
New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel



Abstract
:1. Introduction
2. Results and Discussion
2.1. Structural Elucidation of Aeruginosin KB676


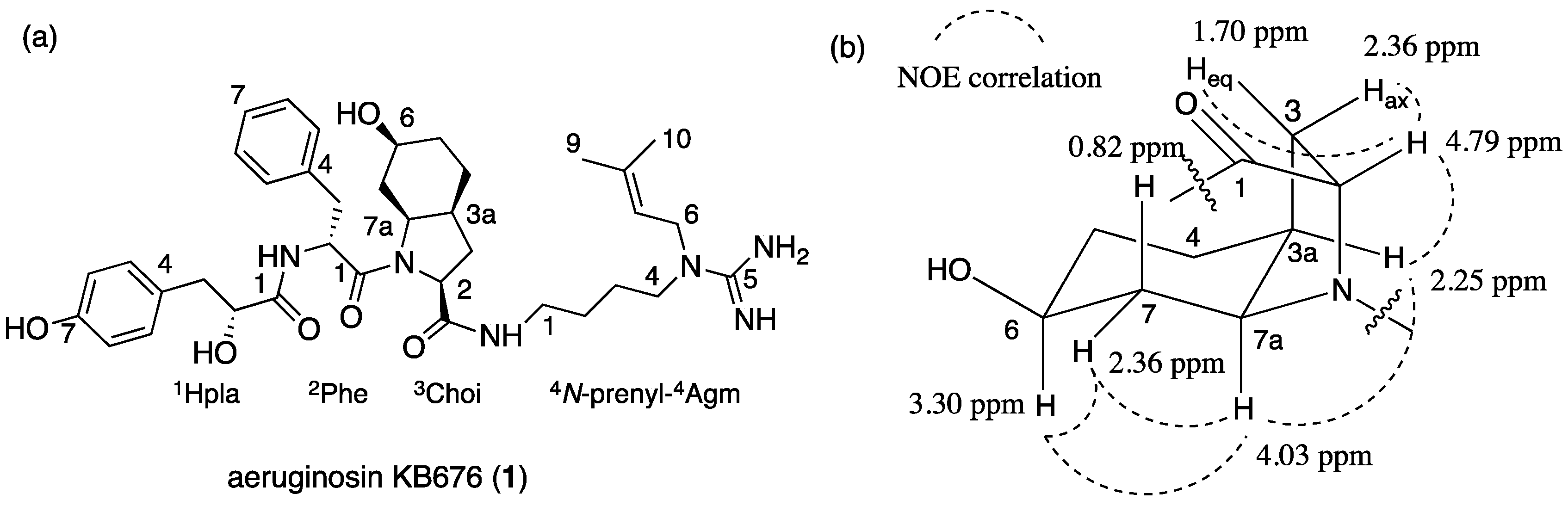{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC mult. b | δH mult. b J in Hz | HMBC Correlations c | NOE Correlations d |
|---|---|---|---|---|
| 1Hpla | ||||
| 1 | 173.5 qC | - | ||
| 2 | 72.4 CH | 3.87 m | Hpla-3,3′,5,5′; Phe-5,5′,6,6′,NH | |
| 3 | 40.0 CH2 | 2.60 dd (14.0,3.5) | Hpla-1,2,4,5,5′ | Hpla-2,3′,5,5′ |
| 2.27 dd (14.0,8.5) | Hpla-1,2,4,5,5′ | Hpla-2,3,5,5′ | ||
| 4 | 128.6 qC | - | ||
| 5,5′ | 130.3 CH | 6.85 d (8.3) | Hpla-3,6,6′,7 | Hpla-2,3,3′,6,6′ |
| 6,6′ | 115.0 CH | 6.59 d (8.3) | Hpla-4,7 | Hpla-5,5',7-OH |
| 7 | 155.8 qC | - | ||
| 2-OH | - | 5.31 brs | ||
| 7-OH | - | 9.09 s | Hpla-7 | Hpla-6,6′ |
| 2Phe | ||||
| 1 | 170.0 qC | - | ||
| 2 | 51.4 CH | 4.26 q (6.0) | Phe-1,3 | Phe-3,5,5′,NH; Choi-2,3 |
| 3 | 36.6 CH2 | 2.80 m | Phe-1,4 | Phe-2,5,5′,NH |
| 4 | 138.2 qC | - | ||
| 5,5′ | 129.3 CH | 7.10 m | Phe-3,7 | Phe-2,3,6,6′; Hpla-2 |
| 6,6′ | 128.2 CH | 7.22 m | Phe-4 | Phe-3,5,5′,7; Hpla-2 |
| 7 | 126.4 CH | 7.16 m | Phe-5 | Phe-6,6′ |
| NH | - | 7.76 d (8.0) | Hpla-1; Phe-2 | Phe-2,3; Hpla-2 |
| 3Choi | ||||
| 1 | 172.4 qC | - | ||
| 2 | 59.7 CH | 4.79 d (9) | Choi-3 | Choi-3,3′,3a; Phe-2; Agm-1-NH |
| 3 (ax) (eq) | 32.8 CH2 | 2.36 m | Choi-7a | Choi-2,3′,3a; Phe-2 |
| 1.70 m | Choi-2,3,3a; Phe-2 | |||
| 3a | 32.2 CH | 2.25 m | Choi-2,3,3′,4,7a | |
| 4 | 22.8 CH2 | 1.59 m, 2H | Choi-3a,5,6,7a | |
| 5 (eq) (ax) | 29.9 CH2 | 1.57 m | Choi-4,5′,6 | |
| 1.22 m | Choi-4,5,7ax | |||
| 6 | 67.0 CH | 3.30 m | Choi-4,5,7eq,7a | |
| 6-OH | - | 4.55 brs | ||
| 7 (eq) (ax) | 36.2 CH2 | 2.36 m | Choi-3a,6,7a | Choi-4,6,7′,7a; Phe-2 |
| 0.82 m | Choi-5ax,7eq | |||
| 7a | 56.8 CH | 4.03 m | Choi-3a,5,6,7eq | |
| 4N-prenyl-4Agm | ||||
| 1 | 38.6 CH2 | 3.16 m | Choi-1 | Agm-1′,2,3,6,7,1-NH |
| 3.05 m | Choi-1, Agm-2,3 | Agm-1,2,3,1-NH | ||
| 2 | 26.4 CH2 | 1.40 m | Agm-1,3 | Agm-1,1′,3,4,6,1-NH |
| 3 | 24.7 CH2 | 1.48 m | Agm-1,2 | Agm-1,1′,2,4,6,7,1-NH |
| 4 | 47.3 CH2 | 3.16 m | Agm-2,3,5,6 | Agm-2,3,6,7,1-NH |
| 5 | 155.8 qC | - | ||
| 6 | 45.9 CH2 | 3.87 brd (6.0) | Agm-4,5,7,8 | Agm-1,2,3,4,7,9,10 |
| 8 | 137.1 qC | - | ||
| 9 | 25.7 CH3 | 1.70 brs | Agm-7,8 | Agm-6,7 |
