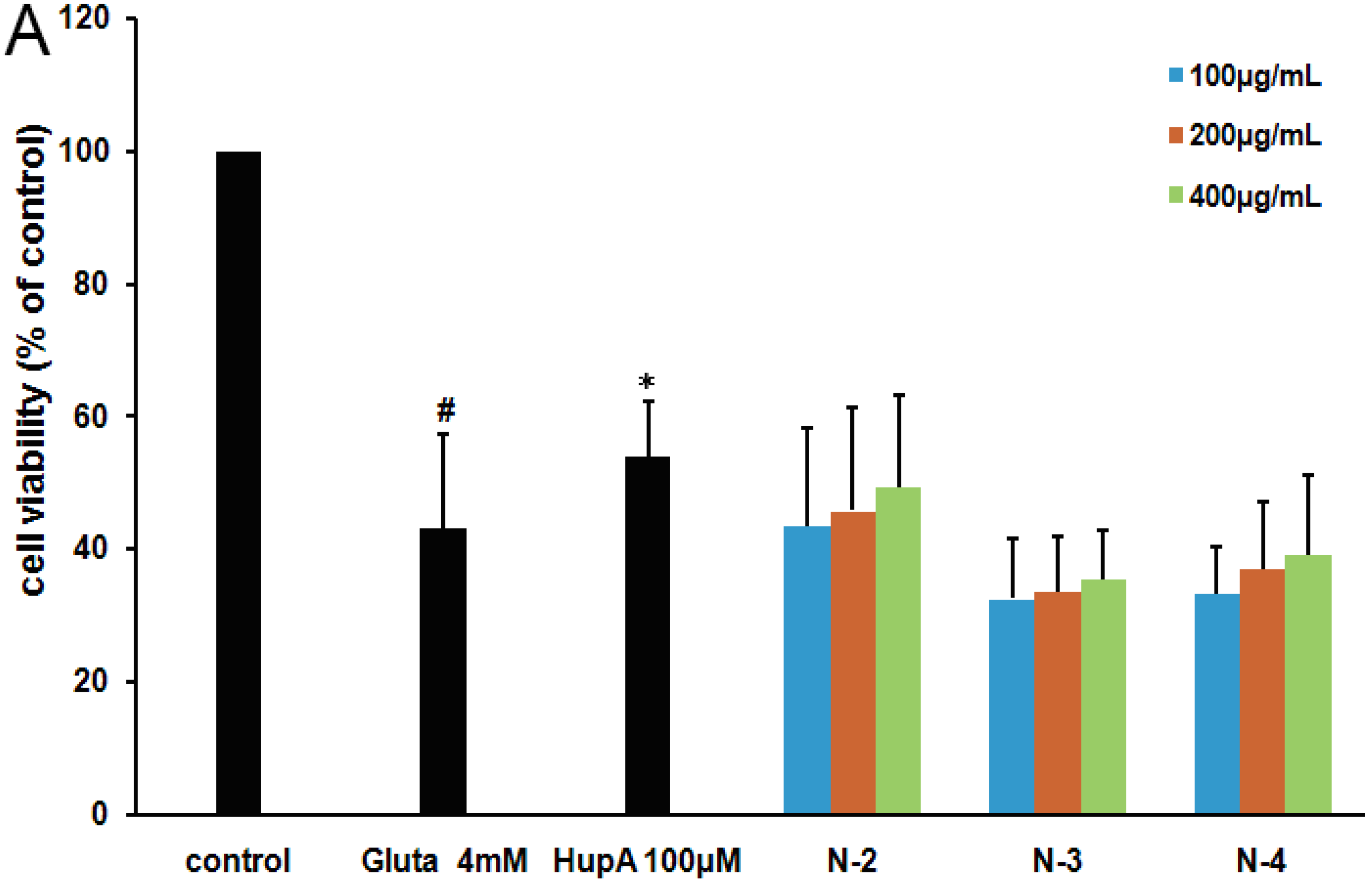
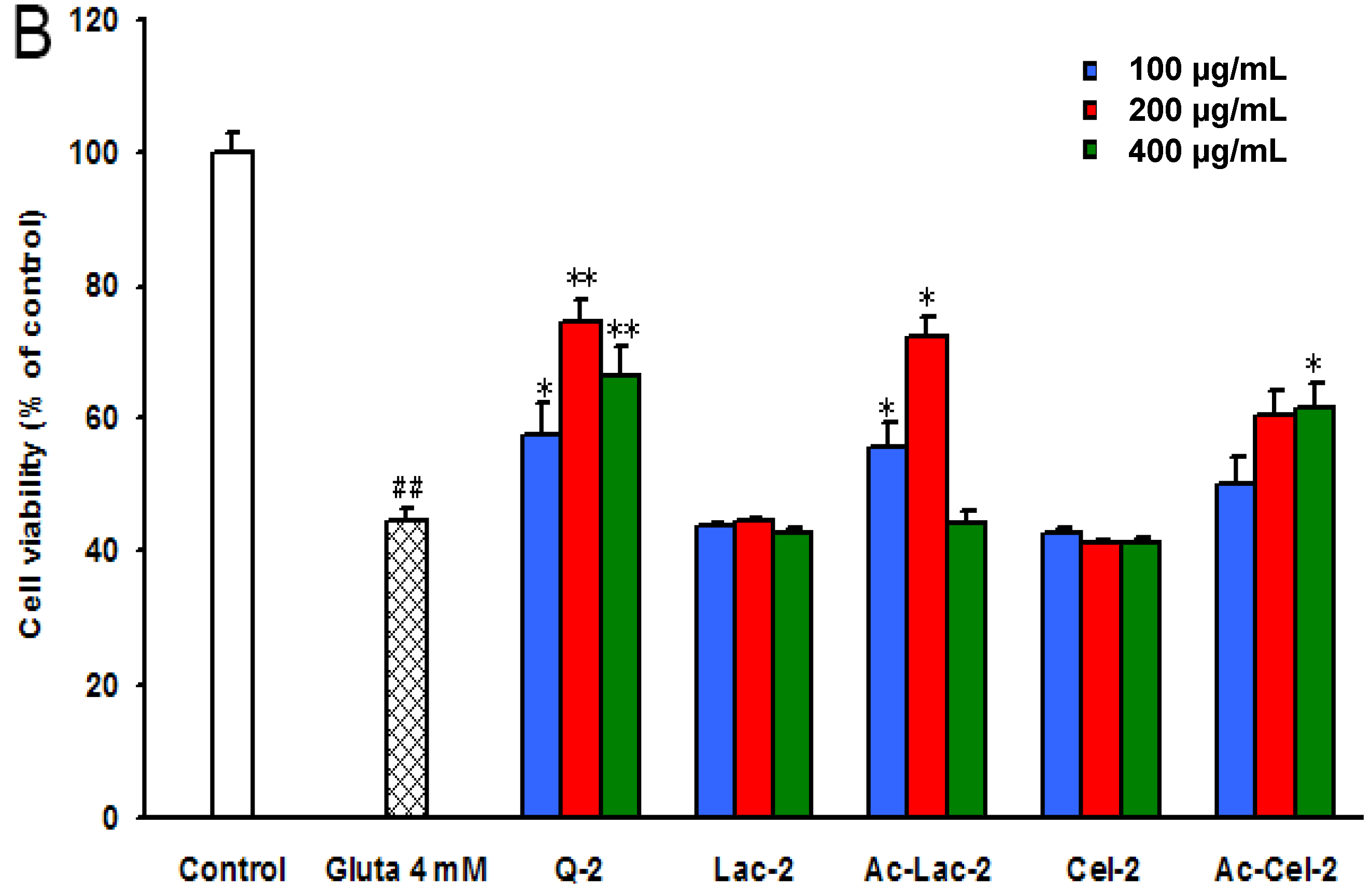
2.4. The Structure-Activity Relationship of Acetylated Chitosan Oligosaccharides


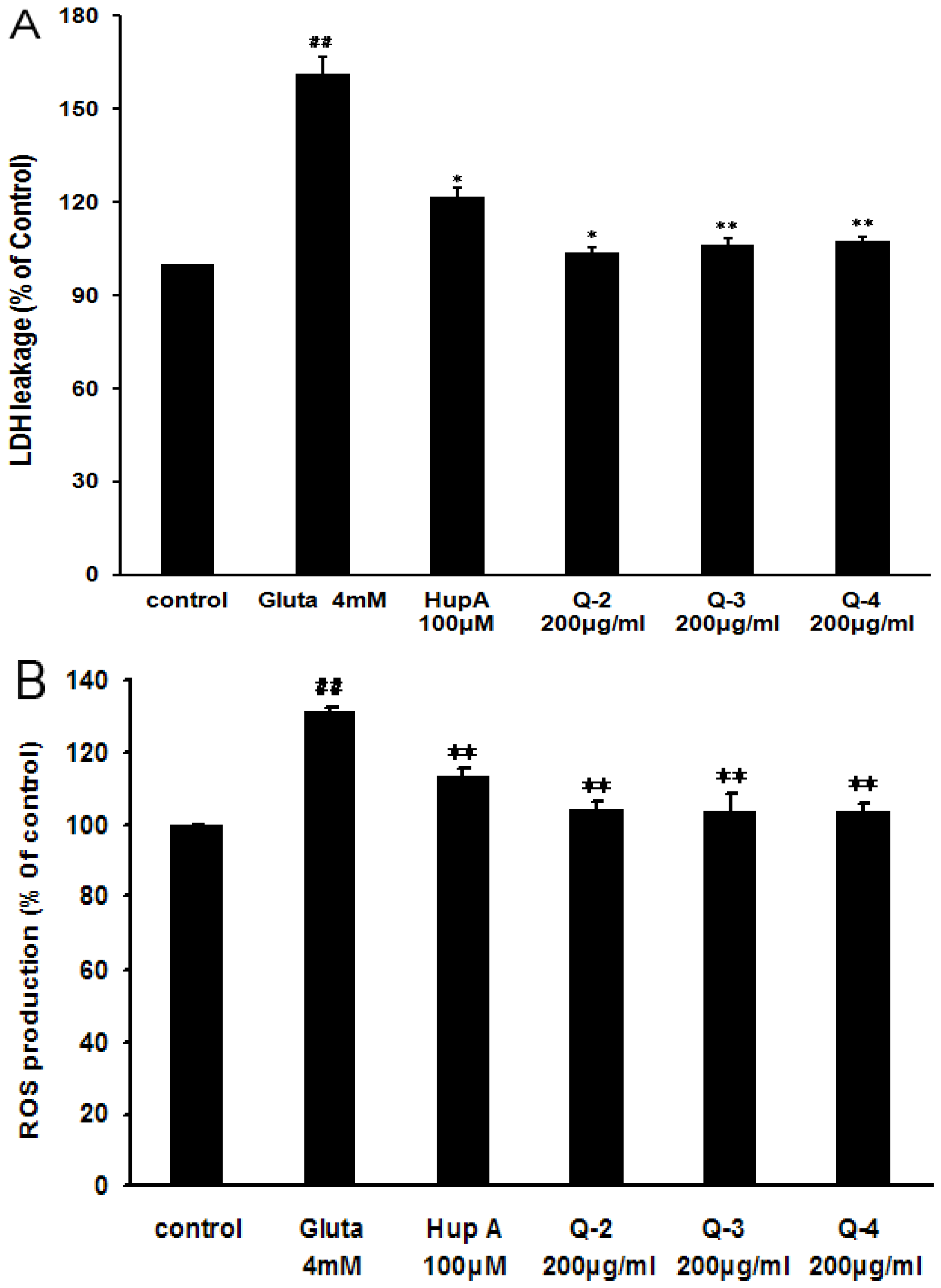
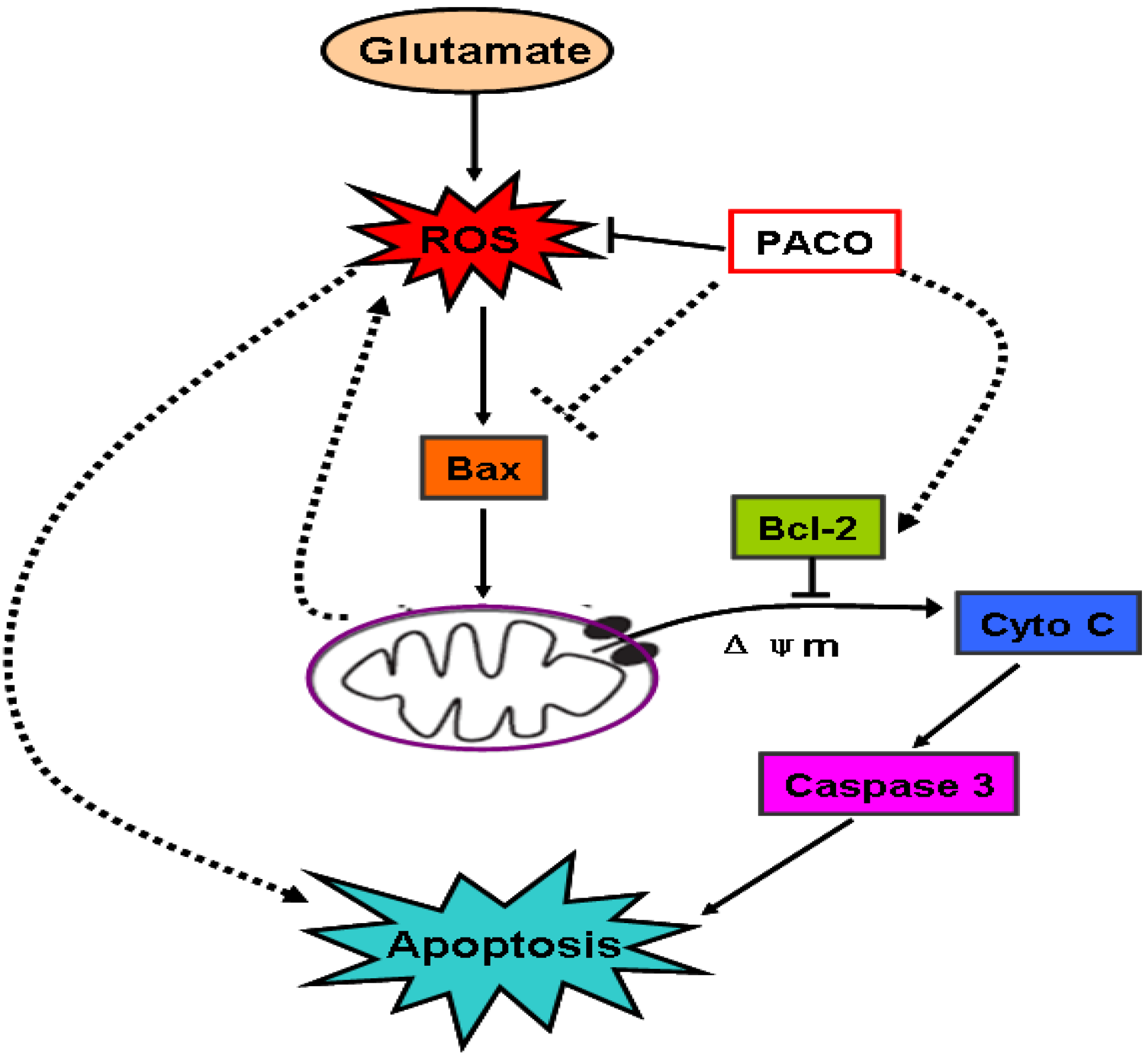
2.5. Effect of Peracetylated Chitosan Oligosaccharides (PACO) on LDH Release and ROS Production

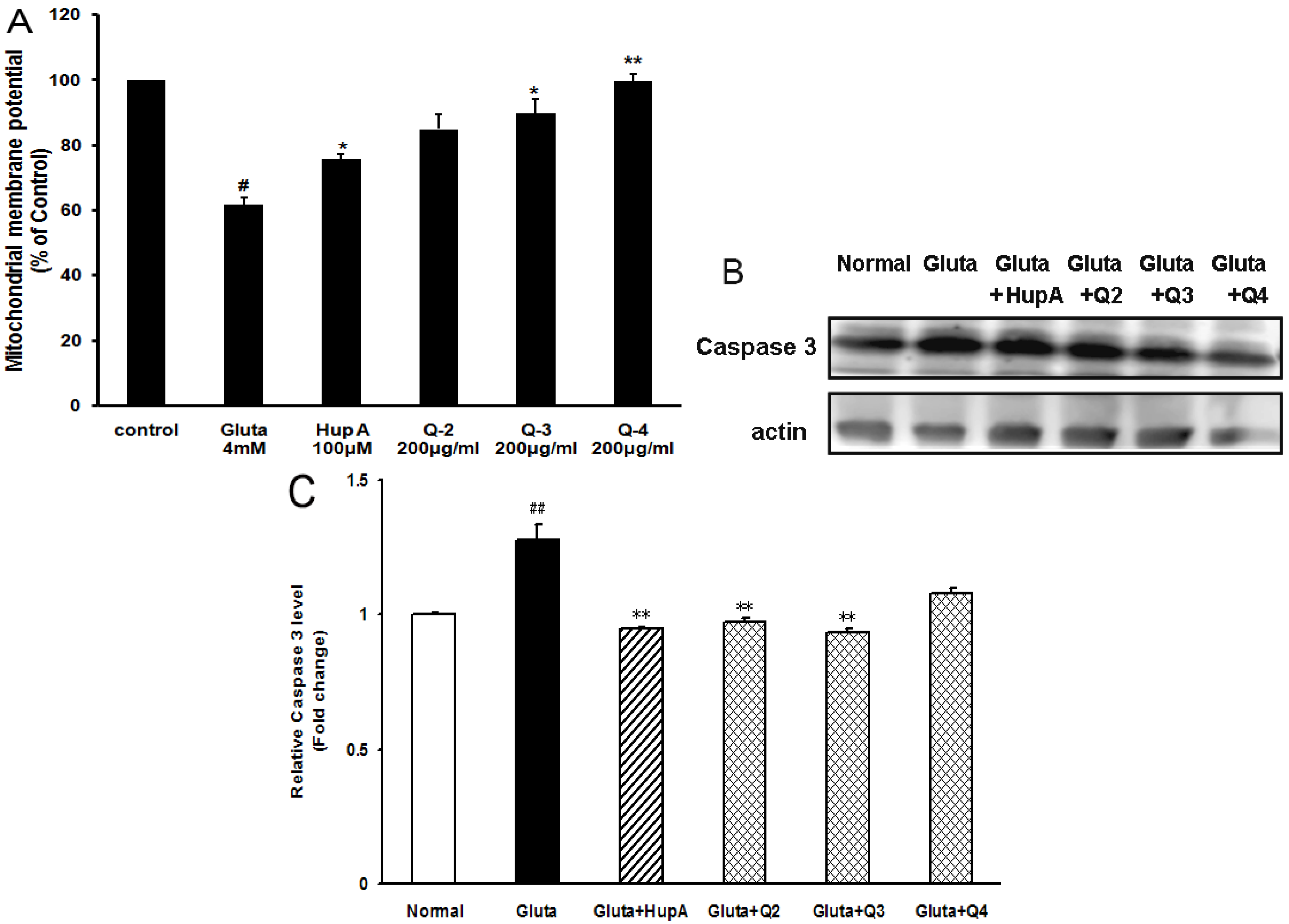
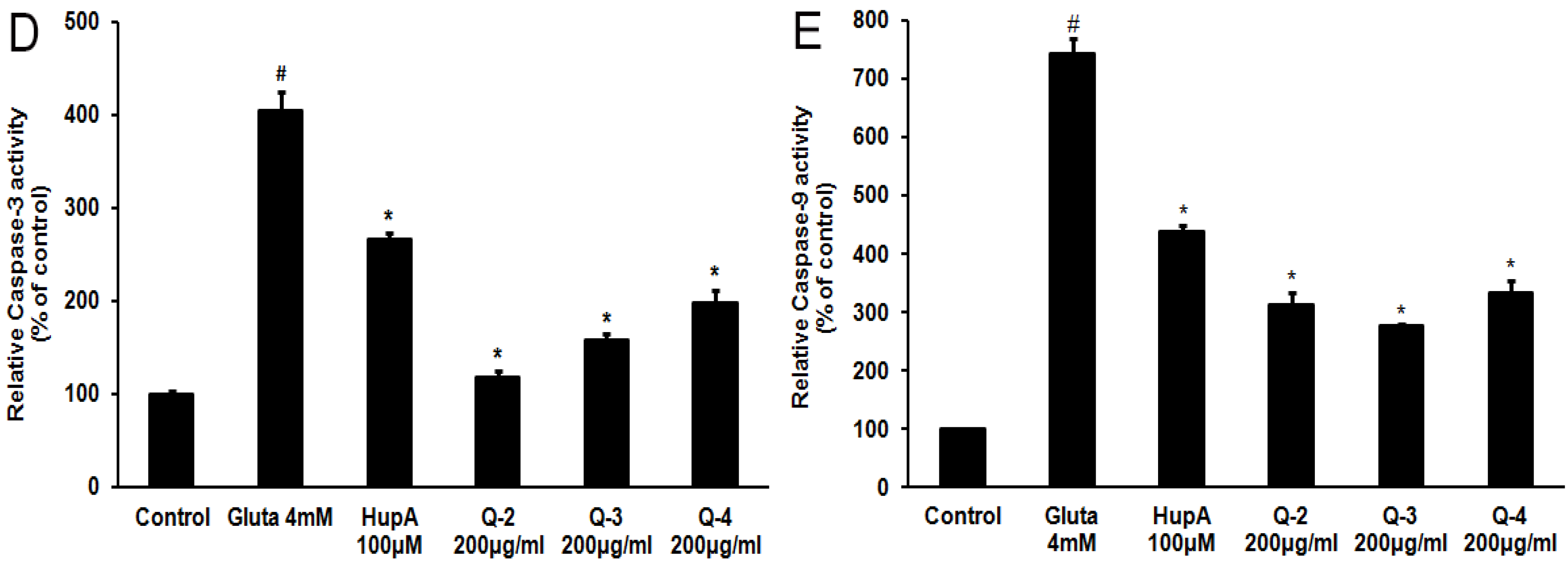
2.6. Effects of PACO on Mitochondrial Membrane Potential and the Activation of Caspase-3 and Caspase-9


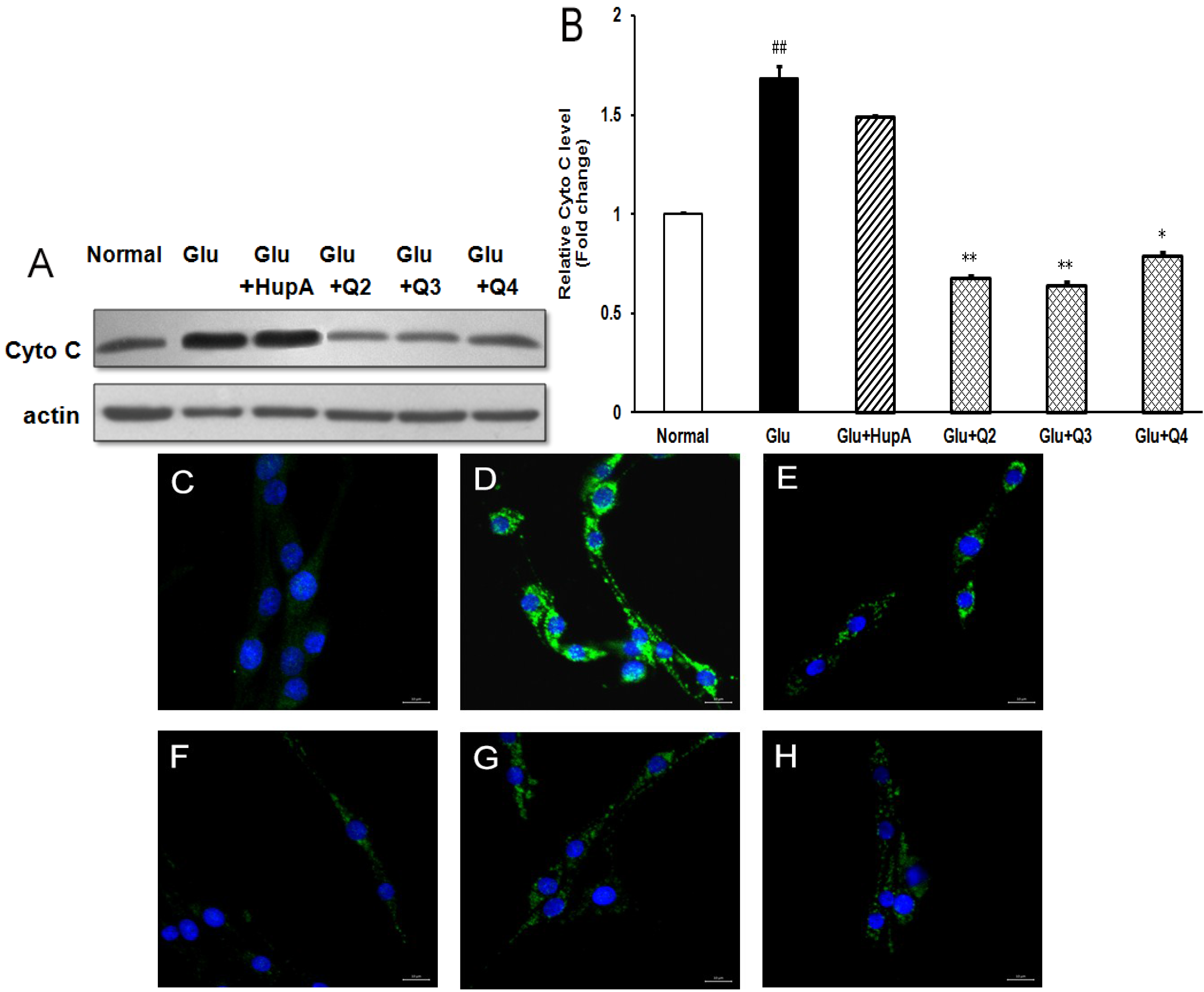
2.7. Effects of PACO on the Cytochrome c (Cyto C) Release from Mitochondria

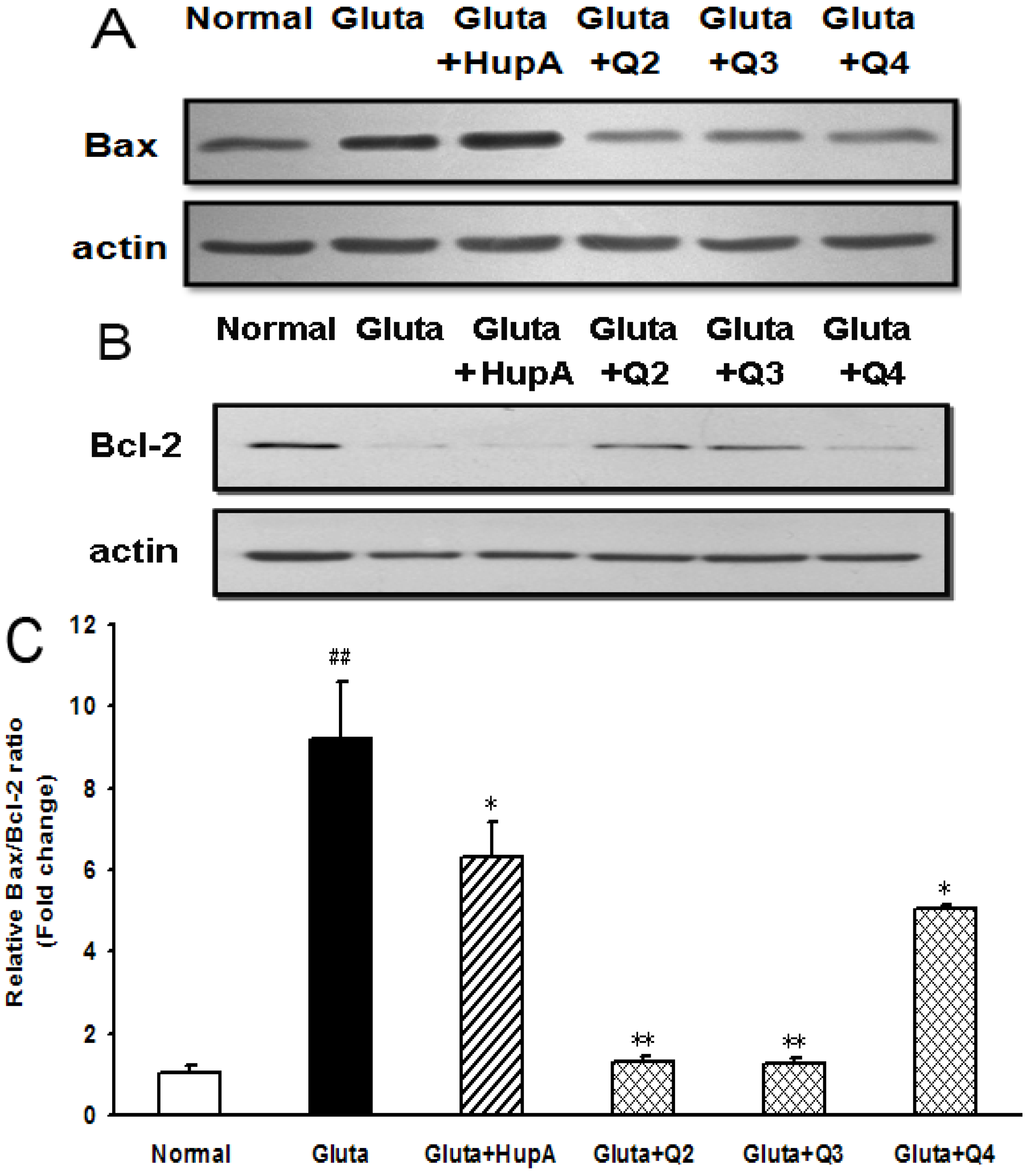
2.8. Effects of PACO on the Protein Expression of Bcl-2 and Bax


3. Experimental Section
3.1. Reagents
3.2. COSs Production and Purification
3.3. PACOs and NACOs Preparation and Purification
3.4. MS and NMR Spectroscopy of Isolated PACOs and NACOs
3.5. Cell Culture
3.6. Determination of Cell Viability
3.7. LDH Release Assay
3.8. Measurement of Intracellular ROS
3.9. Measurement of the Mitochondrial Membrane Potential (MMP)
3.10. Immunofluorescence Assay
3.11. Western Blot Analysis
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tan, J.W.; Tham, C.L.; Israf, D.A.; Lee, S.H.; Kim, M.K. Neuroprotective effects of biochanin A against glutamate induced cytotoxicity in PC12 cells via apoptosis inhibition. Neurochem. Res. 2013, 38, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [PubMed]
