
Toxin Profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese Coast, as Determined by Liquid Chromatography Tandem Mass Spectrometry




Abstract
:1. Introduction

2. Results and Discussion
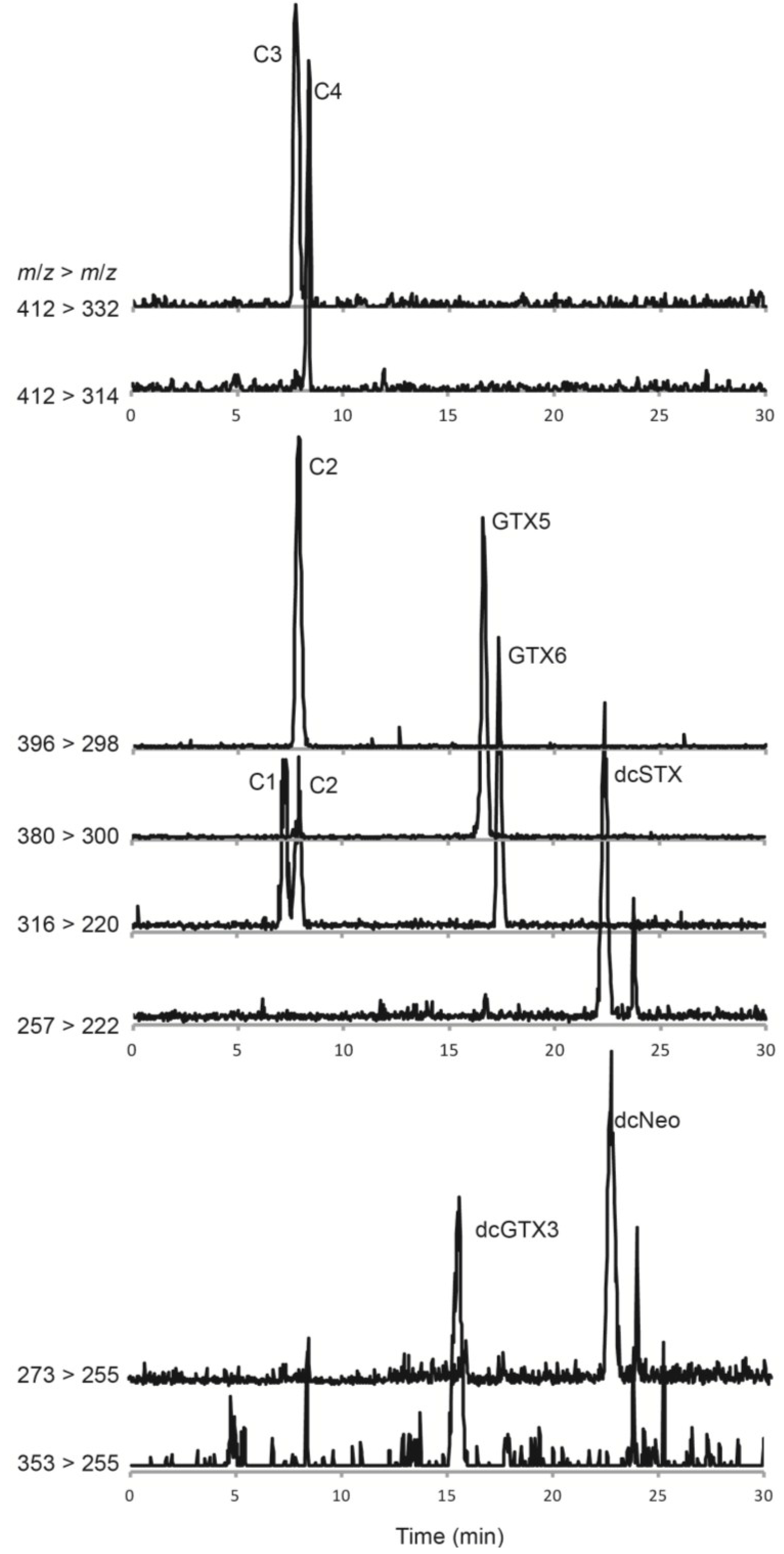
2.1. Identification of “Classic” PSTs in G. catenatum

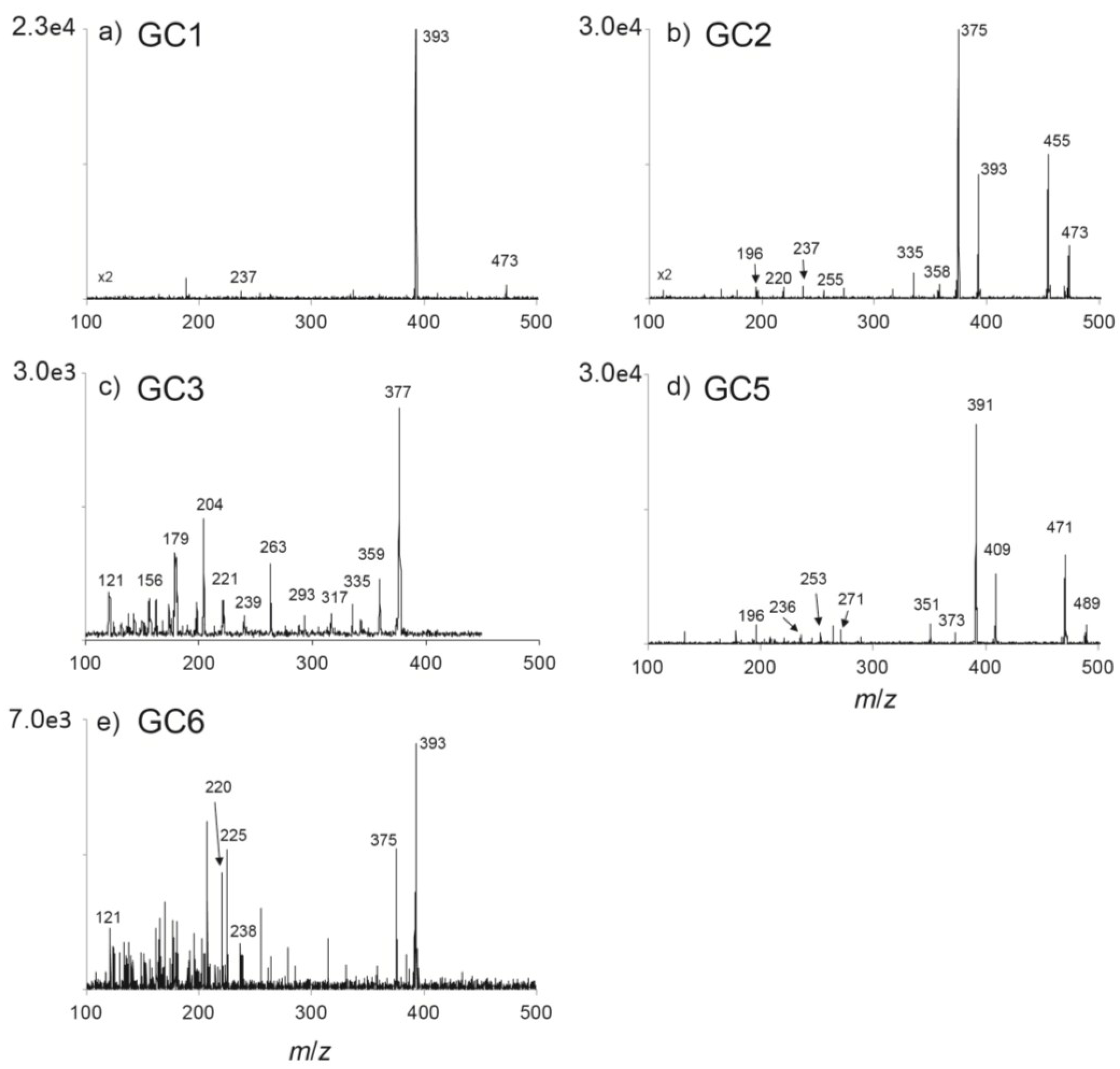
2.2. Identification of Hydroxybenzoate PSTs in Portuguese G. catenatum



2.3. Understanding Gymnodinium catenatum Bloom Hiatus in Portuguese Waters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Toxin Cell Quota (fmol/Cell) ^ | Toxin Profile (mol%) | |||
|---|---|---|---|---|---|
| This Study | This Study | Negri et al. (2007) | |||
| Culture | Seawater | Culture | Seawater | Culture (1989) | |
| C1 + 2 | 6.5 | 4.3 | 21.0 | 22.1 | 14.5 |
| C3 + 4 | 0.5 | 0.4 | 1.6 | 2.0 | 6.0 |
| GTX2 + 3 | nd | nd | nd | nd | 1.0 |
| GTX5 | 3.4 | 6.9 | 11.0 | 35.2 | 12.9 |
| GTX6 | 1.2 | nd | 3.8 | nd | 5.9 |
| dcGTX2 + 3 | 0.3 | nd | 0.8 | nd | 4.0 |
| dcSTX | 2.2 | nd | 7.2 | nd | 2.5 |
| dcNeo | 1.1 | nd | 3.4 | nd | nd |
| STX | nd | nd | nd | nd | 0.4 |
| GC1 + 2 | 5.9 | 6.2 | 19.0 | 31.6 | 35.3 |
| GC3 | 5.0 | 1.6 | 16.2 | 8.3 | 17.6 |
| GC4 * + 5 | 1.7 | 0.2 | 5.6 | 0.8 | na |
| GC6 | 3.2 | nd | 10.3 | nd | na |
2.4. Management Implications
3. Experimental Section
3.1. Reagents and Standards
3.2. Sample Collection and Culture
3.3. Chemical Extraction of PSTs
3.4. Liquid Chromatography Tandem Mass Spectrometry
| Toxin | Transition Ion Pair (m/z) | Collision Energy (eV) | Declustering Potential (V) |
|---|---|---|---|
| dcSTX | 257 > 222 | 19 | 35 |
| 257 > 239 | 10 | 30 | |
| dcNeo | 273 > 255 | 20 | 35 |
| 273 > 225 | 25 | 35 | |
| STX | 300 > 204 | 20 | 35 |
| GTX3 | 316 > 220 | 20 | 35 |
| dcGTX3 | 353 > 255 | 20 | 35 |
| GTX5 | 380 > 300 | 14 | 20 |
| 300 > 204 | 20 | 35 | |
| C2 | 396 > 298 | 10 | 40 |
| 396 > 316 | 15 | 45 | |
| C1 | 316 > 220 | 20 | 35 |
| 396 > 316 | 15 | 45 | |
| GTX6 | 316 > 220 | 20 | 35 |
| 396 > 316 | 15 | 45 | |
| C4 | 412 > 314 | 10 | 40 |
| 412 > 332 | 15 | 45 | |
| C3 | 412 > 332 | 15 | 45 |
| GC1 | 393 > 320 | 15 | 20 |
| 473 > 393 | 15 | 20 | |
| GC2 | 473 > 375 | 15 | 20 |