| 10 | 18.0 CH3 | 1.64 brs | Agm-7,8 | Agm-6,1-NH |
| 1-NH | - | 8.23 t (5.5) | Choi-1 | Agm-1,1′,2,3,4,10; Choi-2 |
| 5-NH,NH2 | - | 7.25 brm |
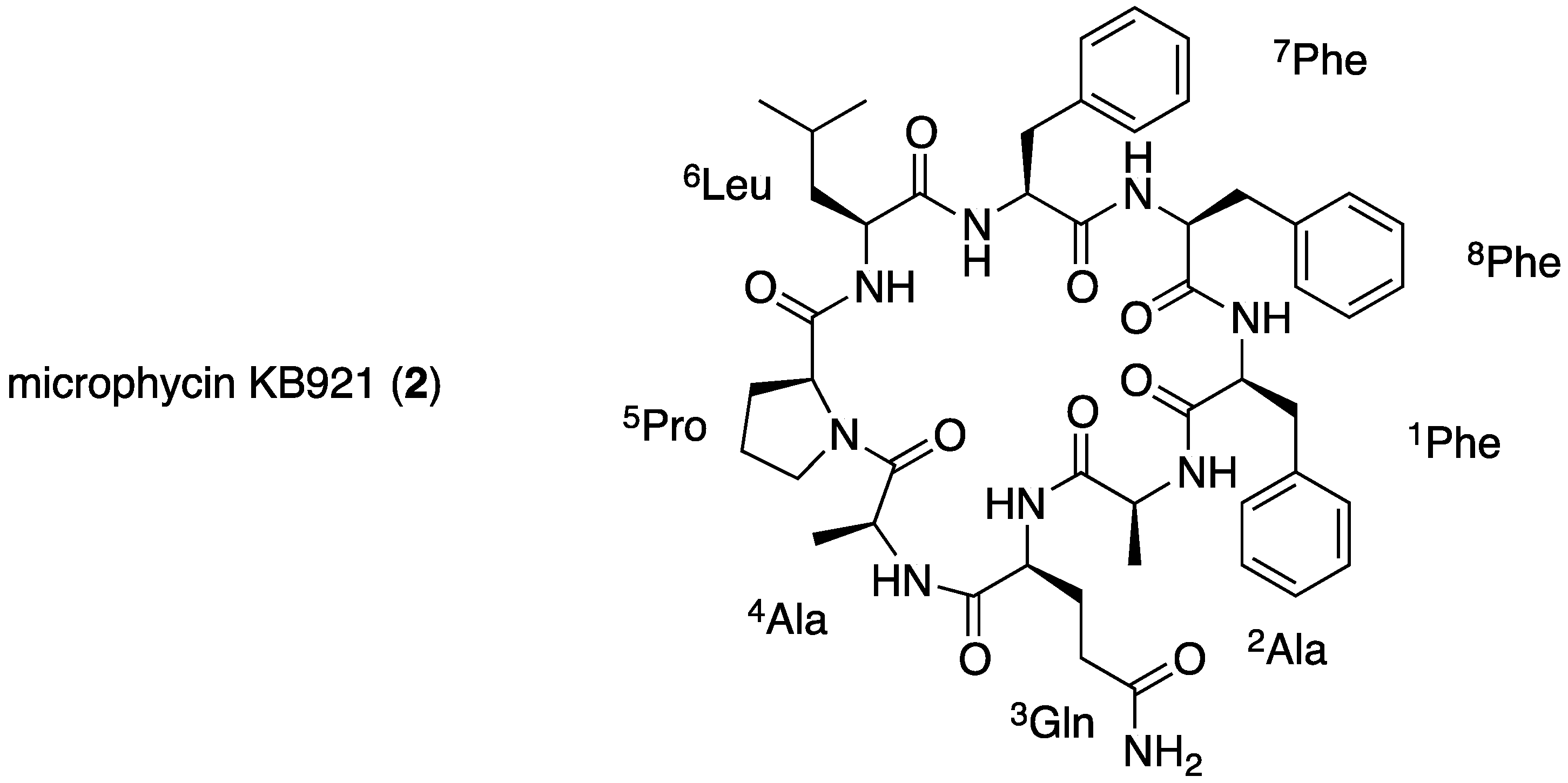
2.2. Structural Elucidation of Microphycin KB921

| Compound | |||||||
|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | |||||
| Position | δC a | δH b | Position | δC a | δH b | δC c | δH d |
| 1Phe | 1Ile | ||||||
| 1 | 171.1 qC | 1 | 170.7 qC | - | 170.7 qC | - | |
| 2 | 52.9 CH | 4.91 brdd | 2 | 58.1 CH | 4.19 m | 58.1 CH | 4.15 m |
| 3 | 39.0 CH2 | 3.38 m | 3 | 36.1 CH | 1.97 m | 35.9 CH | 1.98 m |
| 2.62 t | |||||||
| 4 | 137.7 qC | - | 4 | 24.2 CH2 | 1.31 m | 24.2 CH2 | 1.28 m |
| 0.98 m | 0.93 m | ||||||
| 5,5′ | 129.7 CH | 7.31 m | 5 | 11.6 CH3 | 0.75 t | 11.4 CH3 | 0.75 t |
| 128.2 CH | 7.27 m | 6 | 16.1 CH3 | 0.77 d | 16.1 CH3 | 0.76 d | |
| 7 | 126.5 CH | 7.20 m | NH | - | 8.10 d | - | 8.17 d |
| NH | 7.27 m | 2NMeHty | |||||
| 2Ala | 1 | 169.5 qC | - | 169.4 qC | - | ||
| 1 | 173.6 qC | - | 2 | 59.5 CH | 4.66 t | 59.5 CH | 4.58 t |
| 2 | 52.6 CH | 3.84 dq | 3 | 30.9 CH2 | 2.03 m | 30.8 CH2 | 1.98 m |
| 1.70 m | 1.70 m | ||||||
| 3 | 17.0 CH3 | 1.31 d | 4 | 31.6 CH2 | 2.32 m | 31.4 CH2 | 2.31 m |
| 2.23 m | 2.27 m | ||||||
| NH | 8.96 m | 5 | 131.6 qC | - | 131.6 qC | - | |
| 3Gln | 6,6′ | 129.1 CH | 6.95 d | 129.0 CH | 6.95 d | ||
| 1 | 174.0 qC | - | 7,7′ | 115.4 CH | 6.66 d | 115.3 CH | 6.66 d |
| 2 | 53.1 CH | 4.08 dt | 8 | 155.7 qC | - | 155.7 qC | - |
| 3 | 29.7 CH2 | 2.12 m | NMe | 28.8 CH3 | 2.58 s | 28.6 CH3 | 2.57 s |
| 1.84 m | |||||||
| 4 | 31.8 CH2 | 2.14 m | 3Hph | ||||
| 5 | 174.3 qC | - | 1 | 172.4 qC | - | 173.2 qC | - |
| NH | - | 8.18 d | 2 | 48.6 CH | 4.71 m | 48.1 CH | 4.69 m |
| NH2 | - | 6.80 s | 3 | 33.2 CH2 | 2.00 m | 33.2 CH2 | 2.02 m |
| 7.33 s | 1.78 m | 1.75 m | |||||
| 4Ala | 4 | 31.7 CH2 | 2.81 m | 31.6 CH2 | 2.81 m | ||
| 2.66 m | 2.66 m | ||||||
| 1 | 171.9 qC | - | 5 | 141.3 qC | - | 141.2 qC | |
| 2 | 47.2 CH | 4.58 dq | 6,6’ | 128.6 CH | 7.25 d | 128.6 CH | 7.25 m |
| 3 | 16.4 CH3 | 1.21 d | 7,7’ | 128.6 CH | 7.23 t | 128.5 CH | 7.23 m |
| NH | - | 7.07 m | 8 | 126.4 CH | 7.18 t | 126.3 CH | 7.18 m |
| 5Pro | NH | - | 8.93 d | - | 8.98 d | ||
| 1 | 174.8 qC | - | 4Ile/Val | ||||
| 2 | 62.8 CH | 4.31 t | 1 | 173.1 qC | - | 172.9 qC | - |
| 3 | 29.2 CH2 | 2.15 dd | 2 | 55.6 CH | 4.21 t | 58.1 CH | 3.94 t |