- Mao, Q.Q.; Zhong, X.M.; Feng, C.R.; Pan, A.J.; Li, Z.Y.; Huang, Z. Protective effects of paeoniflorin against glutamate-induced neurotoxicity in PC12 cells via antioxidant mechanisms and Ca2+ antagonism. Cell Mol. Neurobiol. 2010, 30, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W. Glutamate neurotoxicity and diseases of the nervous system. Neuron 1988, 1, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.H.; Miyamoto, M.; Sastre, A.; Schnaar, R.L.; Coyle, J.T. Glutamate toxicity in a neuronal cell line involves inhibition of cystine transport leading to oxidative stress. Neuron 1989, 2, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Zablocka, A.; Janusz, M. The two faces of reactive oxygen species. Postep. Hig. Med. Dosw. 2008, 62, 118–124. [Google Scholar]
- Monaghan, D.T.; Bridges, R.J.; Cotman, C.W. The excitatory amino acid receptors: Their classes, pharmacology, and distinct properties in the function of the central nervous system. Annu. Rev. Pharmacol. Toxicol. 1989, 29, 365–402. [Google Scholar] [CrossRef] [PubMed]
- Bleich, S.; Romer, K.; Wiltfang, J.; Kornhuber, J. Glutamate and the glutamate receptor system: A target for drug action. Int. J. Geriatr. Psychiatry 2003, 18, S33–S40. [Google Scholar] [CrossRef] [PubMed]
- Penugonda, S.; Mare, S.; Goldstein, G.; Banks, W.A.; Ercal, N. Effects of N-acetylcysteine amide (NACA), a novel thiol antioxidant against glutamate-induced cytotoxicity in neuronal cell line PC12. Brain Res. 2005, 1056, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Penugonda, S.; Mare, S.; Lutz, P.; Banks, W.A.; Ercal, N. Potentiation of lead-induced cell death in PC12 cells by glutamate: Protection by N-acetylcysteine amide (NACA), a novel thiol antioxidant. Toxicol. Appl. Pharmacol. 