| 473 > 455 | 15 | 20 | |
| GC3 | 377 > 204 | 25 | 40 |
| 377 > 359 | 20 | 30 | |
| GC4 | 489 > 409 | 20 | 30 |
| GC5 | 489 > 391 | 20 | 30 |
| 489 > 471 | 15 | 20 | |
| GC6 | 393 > 220 | 20 | 40 |
| 393 > 375 | 15 | 20 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Estrada, M.; Sanchez, F.J.; Fraga, S. Gymnodinium catenatum Graham en las rias gallegas (NO España). Investig. Pesq. 1984, 48, 31–40. [Google Scholar]
- Franca, S.; Almeida, J.F. Paralytic shellfish poisons in bivalve molluscs on the Portuguese coast caused by a bloom of the dinoflagellate Gymnodinium catenatum. In Red Tides: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 93–96. [Google Scholar]
- Mee, L.D.; Espinosa, M.; Diaz, G. Paralytic shellfish poisoning with a Gymnodinium catenatum red-tide on the Pacific coast of Mexico. Mar. Environ. Res. 1986, 19, 77–92. [Google Scholar] [CrossRef]
- Llewellyn, L.E.; Dodd, M.J.; Robertson, A.; Ericsona, G.; de Koning, C.; Negri, A.P. Post-mortem analysis of samples from a human victim of a fatal poisoning caused by the xanthid crab, Zosimus aeneus. Toxicon 2002, 40, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Bravo, M.C.; Lagos, M.; Lagos, N. Paralytic shellfish poisoning: post-mortem analysis of tissue and body fluid samples from human victims in the Patagonia fjords. Toxicon 2004, 43, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.M.; Carvalho, M.; Mestre, T.; Ferreira, J.J.; Coelho, M.; Peralta, R.; Vale, P. Paralytic shellfish poisoning due to ingestion of Gymnodinium catenatum contaminated cockles—Application of the AOAC HPLC Official Method. Toxicon 2012, 59, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Nishiyama, A. Actions of saxitoxin on peripheral neuromuscular systems. J. Physiol. 1965, 180, 50–66. [Google Scholar] [PubMed]
- Ritchie, J.M.; Rogart, R.B. The binding of saxitoxin and tetrodotoxin to excitable tissue. Rev. Physiol. Biochem. Pharmacol. 1977, 79, 1–49. [Google Scholar] [PubMed]