| 1.67 m | |||||||
| 4 | 24.6 CH2 | 1.85 m | 3 | 36.4 CH | 1.80 m | 30.1 CH | 1.88 m |
| 1.47 m | |||||||
| 5 | 47.3 CH2 | 3.54 m | 4 | 25.7 CH2 | 1.34 m | 19.1 CH3 | 0.88 d |
| 3.38 m | 1.12 m | ||||||
| 6Leu | 5 | 11.8 CH3 | 0.83 t | 18.7 CH3 | 0.86 d | ||
| 1 | 171.0 qC | - | 6 | 14.5 CH3 | 0.78 d | - | - |
| 2 | 53.8 CH | 3.67 m | NH | - | 6.70 d | - | 6.78 d |
| 3 | 38.6 CH2 | 1.30 m | 5Lys | ||||
| 1.11 m | |||||||
| 4 | 24.3 CH | 1.33 m | 1 | 172.4 qC | - | 172.4 qC | - |
| 5 | 22.6 CH3 | 0.79 d | 2 | 55.0 CH | 3.88 ddd | 54.9 CH | 3.85 ddd |
| 6 | 21.6 CH3 | 0.67 d | 3 | 30.9 CH2 | 1.61 m | 31.1 CH2 | 1.61 m |
| NH | - | 8.16 d | 4 | 20.6 CH2 | 1.30 m | 20.6 CH2 | 1.39 m |
| 1.24 m | 1.32 m | ||||||
| 7Phe | 5 | 28.3 CH2 | 1.38 m | 28.2 CH2 | 1.40 m | ||
| 1 | 170.4 qC | - | 6 | 38.2 CH2 | 3.48 m | 38.3 CH2 | 3.43 m |
| 2.78 m | |||||||
| 2 | 53.2 CH | 4.32 brdd | α-NH | - | 6.51 d | - | 2.81 m |
| 3 | 36.9 CH2 | 3.47 m | ε-NH | - | 7.13 m | - | 7.07 m |
| 2.73 t | |||||||
| 4 | 138.6 qC | - | 6Arg/Tyr | ||||
| 5,5′ | 129.2 CH | 7.14 m | 1 | 174.3 qC | - | 173.9 qC | - |
| 6,6′ | 128.2 CH | 7.22 m | 2 | 52.1 CH | 4.09 dt | 54.2 CH | 4.26 m |
| 7 | 126.4 CH | 7.17 m | 3 | 29.6 CH2 | 1.69 m | 36.9 CH2 | 2.86 m |
| 1.52 m | 2.75 m | ||||||
| NH | - | 7.78 d | 4 | 25.2 CH2 | 1.46 m | 127.4 qC | - |
| 8Phe | 5 | 40.5 CH2 | 3.09 m | 130.3 CH | 6.95 d | ||
| 1 | 169.5 qC | 6 | 156.9 qC | - | 115.2 CH | 6.64 d | |
| 2 | 56.9 CH | 3.65 m | 7 | - | - | 156.0 qC | - |
| 3 | 32.6 CH2 | 3.22 m | α-NH | - | 6.42 d | - | 6.19 d |
| 4 | 139.5 qC | - | δ -NH/OH | - | 7.50 brm | - | 9.21 s |
| 5,5′ | 129.0 CH | 7.07 m | NH/NH2 | - | 6.65 brm | ||
| 7.30 brm | |||||||
| 6,6′ | 128.4 CH | 7.20 m | CO | 157.5 qC | - | 157.2 qC | |
| 7 | 126.2 CH | 7.18 m | |||||
| NH | - | 7.57 d | |||||

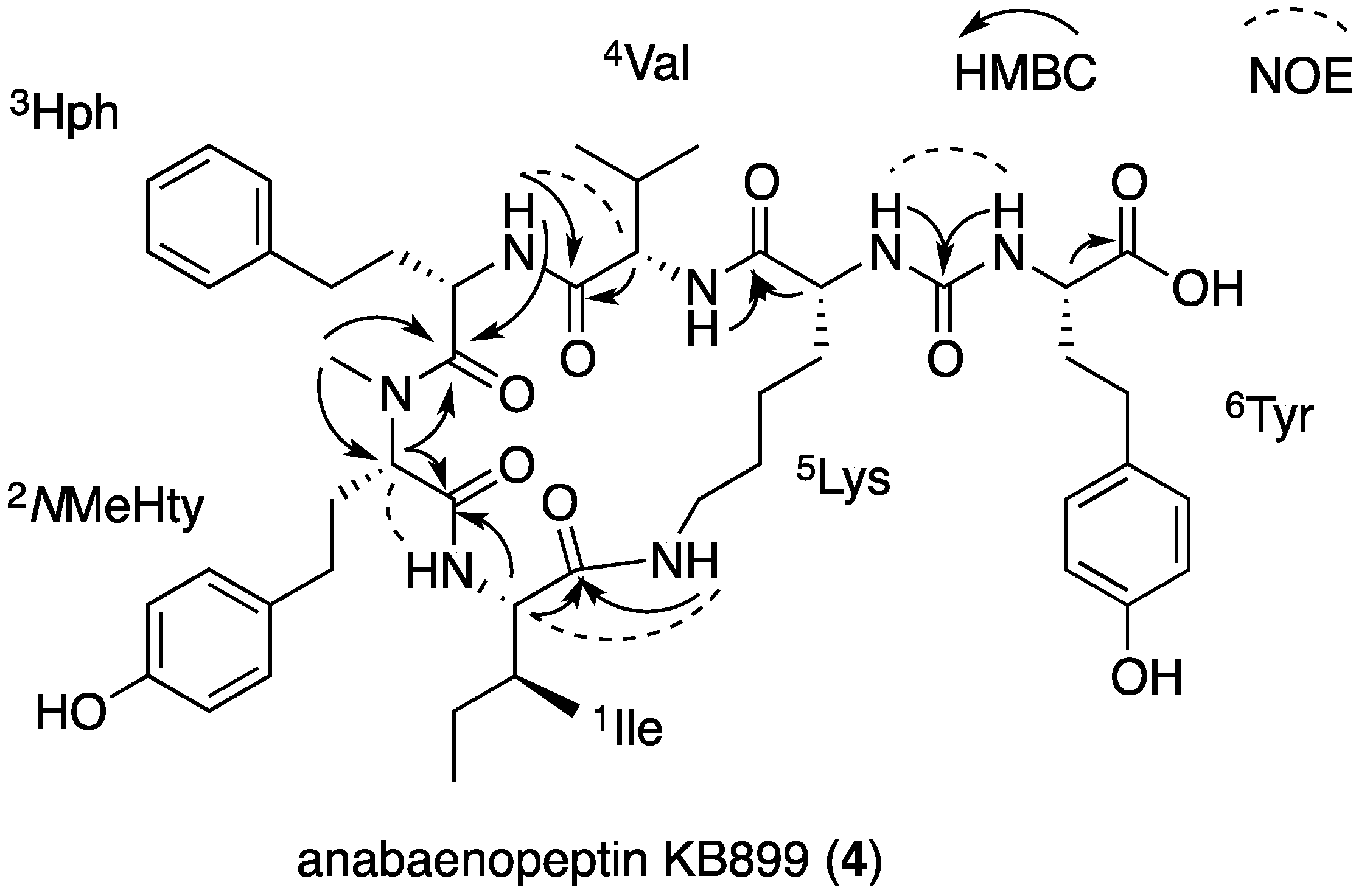
2.3. Structural Elucidation of Anabaenopeptins KB906 and KB899


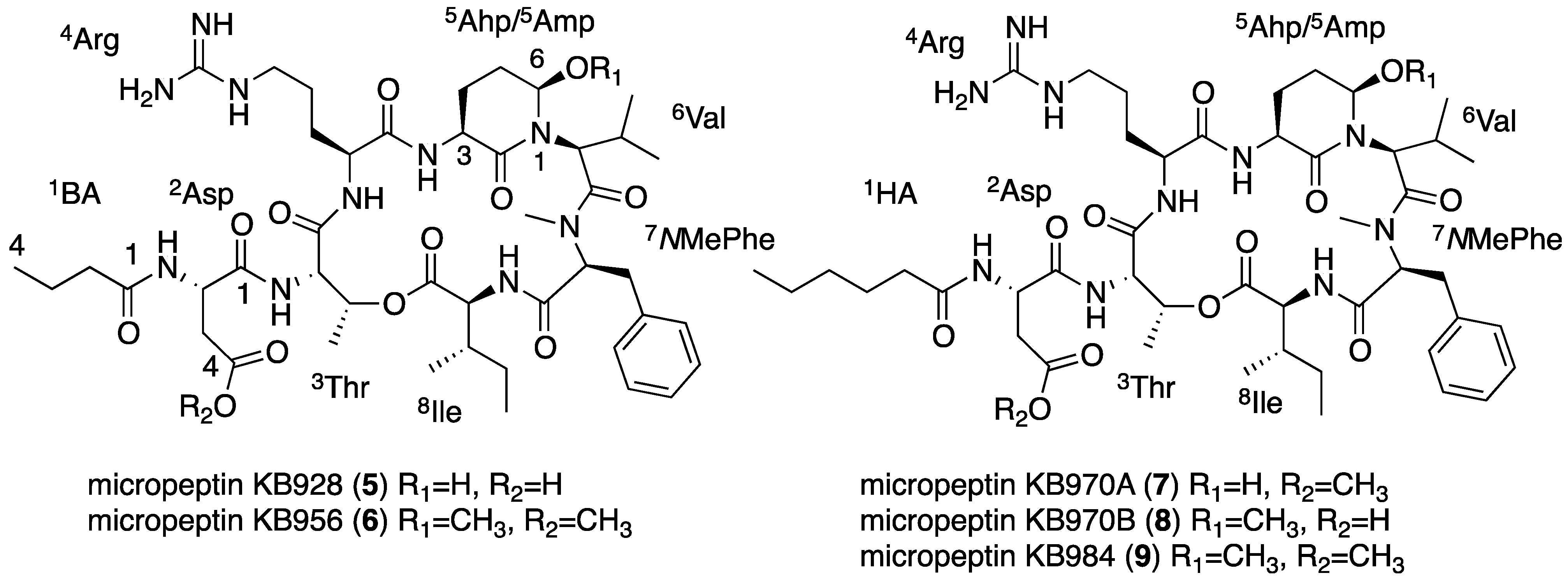
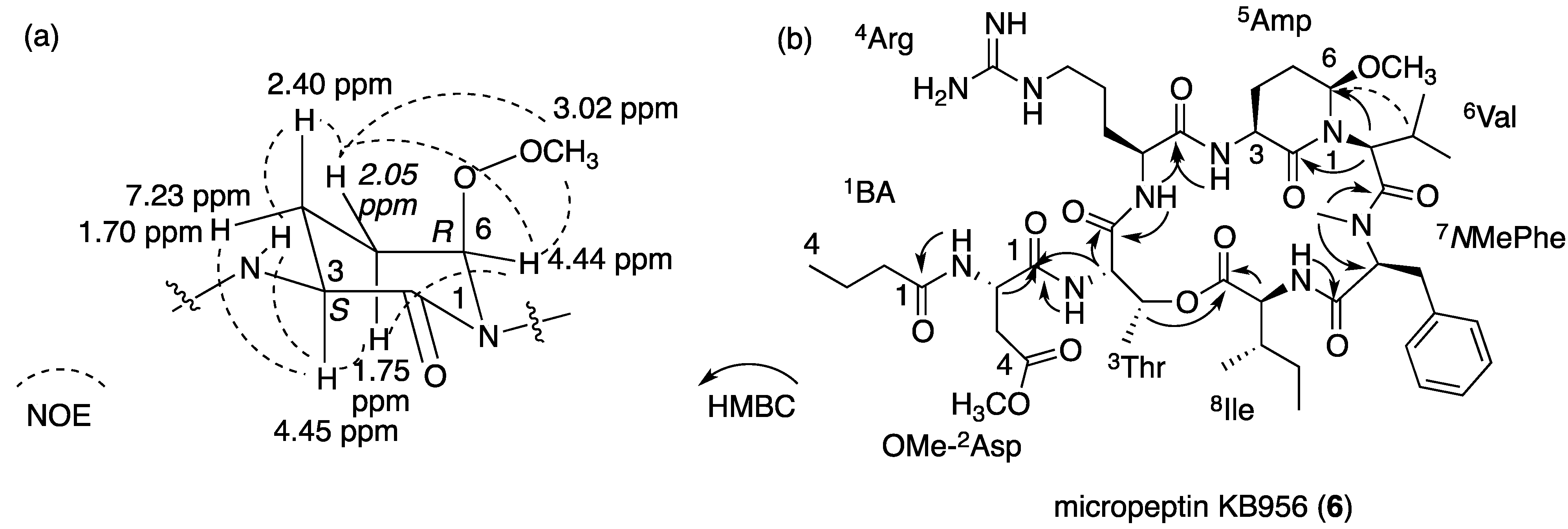
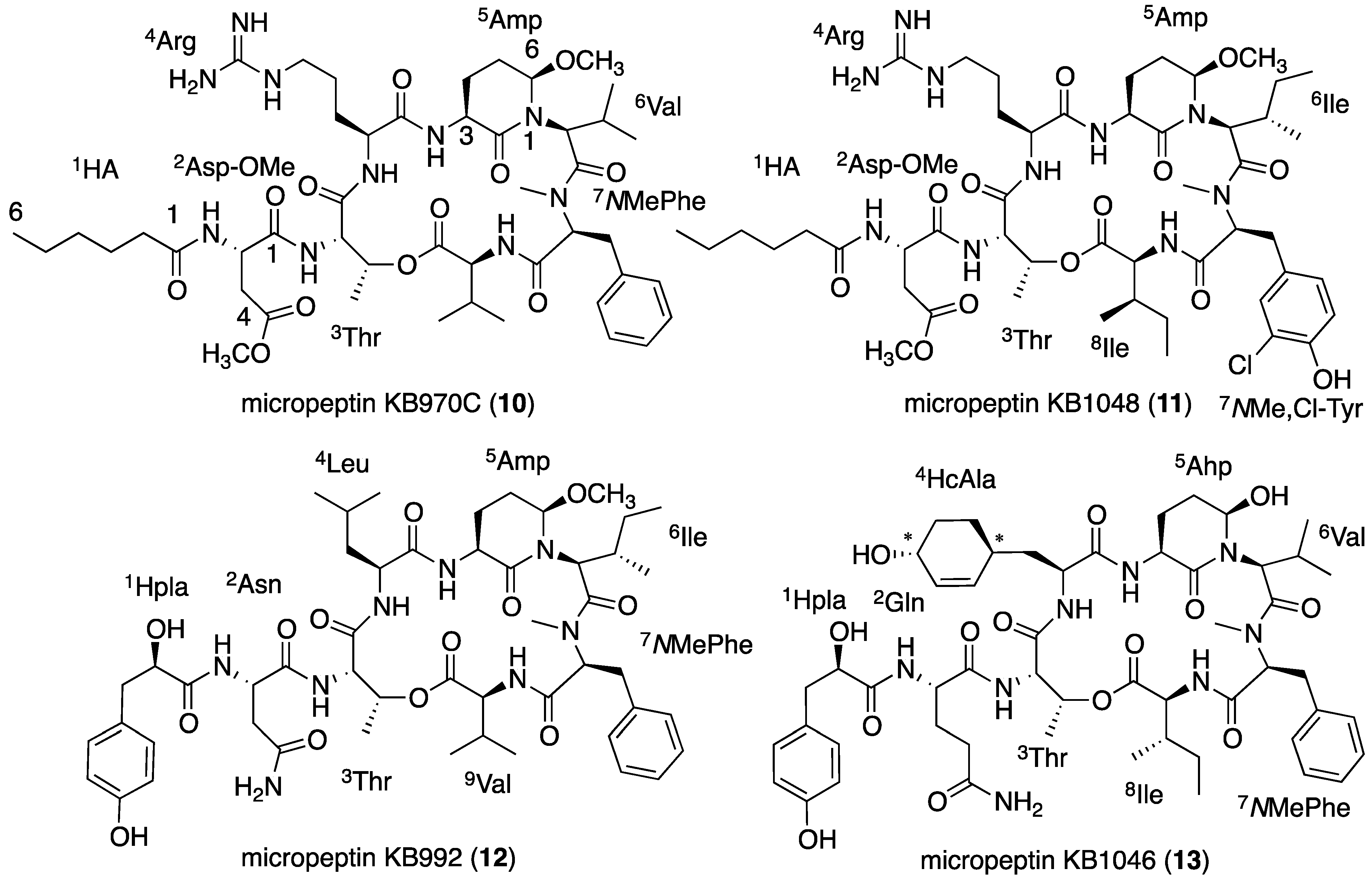
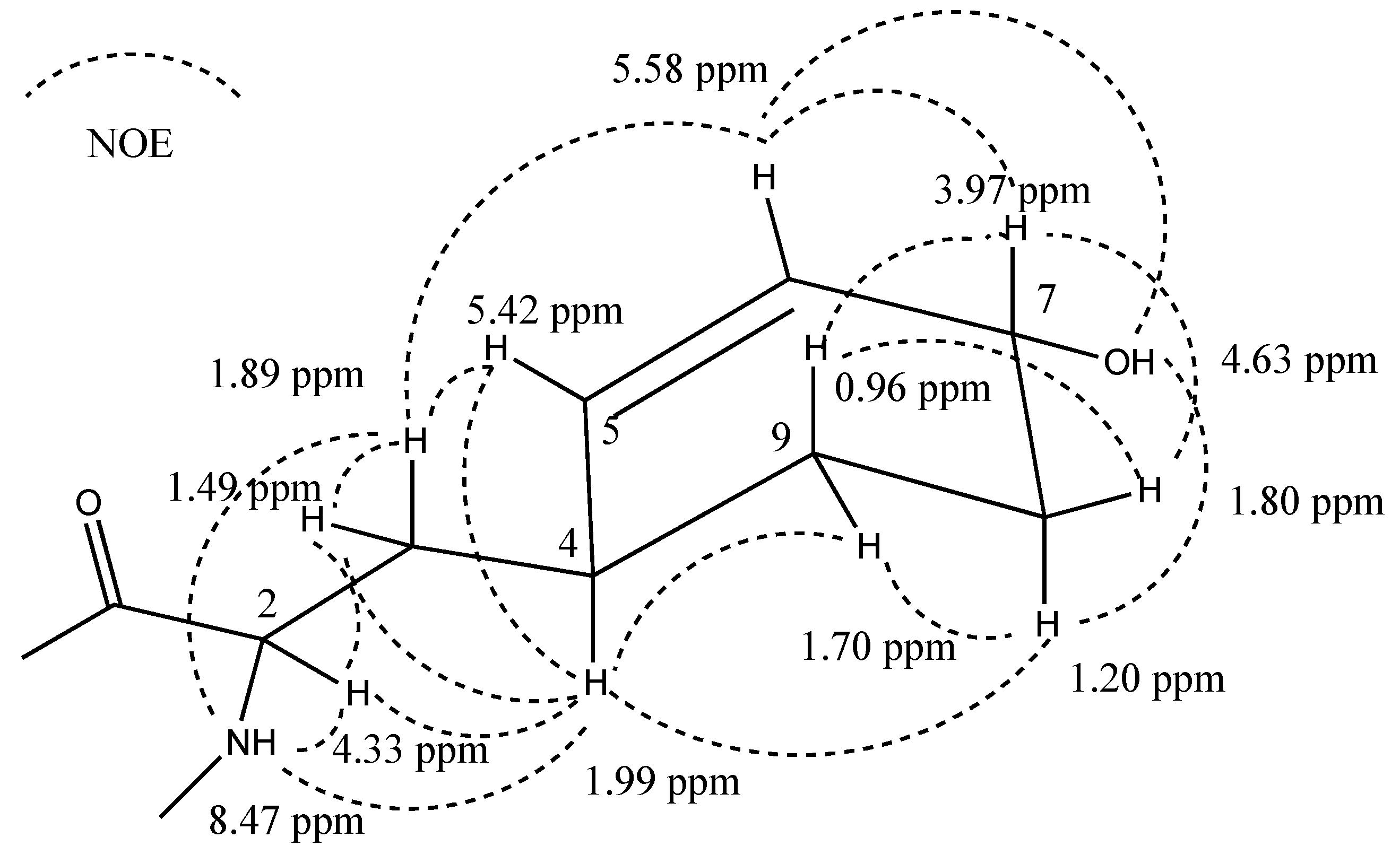
2.4. Structural Elucidation of the New Micropeptins

| Compound | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 6 | 7 | 8 | 9 | ||||||
| Position | δC a | δH b | δC a | δH b | δC a | δH b | δC c | δH d | δC a | δH b |
| 1BA/1HA e | ||||||||||