2006, 216, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, B.; Dluzen, D.E.; Jin, Y. Protective effects of ginsenoside Rg2 against glutamate-induced neurotoxicity in PC12 cells. J. Ethnopharmacol. 2007, 111, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Wu, L.L.; Pan, Y.; Song, Q.; Ran, C.L.; Liu, S.K. The effect of Jiaweisinisan on cAMP response element binding protein and phosphorylation in PC12 cells injured by Corticosterone and Glutamate. Chin. Pharmacol. Bull. 2009, 25, 270–274. [Google Scholar]
- Li, Y.F.; Zhang, Y.Z.; Liu, Y.Q.; Wang, H.L.; Cao, J.B.; Guan, T.T.; Luo, Z.P. Inhibition of N-methyl-d-aspartate receptor function appears to be one of the common actions for antidepressants. J. Psychopharmacol. 2006, 20, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, W.; Liu, P.; Cheng, Q.; Tahi, T.; Gu, W.; Li, B. Chitosan modification and pharmaceutical/biomedical applications. Mar. Drugs 2010, 8, 1962–1987. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.K. Neuroprotective Properties of Chitosan and Its Derivatives. Mar. Drugs 2010, 8, 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Park, J.-S.; Kim, S.-K.; Ahn, C.-B.; Je, J.-Y. Chitooligosaccharides suppress the level of protein expression and acetylcholinesterase activity induced by A[beta]25–35 in PC12 cells. Bioorganic Med. Chem. Lett. 2009, 19, 860–862. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Ngo, D.-N.; Kim, S.-K. Acetylcholinesterase inhibitory activity of novel chitooligosaccharide derivatives. Carbohydr. Polym. 2009, 78, 869–872. [Google Scholar] [CrossRef]