- Narahashi, T.; Moore, J.W. Neuroactive agents and nerve membrane conductances. J. Gen. Physiol. 1968, 51, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Lagos, M.; Andrinolo, D. Paralytic shellfish poisoning (PSP): Toxicology and kinetics. In Seafood and Freshwater Toxins; Botana, L.M., Ed.; Marcel Dekker: New York, NY, USA, 2000; pp. 203–215. [Google Scholar]
- Genenah, A.A.; Shimizu, Y. Specific toxicity of paralytic shellfish poisons. J. Agric. Food. Chem. 1981, 29, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y. Postcolumn derivatization liquid-chromatography method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar]
- Catterall, W.A. Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Negri, A.P.; Stirling, D.; Quilliam, M.; Blackburn, S.; Bolch, C.; Burton, I.; Eaglesham, G.; Thomas, K.; Walter, J.; Willis, R. Three novel hydroxybenzoate saxitoxin analogues isolated from the dinoflagellate Gymnodinium catenatum. Chem. Res. Toxicol. 2003, 16, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.; Negri, A.; Quilliam, M. High affinity for the rat brain sodium channel of newly discovered hydroxybenzoate saxitoxin analogues from the dinoflagellate Gymnodinium catenatum. Toxicon 2004, 43, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Negri, A.P.; Bolch, C.J.S.; Geier, S.; Green, D.H.; Park, T.; Blackburn, S.I. Widespread presence of hydrophobic paralytic shellfish toxins in Gymnodinium catenatum. Harmful Algae 2007, 6, 774–780. [Google Scholar] [CrossRef]
- Vale, P. Complex profiles of hydrophobic shellfish poisoning compounds in Gymnodinium catenatum identified by liquid chromatography with fluorescence detection and mass spectrometry. J. Chromatogr. A 2008, 1195, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M. Red tides. Sci. Am. 1994, 271, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Moita, T.M.; Oliveira, P.B.; Mendes, J.C.; Palma, A.S. Distribution of chlorophyll a and Gymnodinium catenatum associated with coastal upwelling plumes off central Portugal. Acta Oecol. 2003, 24, S125–S132. [Google Scholar] [CrossRef]
- Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Gomes, S.S.; Sampayo, M.A.M. Two decades of marine biotoxin monitoring in bivalves from Portugal (1986–2006): A review of exposure assessment. Harmful Algae 2008, 7, 11–25. [Google Scholar] [CrossRef]