| 1 | 172.0 | - | 172.8 | - | 173.0 | - | 173.0 | - | 173.1 | - |
| 2 | 37.4 | 2.11 t | 37.3 | 2.11 t | 35.4 | 2.13 t | 35.6 | 2.13 t | 35.7 | 2.13 t |
| 3 | 18.9 | 1.53 qi | 18.9 | 1.55 qi | 25.2 | 1.52 qi | 25.1 | 1.53 qi | 25.2 | 1.51 qi |
| 4 | 13.8 | 0.88 t | 13.8 | 0.88 t | 31.1 | 1.22 m | 31.1 | 1.25 m | 31.1 | 1.23 m |
| 5 | - | - | - | - | 22.1 | 1.27 m | 22.0 | 1.27 m | 22.1 | 1.27 m |
| 6 | - | - | - | - | 14.2 | 0.84 t | 14.1 | 0.85 t | 14.2 | 0.84 t |
| 2Asp | ||||||||||
| 1 | 172.6 | - | 171.4 | - | 171.6 | - | 171.7 | - | 171.6 | - |
| 2 | 49.8 | 4.55 m | 49.5 | 4.86 q | 49.7 | 4.71 ddd | 49.7 | 4.63 m | 49.6 | 4.71 q |
| 3 | 39.0 | 2.58 m | 35.5 | 2.80 m | 35.7 | 2.82 dd | 35.4 | 2.80 m | 35.4 | 2.81 m |
| 2.20 m | 2.60 dd | 2.58 dd | 2.58 m | 2.58 dd | ||||||
| 4 | 172.0 | - | 170.9 | - | 171.0 | - | 172.0 | - | 171.0 | - |
| NH | - | 8.09 d | - | 8.33 d | - | 8.27 d | - | 8.28 d | - | 8.29 d |
| OMe | - | - | 51.7 | 3.56 s | 51.8 | 3.56 s | - | - | 51.8 | 3.56 s |
| 3Thr | ||||||||||
| 1 | 169.2 | - | 169.0 | - | 169.3 | - | 169.1 | - | 169.2 | - |
| 2 | 54.8 | 4.62 d | 54.9 | 4.60 d | 55.1 | 4.57 d | 54.9 | 4.60 d | 55.1 | 4.60 d |
| 3 | 72.2 | 5.45 q | 72.2 | 5.49 q | 72.2 | 5.47 q | 72.3 | 5.48 q | 72.3 | 5.49 q |
| 4 | 17.8 | 1.17 d | 18.2 | 1.18 d | 18.2 | 1.19 d | 17.8 | 1.17 d | 18.0 | 1.18 d |
| NH | - | 7.44 brd | - | 7.65 d | - | 7.46 d | - | 7.63 d | - | 7.68 d |
| 4Arg | ||||||||||
| 1 | 170.3 | 170.3 | - | 170.4 | - | 170.4 | - | 170.5 | - | |
| 2 | 51.8 | 4.28 m | 52.2 | 4.28 m | 52.5 | 4.26 m | 52.2 | 4.28 m | 52.4 | 4.27 m |
| 3 | 27.5 | 2.02 m | 27.7 | 2.02 m | 27.6 | 2.02 m | 27.8 | 2.02 m | 27.9 | 2.02 m |
| 1.45 m | 1.45 m | 1.45 m | 1.45 m | 1.45 m | ||||||
| 4 | 25.2 | 1.45 m | 25.4 | 1.45 m | 25.8 | 1.45 m | 25.4 | 1.45 m | 25.5 | 1.45 m |
| 5 | 40.1 | 3.05 m | 40.8 | 3.08 m | 41.0 | 3.09 m | 40.2 | 3.09 m | 40.7 | 3.09 m |
| NH | - | 8.57 d | - | 8.57 d | - | 8.50 d | - | 8.54 d | - | 8.56 d |
| 5-NH | - | 7.49 brt | - | 7.53 t | - | 7.49 t | - | 7.47 m | - | 7.56 t |
| 6 | 156.9 | - | 156.8 | - | 157.0 | - | 156.9 | - | 157.0 | - |
| 6-NH,NH2 | - | 7.30 brm | - | 7.30 brm | - | 7.30 brm | 7.30 brm | - | 7.30 brm | |
| 6.60 brm | 6.70 brm | 6.80 brm | 6.77 brm | 6.85 brm | ||||||
| 5Ahp/5Amp | ||||||||||
| 2 | 169.5 | - | 169.2 | - | 169.5 | - | 169.2 | - | 169.3 | - |