- Zhou, S.; Yang, Y.; Gu, X.; Ding, F. Chitooligosaccharides protect cultured hippocampal neurons against glutamate-induced neurotoxicity. Neurosci. Lett. 2008, 444, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.-X.; Li, C.-X.; Wang, S.-X.; Zhao, X.; Guan, H.-S. Preparation and analysis of Chitooligosaccharide isomers with different degree. Chin. J. Mar. Drugs 2013, 32, 21–27. [Google Scholar]
- Wang, J.; Li, Y.-X.; Song, N.; Guan, H.-S. Preparation and Characterization of Chito-oligosaccharide and Peracetylated-chito-oligosaccharides. Period. Ocean Univers. China 2005, 35, 994–1000. [Google Scholar]
- Nociari, M.M.; Shalev, A.; Benias, P.; Russo, C. A novel one-step, highly sensitive fluorometric assay to evaluate cell-mediated cytotoxicity. J. Immunol. Methods 1998, 213, 157–167. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.J.; Zhang, M.; Hao, C.; Wang, W.; Qiu, P.J.; Wan, S.B.; Zhang, L.J.; Jiang, T. Different cytotoxicities and cellular localizations of novel quindoline derivatives with or without boronic acid modifications in cancer cells. Chem. Commun. 2013, 49, 8516–8518. [Google Scholar] [CrossRef]
- McElnea, E.M.; Quill, B.; Docherty, N.G.; Irnaten, M.; Siah, W.F.; Clark, A.F.; O’Brien, C.J.; Wallace, D.M. Oxidative stress, mitochondrial dysfunction and calcium overload in human lamina cribrosa cells from glaucoma donors. Mol. Vis. 2011, 17, 1182–1191. [Google Scholar] [PubMed]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ali, S.F.; McKee, M.; Bondy, S.C. Organometal-induced increases in oxygen reactive species: The potential of 2′,7′-dichlorofluorescin diacetate as an index of neurotoxic damage. Toxicol. Appl. Pharmacol. 