- Moita, M.T.; Palma, S.; Oliveira, P.B.; Vidal, T.; Silva, A.; Vilarinho, M.G. The return of Gymnodinium catenatum after 10 years: Bloom initiation and transport off the Portuguese coast. In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; Intergovernmental Oceanographic Commission (IOC) of UNESCO: Copenhagen, Denmark, 2008; p. 242. [Google Scholar]
- Hall, S.; Strichartz, G.; Moczydlowski, E.; Ravindran, A.; Reichardt, P.B. The saxitoxins. In Marine Toxins: Origin, Structure and Molecular Pharmacology; Hall, S., Strichartz, G., Eds.; ACS Symposium Series: Washington, DC, USA, 1990; Volume 418, pp. 29–65. [Google Scholar]
- Krock, B.; Seguel, C.G.; Cembella, A.D. Toxin profile of Alexandrium catenella from Chilean coast as determined by liquid chromatography with fluorescence detection and liquid chromatography coupled with tandem mass spectrometry. Harmful Algae 2007, 6, 734–744. [Google Scholar] [CrossRef] [Green Version]
- Graham, H.W. Gymnodinium catenatum, a new dinoflagellate from the Gulf of California. Trans. Am. Microsc. Soc. 1943, 62, 259–261. [Google Scholar] [CrossRef]
- Hallegraef, G.M.; Blackburn, S.I.; Doblin, M.A.; Bolch, C.J.S. Global toxicology, ecophysiology and population relationships of the chainforming PST dinoflagellate Gymnodinium catenatum. Harmful Algae 2012, 14, 130–143. [Google Scholar] [CrossRef]
- Amorim, A.; Dale, B. Historical cyst record as evidence for the recent introduction of the dinoflagellate Gymnodinium catenatum in the north-eastern Atlantic. Afr. J. Mar. Sci. 2006, 28, 193–197. [Google Scholar] [CrossRef]
- Ribeiro, S.; Amorim, A.; Andersen, T.J.; Abrantes, F.; Ellegaard, M. Reconstructing the history of an invasion: the toxic phytoplankton species Gymnodinium catenatum in the Northeast Atlantic. Biol. Invas. 2012, 14, 969–985. [Google Scholar] [CrossRef]
- Moita, M.T.; Vilarinho, M.G.; Palma, A.S. On the variability of Gymnodinium catenatum Graham blooms in Portuguese waters. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO, Santiago de Compostela: Amsterdam, The Netherlands, 1998; pp. 118–121. [Google Scholar]