| 3 | 49.0 | 4.44 m | 49.3 | 4.45 m | 49.3 | 4.44 m | 49.4 | 4.47 m | 49.4 | 4.47 m |
| 4 | 21.7 | 2.55 m | 21.7 | 2.40 brq | 21.9 | 2.53 m | 21.7 | 2.40 brq | 21.9 | 2.40 brq |
| 1.73 m | 1.70 m | 1.73 m | 1.75 m | 1.70 m | ||||||
| 5 | 29.2 | 1.70 m | 23.8 | 2.05 m | 30.0 | 1.72 m | 23.8 | 2.05 m | 23.9 | 2.05 m |
| 1.75 m | 1.70 m | 1.75 m | ||||||||
| 6 | 74.2 | 4.91 brs | 83.3 | 4.44 brs | 74.3 | 4.91 brs | 83.3 | 4.44 brs | 83.4 | 4.44 brs |
| NH | - | 7.30 d | - | 7.23 m | - | 7.36 d | 7.24 m | - | 7.25 m | |
| OH/OMe | 6.11 d | 55.5 | 3.02 s | - | 6.17 d | 55.6 | 3.02 s | 55.6 | 3.02 s | |
| 6Val | ||||||||||
| 1 | 169.8 | - | 169.7 | - | 170.0 | - | 169.8 | - | 169.9 | - |
| 2 | 55.7 | 4.31 d | 55.7 | 4.35 d | 56.1 | 4.31 d | 55.8 | 4.35 d | 55.9 | 4.36 d |
| 3 | 27.5 | 1.90 m | 27.2 | 1.95 m | 27.8 | 1.90 m | 27.3 | 1.95 m | 27.4 | 1.95 m |
| 4 | 18.2 | 0.46 d | 18.9 | 0.46 d | 18.4 | 0.46 d | 18.3 | 0.46 d | 18.4 | 0.46 d |
| 5 | 18.1 | −0.20 d | 17.8 | −0.24 d | 18.4 | −0.19 d | 17.8 | −0.23 d | 18.0 | −0.23 d |
| 7NMePhe | ||||||||||
| 1 | 169.2 | - | 169.1 | - | 169.5 | - | 169.2 | - | 169.3 | - |
| 2 | 60.6 | 5.06 brd | 60.8 | 5.09 brd | 60.8 | 5.06 brd | 60.9 | 5.10 brd | 61.0 | 5.10 brd |
| 3 | 34.3 | 3.28 m | 34.2 | 3.25 m | 34.4 | 3.26 m | 34.3 | 3.30 m | 34.4 | 3.29 brd |
| 2.80 m | 2.80 m | 2.80 dd | 2.81 dd | 2.80 m | ||||||
| 4 | 137.8 | 137.7 | - | 137.9 | - | 137.8 | - | 137.8 | - | |
| 5,5′ | 129.8 | 7.23 d | 129.7 | 7.22 m | 129.9 | 7.22 d | 129.8 | 7.22 m | 129.9 | 7.22 m |
| 6,6′ | 128.7 | 7.26 t | 128.8 | 7.27 m | 128.9 | 7.27 d | 128.9 | 7.27 m | 129.0 | 7.26 m |
| 7 | 126.8 | 7.19 t | 126.9 | 7.19 m | 127.0 | 7.19 d | 127.0 | 7.19 m | 127.0 | 7.19 m |
| NMe | 30.3 | 2.73 s | 30.2 | 2.73 s | 30.4 | 2.73 s | 30.3 | 2.73 s | 30.4 | 2.73 s |
| 8Ile | ||||||||||
| 1 | 172.7 | - | 172.6 | - | 172.9 | - | 172.6 | - | 172.7 | - |
| 2 | 55.9 | 4.69 dd | 56.6 | 4.64 dd | 56.0 | 4.63 dd | 55.6 | 4.64 m | 55.8 | 4.65 dd |
| 3 | 37.4 | 1.78 m | 37.7 | 1.74 m | 37.3 | 1.77 m | 37.8 | 1.74 m | 37.8 | 1.74 m |
| 4 | 24.7 | 1.23 m | 24.8 | 1.31 m | 24.8 | 1.23 m | 24.9 | 1.30 m | 25.0 | 1.30 m |
| 1.04 m | 1.10 m | 1.02 m | 1.10 m | 1.10 m | ||||||
| 5 | 11.3 | 0.81 t | 11.1 | 0.83 t | 11.4 | 0.80 t | 11.2 | 0.85 t | 11.2 | 0.84 t |
| 6 | 16.0 | 0.83 d | 16.0 | 0.85 d | 16.2 | 0.84 t | 16.0 | 0.87 d | 16.1 | 0.87 d |