1990, 104, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Noshita, N.; Lewen, A.; Gashe, Y.; Ferrand-Drake, M.; Fujimura, M.; Morita-Fujimura, Y.; Chan, P.H. Overexpression of copper/zinc superoxide dismutase in transgenic rats protects vulnerable neurons against ischemia damage by blocking the mitochondrial pathway of caspase activation. J. Neurosci. 2002, 22, 209–217. [Google Scholar] [PubMed]
- Brown, J.R.; Fuster, M.M.; Li, R.X.; Varki, N.; Glass, C.A.; Esko, J.D. A Disaccharide-Based Inhibitor of Glycosylation Attenuates MetastaticTumor Cell Dissemination. Clin. Cancer Res. 2006, 12, 2894–2901. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Jeong, H.Y.; Lee, H.K.; Kim, S.; Hwang, B.Y.; Bae, K.; Seong, Y.H. Neuroprotection of the leaf and stem of Vitis amurensis and their active compounds against ischemic brain damage in rats and excitotoxicity in cultured neurons. Phytomedicine 2011, 19, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Liu, H.; Jiao, H.; Wang, L.; Chen, L.; Liang, J.; Zhao, M.; Zhang, X. Neuroprotective effect of ginkgolide K on glutamate-induced cytotoxicity in PC 12 cells via inhibition of ROS generation and Ca2+ influx. NeuroToxicology 2012, 33, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kanki, R.; Nakamizo, T.; Yamashita, H.; Kihara, T.; Sawada, H.; Uemura, K.; Kawamata, J.; Shibasaki, H.; Akaike, A.; Shimohama, S. Effects of mitochondrial dysfunction