- Franca, S.; Alvito, P.; Sousa, I.; Gago, A.; Rodríguez-Vásquez, J.A.; Leão, J.M.; Comesaña, M.; Thibault, P.; Burdaspal, P.; Bustos, J.; et al. The toxin profile of some PSP toxin producing dinoflagellates occurring in Portuguese coastal waters as determined by alternative analytical methods. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 519–522. [Google Scholar]
- Flynn, K.J.; Flynn, K; John, E.H.; Reguera, B.; Reyero, M.I.; Franco, J.M. Changes in toxins, intracellular and dissolved free amino acids of the toxic dinoflagellate Gymnodinium catenatum in response to changes in inorganic nutrients and salinity. J. Plank. Res. 1996, 18, 2093–2111. [Google Scholar] [CrossRef]
- Band-Schmidt, C.; Bustillos-Guzman, J.; Morquecho, L.; Gárate-Lizárraga, I.; Alonso-Rodríguez, R.; Reyes-Salinas, A.; Erler, K.; Luckas, B. Variations of PSP toxin profiles during different growth phases in Gymnodinium catenatum (Dinophyceae) strains isolated from three locations in the Gulf of California, Mexico. J. Phycol. 2006, 42, 757–768. [Google Scholar] [CrossRef]
- Oshima, Y.; Blackburn, S.I.; Hallegraeff, G.M. Comparative study on paralytic shellfish toxin profiles of the dinoflagellate Gymnodinium catenatum from three different countries. Mar. Biol. 1993, 116, 471–476. [Google Scholar] [CrossRef]
- Negri, A.P.; Bolch, C.J.S.; Blackburn, S.I.; Dickman, M.; Llewellyn, L.E.; Méndez, S. Paralytic shellfish toxins in Gymnodinium catenatum strains from six different countries. In Harmful Algal Blooms 2000; Hallegraef, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; pp. 210–213. [Google Scholar]
- Anderson, D.M.; Jacobson, D.M.; Bravo, I.; Wrenn, J.H. The unique, microreticulate cyst of the naked dinoflagellate Gymnodinium catenatum. J. Phycol. 1998, 24, 255–262. [Google Scholar] [CrossRef]
- Amorim, A.; Dale, B.; Godinho, R.; Brotas, V. Gymnodinium catenatum-like cysts (Dinophyceae) in recent sediments from the coast of Portugal. Phycologia 2002, 40, 572–582. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar] [PubMed]
- European Commission (EC) Regulation No. 1664/2006 of the European Parliament and of the Council of 6 November 2006. Off. J. Eur. Commun. 2006, L320, 13–45.