| NH | - | 7.68 d | - | 6.96 d | - | 7.76 d | - | 6.95 d | - | 6.96 d |


| Compound | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 10 | 11 | 12 | 13 | ||||||
| Position | δC a | δH b | δC a | δH b | Position | δC a | δH b | δC c | δH d |
| 1HA | 1Hpla | ||||||||
| 1 | 172.9 | - | 173.0 | - | 1 | 174.2 | - | 173.6 | - |
| 2 | 35.6 | 2.13 t | 35.5 | 2.17 t | 2 | 73.0 | 3.99 ddd | 72.7 | 4.01 ddd |
| 3 | 25.1 | 1.52 qi | 25.1 | 1.51 qi | 3 | 41.0 | 2.93 dd | 39.8 | 2.87 dd |
| 2.54 m | 2.55 m | ||||||||
| 4 | 31.0 | 1.25 m | 31.0 | 1.22 m | 4 | 129.1 | 128.8 | - | |
| 5 | 22.1 | 1.27 m | 22.1 | 1.24 m | 5,5′ | 130.5 | 7.02 d | 130.4 | 7.00 d |
| 6 | 14.1 | 0.84 t | 14.8 | 0.85 t | 6,6′ | 115.0 | 6.64 d | 115.0 | 6.62 d |
| 2Asp | 7 | 155.8 | - | 155.8 | - | ||||
| 1 | 171.5 | - | 171.5 | - | 2-OH | - | 5.60 d | - | 5.52 d |
| 2 | 49.5 | 4.70 q | 49.5 | 4.70 q | 7-OH | - | 9.11 s | - | 9.06 s |
| 3 | 35.3 | 2.80 m | 35.5 | 2.81 dd | 2Asn/2Gln | ||||
| 2.58 dd | 2.59 dd | ||||||||
| 4 | 170.8 | - | 170.9 | - | 1 | 171.8 | - | 172.4 | - |
| NH | - | 8.27 d | - | 8.28 d | 2 | 49.3 | 4.70 q | 51.4 | 4.51 dt |
| OMe | 51.7 | 3.56 s | 51.7 | 3.57 s | 3 | 36.9 | 2.54 m | 28.9 | 1.82 m |
| 1.75 m | |||||||||
| 3Thr | 4 | 172.2 | - | 31.6 | 2.10 t | ||||
| 1 | 169.1 | - | 169.1 | - | 5 | 174.0 | - | ||
| 2 | 55.0 | 4.58 d | 55.0 | 4.59 d | NH | - | 8.23 d | - | 7.78 d |
| NH2 | - | 7.39 s6.89 s | - | 7.18 s6.70 s | |||||
| 3 | 72.2 | 5.50 q | 72.3 | 5.47 q | 3Thr | ||||
| 4 | 17.9 | 1.18 d | 17.8 | 1.17 d | 1 | 169.2 | - | 169.6 | - |
| NH | - | 7.68 d | - | 7.71 d | 2 | 55.1 | 4.60 d | 55.3 | 4.62 d |
| 4Arg | 3 | 72.2 | 5.50 q | 71.8 | 5.52 q | ||||
| 1 | 170.4 | - | 170.3 | - | 4 | 17.9 | 1.22 d | 17.9 | 1.23 d |
| 2 | 52.2 | 4.27 m | 52.1 | 4.29 m | NH | - | 7.74 d | - | 8.20 d |
| 3 | 27.8 | 2.02 m | 27.7 | 2.00 m | 4Leu/4HcAla | ||||
| 1.45 m | 1.45 m | ||||||||
| 4 | 25.4 | 1.45 m | 25.4 | 1.45 m | 12 | 171.0 | - | 170.7 | - |
| 5 | 40.2 | 3.09 m | 40.1 | 3.08 m | 2 | 51.0 | 4.28 ddd | 50.0 | 4.33 m |
| NH | - | 8.56 d | - | 8.56 d | 3 | 39.0 | 1.80 m | 36.6 | 1.89 m |
| 1.39 m | 1.49 m | ||||||||
| 5-NH | - | 7.56 t | - | 7.50 t | 4 | 24.4 | 1.50 m | 32.0 | 1.99 m |
| 6 | 156.9 | - | 156.9 | - | 5 | 23.9 | 0.87 d | 132.2 | 5.42 brd |
| 6-NH,NH2 | - | 7.30 brm | - | 7.30 brm | 6 | 21.2 | 0.78 d | 133.0 | 5.58 brd |