on glutamate receptor-mediated neurotoxicity in cultured rat spinal motor neurons. Brain Res. 2004, 1015, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Yamamoto, H.; Atta, M.S.; Mahmoud, S.; Oh-Hashi, K.; Kiuchi, K. Chloroquine inhibits glutamate-induced death of a neuronal cell line by reducing reactive oxygen species through sigma-1 receptor. J. Neurochem. 2011, 119, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chua, K.W.; Chua, C.C.; Yu, H.; Pei, A.; Chua, B.H.; Hamdy, R.C.; Xu, X.; Liu, C.F. Antioxidant activity of 7,8-dihydroxyflavone provides neuroprotection against glutamate-induced toxicity. Neurosci. Lett. 2011, 499, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; Yin, X.M.; Wang, K.; Wei, M.C.; Jockel, J.; Milliman, C.; Erdjument-Bromage, H.; Tempst, P.; Korsmeyer, S.J. Caspase cleaved BID targets mitochondria and is required for cytochrome c release, while BCL-XL prevents this release but not tumor necrosis factor-R1/Fas death. J. Biol. Chem. 1999, 274, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Osawa, T. Astaxanthin protects neuronal cells against oxidative damage and is a potent candidate for brain food. Forum Nutr. 2009, 61, 129–135. [Google Scholar] [PubMed]
- Lan, A.P.; Xiao, L.C.; Yang, Z.L.; Yang, C.T.; Wang, X.Y.; Chen, P.X.; Gu, M.F.; Feng, J.Q. Interaction between ROS and p38MAPK contributes to chemical hypoxia-induced injuries in PC12 cells. Mol. Med. Rep. 2011, 5, 250–255. [Google Scholar] [PubMed]