- Perez, S.; Vale, C.; Botana, A.M.; Alonso, W.; Vieytes, M.R.; Botana, L.M. Determination of toxicity equivalent factors for paralytic shellfish toxins by electrophysiological measurements in cultured neurons. Chem. Res. Toxicol. 2001, 24, 1153–1157. [Google Scholar] [CrossRef]
- Costa, P.R.; Moita, T.; Rodrigues, S.M. Estimating the contribution of N-sulfocarbamoyl paralytic shellfish toxin analogs GTX6 and C3+4 to the toxicity of mussels (Mytilus galloprovincialis) over a bloom of Gymnodinium catenatum. Harmful Algae 2014, 31, 35–40. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, P.; Guillherme, S.; Barata, M.; Santos, M.A.; Pacheco, M.; Pousão-Ferreira, P. Hydroxybenzoate paralytic shellfish toxins induce transient GST activity depletion and chromosomal damage in white seabream (Diplodus sargus). Mar. Environ. Res. 2012, 79, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Vale, P. Fate of benzoate paralytic shellfish poisoning toxins from Gymnodinium catenatum in shellfish and fish detected by pre-column oxidation and liquid chromatography with fluorescence detection. J. Chromatogr. A 2008, 1190, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M.; Fire, S.E.; Leighfield, T.A.; Mikulski, C.M.; Doucette, G.J. Determination of paralytic shellfish toxins in shellfish by receptor binding assay: collaborative study. J. AOAC Int. 2012, 95, 795–812. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M.; Leighfield, T.A.; Doucette, G.J.; Bean, L.; Niedzwiadek, B.; Rawn, D.F.K. Single-laboratory validation of the microplate receptor binding assay for paralytic shellfish toxins in shellfish. J. AOAC Int. 2009, 92, 1705–1713. [Google Scholar] [PubMed]
- Costa, P.R.; Botelho, M.J.; Lefebvre, K.A. Characterization of paralytic shellfish toxins in seawater and sardines (Sardina pilchardus) during blooms of Gymnodinium catenatum. Hydrobiologia 2010, 655, 89–97. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Bill, B.D.; Erikson, A.; Baugh, K.A.; O’Rourke, L.; Costa, P.R.; Nance, S.; Trainer, V.L. Characterization of intracellular and extracellular saxitoxin levels in both field and cultured Alexandrium spp. samples from Sequim Bay, Washington. Mar. Drugs 2008, 6, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Quilliam, M.A.; Hess, P.; Dell’Aversano, C. Recent developments in the analysis of phycotoxins by liquid chromatography-mass spectrometry. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Century; DeKoe, W.J., Samson, R.A., van Egmond, H.P., Gilbert, J., Sabino, M., Eds.; Publisher: Wageningen, The Netherlands, 2011; pp. 383–391. [Google Scholar]
- Dell’Aversano, C.; Hess, P.; Quilliam, M.A. Hydrophillic interaction liquid chromatography-mass spectrometry for the analysis of paralytic shellfish poisoning (PSP) toxins. J. Chromatogr. A 2005, 1081, 190–201. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, P.R.; Robertson, A.; Quilliam, M.A. Toxin Profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese Coast, as Determined by Liquid Chromatography Tandem Mass Spectrometry. Mar. Drugs 2015, 13, 2046-2062. https://doi.org/10.3390/md13042046
Costa PR, Robertson A, Quilliam MA. Toxin Profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese Coast, as Determined by Liquid Chromatography Tandem Mass Spectrometry. Marine Drugs. 2015; 13(4):2046-2062. https://doi.org/10.3390/md13042046
Chicago/Turabian StyleCosta, Pedro R., Alison Robertson, and Michael A. Quilliam. 2015. "Toxin Profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese Coast, as Determined by Liquid Chromatography Tandem Mass Spectrometry" Marine Drugs 13, no. 4: 2046-2062. https://doi.org/10.3390/md13042046
APA StyleCosta, P. R., Robertson, A., & Quilliam, M. A. (2015). Toxin Profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese Coast, as Determined by Liquid Chromatography Tandem Mass Spectrometry. Marine Drugs, 13(4), 2046-2062. https://doi.org/10.3390/md13042046