| 6.90 brm | 6.90 brm | ||||||||
| 5Amp | 7 | 65.5 | 3.97 m | ||||||
| 2 | 169.2 | - | 169.3 | - | 8 | 31.6 | 1.80 m | ||
| 1.20 m | |||||||||
| 3 | 49.4 | 4.47 m | 49.4 | 4.46 m | 9 | 26.2 | 1.70 m | ||
| 0.96 m | |||||||||
| NH | 8.45 d | - | 8.47 d | ||||||
| OH | 4.63 d | ||||||||
| 4 | 21.7 | 2.40 brq | 21.9 | 2.40 brq | |||||
| 1.75 m | 1.75 m | ||||||||
| 5 | 23.8 | 2.05 m | 23.9 | 2.06 brd | 5Amp/5Ahp 2 | 169.3 | - | 169.6 | - |
| 1.70 m | 1.70 m | ||||||||
| 6 | 83.3 | 4.44 brs | 83.3 | 4.46 brs | 3 | 49.4 | 4.47 m | 48.9 | 4.45 m |
| NH | - | 7.24 m | - | 7.26 d | 4 | 21.9 | 2.40 q | 22.0 | 2.55 m |
| 1.70 m | 1.72 m | ||||||||
| OMe | 55.6 | 3.03 s | 55.7 | 3.02 s | 5 | 23.9 | 2.05 brd | 29.9 | 1.71 m |
| 1.75 m | |||||||||
| 6Val/6Ile 1 | 169.8 | - | 169.9 | - | 6 | 83.3 | 4.43 brs | 74.2 | 4.92 brs |
| 2 | 55.8 | 4.36 d | 54.1 | 4.46 m | NH | - | 7.21 m | 7.37 d | |
| 3 | 27.3 | 1.95 m | 33.1 | 1.86 m | OH/OMe | 55.5 | 3.01 s | 6.09 brs | |
| 1.10 m | |||||||||
| 4 | 18.3 | 0.46 d | 23.9 | 0.63 m | 6Ile/6Val 1 | 169.8 | - | 169.9 | - |
| 5 | 17.9 | −0.23 d | 10.4 | 0.63 m | 2 | 54.1 | 4.43 m | 56.0 | 4.32 d |
| 6 | - | - | 13.7 | −0.13 d | 3 | 32.9 | 1.78 m | 27.7 | 1.90 m |
| 7NMePhe 1 | 169.4 | - | 169.3 | - | 4 | 23.5 | 1.05 m | 18.3 | 0.46 d |
| 7ClNMeTyr | 0.59 m | ||||||||
| 2 | 61.0 | 5.10 brd | 61.0 | 5.08 brd | 5 | 10.5 | 0.59 m | 18.2 | −0.21 d |
| 3 | 34.3 | 3.30 m | 32.9 | 3.20 brd | 6 | 13.7 | −0.30 d | ||
| 2.80 m | 2.71 m | ||||||||
| 4 | 137.7 | - | 129.3 | 7NMePhe 1 | 169.3 | - | 169.3 | - | |
| 5(5′) | 129.8 | 7.23 d | 130.7 | 7.13 s | 2 | 60.9 | 5.17 dd | 60.7 | 5.08 brd |
| 6(6′) | 128.9 | 7.27 t | 120.1 | - | 3 | 34.4 | 3.30 m | 34.4 | 3.28 m |
| 2.80 dd | 2.80 m | ||||||||
| 7 | 127.0 | 7.19 t | 152.3 | - | 4 | 137.8 | - | 137.8 | - |
| 8 | - | - | 117.0 | 6.84 d | 5,5′ | 129.8 | 7.21 d | 129.9 | 7.22 m |
| 9 | - | - | 129.6 | 6.96 d | 6,6′ | 128.9 | 7.25 t | 128.8 | 7.26 m |
| NMe | 30.4 | 2.75 s | 30.4 | 2.71 s | 7 | 126.9 | 7.19 t | 126.7 | 7.19 m |
| OH | - | - | - | 9.96 s | NMe | 30.5 | 2.74 s | 30.3 | 2.73 s |
| 8Val/8Ile 1 | 172.5 | - | 172.6 | - | 8Val/8Ile 1 | 172.5 | - | 172.9 | - |
| 2 | 56.7 | 4.60 m | 54.8 | 4.79 dd | 2 | 56.5 | 4.64 dd | 55.7 | 4.77 d |
| 3 | 31.2 | 2.00 m | 37.6 | 1.80 m | 3 | 31.4 | 1.97 m | 37.7 | 1.80 m |