- Tao, L.; Li, X.; Zhang, L.; Tian, J.; Li, X.; Sun, X.; Li, X.; Jiang, L.; Zhang, X.; Chen, J. Protective effect of tetrahydroxystilbene glucoside on 6-OHDA-induced apoptosis in PC12 cells through the ROS-NO pathway. PLoS One 2011, 6, e26055. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yang, N.; Liu, Y.; Li, Q.; Zuo, P. The neuroprotective effects of phytoestrogen azearalanol on b-amyloid-induced toxicity in differentiated PC-12 cells. Eur. J. Pharmacol. 2011, 670, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Lee, S.J.; Moon, S.H.; Jeon, B.T.; Kim, B.; Park, T.K.; Han, J.S.; Park, P.J. Neuroprotective effects of a novel peptide purified from venison protein. J. Microbiol. Biotechnol. 2010, 20, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, O.V.; Yuzbasioglu, M.F.; Ciralik, H.; Kurutas, E.B.; Yonden, Z.; Aydin, M.; Bulbuloglu, E.; Semerci, E.; Goksu, M.; Atli, Y.; et al. Resveratrol, a natural antioxidant, attenuates intestinal ischemia/reperfusion injury in rats. Tohoku J. Exp. Med. 2009, 218, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Zhu, G.; Yang, S.; Wang, X.; Cheng, H.; Wang, F.; Li, X.X.; Li, Q.L. Paeonol prevents excitotoxicity in rat pheochromocytoma PC12 cells via downregulation of ERK activation and inhibition of apoptosis. Planta Med. 2011, 77, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ma, R.; Huang, C.; Tang, Q.; Fu, Q.; Liu, H.; Hu, B.; Xiang, J. Protective effect of erythropoietin on b-amyloid-induced PC12 cell death through antioxidant mechanisms. Neurosci. Lett. 2008, 442, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.C.; Mong, M.C.; Yin, A.C. Antioxidative and anti-inflammatory, neuroprotective effects of astaxanthin and canthaxanthin in nerve growth factor differentiated PC12 cells. J. Food Sci. 2009, 74, H225–H231. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Hu, C.P.; Xu, K.P.; Yuan, Q.; Li, F.S.; Zou, H.; Tan, G.S.; Li, Y.J. Protective effect of selaginellin on glutamate-induced cytotoxicity and apoptosis in differentiated PC12 cells. Naunyn Schmiedebergs Arch. Pharmacol. 2010, 381, 73–81. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, C.; Gao, L.; Zhang, Y.; Wang, W.; Yu, G.; Guan, H.; Zhang, L.; Li, C. Acetylated Chitosan Oligosaccharides Act as Antagonists against Glutamate-Induced PC12 Cell Death via Bcl-2/Bax Signal Pathway. Mar. Drugs 2015, 13, 1267-1289. https://doi.org/10.3390/md13031267
Hao C, Gao L, Zhang Y, Wang W, Yu G, Guan H, Zhang L, Li C. Acetylated Chitosan Oligosaccharides Act as Antagonists against Glutamate-Induced PC12 Cell Death via Bcl-2/Bax Signal Pathway. Marine Drugs. 2015; 13(3):1267-1289. https://doi.org/10.3390/md13031267
Chicago/Turabian StyleHao, Cui, Lixia Gao, Yiran Zhang, Wei Wang, Guangli Yu, Huashi Guan, Lijuan Zhang, and Chunxia Li. 2015. "Acetylated Chitosan Oligosaccharides Act as Antagonists against Glutamate-Induced PC12 Cell Death via Bcl-2/Bax Signal Pathway" Marine Drugs 13, no. 3: 1267-1289. https://doi.org/10.3390/md13031267
APA StyleHao, C., Gao, L., Zhang, Y., Wang, W., Yu, G., Guan, H., Zhang, L., & Li, C. (2015). Acetylated Chitosan Oligosaccharides Act as Antagonists against Glutamate-Induced PC12 Cell Death via Bcl-2/Bax Signal Pathway. Marine Drugs, 13(3), 1267-1289. https://doi.org/10.3390/md13031267