| 4 | 19.4 | 0.89 d | 26.0 | 1.32 m | 4 | 19.5 | 0.83 d | 24.7 | 1.26 m |
| 1.10 m | 1.00 m | ||||||||
| 5 | 18.1 | 0.83 d | 11.6 | 0.91 t | 5 | 18.0 | 0.77 d | 11.5 | 0.80 t |
| 6 | - | - | 14.1 | 0.77 d | 6 | - | - | 16.3 | 0.83 d |
| NH | - | 6.97 d | - | 6.80 d | NH | - | 6.90 d | - | 7.67 d |

2.5. Biological Activity Assessment
| Compound | Trypsin (IC50 in μM) | Chymotrypsin (IC50 in μM) |
|---|---|---|
| Aeruginosin KB676 (1) | 40.0 | >45.5 |
| Microphycin KB921 (2) | >45.5 | >45.5 |
| Anabaenopeptin KB906 (3) | >45.5 | >45.5 |
| Anabaenopeptin KB899 (4) | >45.5 | >45.5 |
| Micropeptin KB928 (5) | 0.09 | >45.5 |
| Micropeptin KB956 (6) | 0.62 | >45.5 |
| Micropeptin KB970A (7) | 0.09 | >45.5 |
| Micropeptin KB970B (8) | 0.65 | >45.5 |
| Micropeptin KB984 (9) | 1.12 | >45.5 |
| Micropeptin KB970C (10) | 4.27 | >45.5 |
| Micropeptin KB1048 (11) | 2.01 | 0.63 |
| Micropeptin KB992 (12) | >45.5 | 0.87 |
| Micropeptin KB1046 (13) | >45.5 | 0.22 |
2.6. Methods of Dereplication
3. Experimental Section
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Isolation Procedure
3.4. Determination of the Absolute Configuration of the Amino Acids by Marfey’s Method
3.5. Determination of the Absolute Configuration of Hydroxy Phenyl Lactic Acid
3.6. Protease Inhibition Assays
4. Conclusions
Supplementary Information
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zohary, T.; Roberts, R.D. Hyperscums and the population dynamics of Microcystis aeruginosa. J. Plank. Res. 1990, 12, 423–432. [Google Scholar] [CrossRef]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Doheren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Xu, Y.; Li, R. Genetic Diversity of Bloom-Forming Microcystis (Cyanobacteria) Populations in a Hyper-Eutrophic Pond in Central China. Curr. Microbiol. 2012, 65, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Moree, W.J.; Yang, J.Y.; Zhao, X.; Liu, W.T.; Aparicio, M.; Atencio, L.; Ballesteros, J.; Sánchez, J.; Gavilán, R.G.; Gutiérrez, M.; et al. Imaging Mass Spectrometry of a Coral Microbe Interaction with Fungi. J. Chem. Ecol. 2013, 39, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Traxler, M.F.; Watrous, J.D.; Alexandrov, T.; Dorrestein, P.C.; Kolter, R. Interspecies Interactions Stimulate Diversification of the Streptomyces coelicolor Secreted Metabolome. mBio 2013, 4. [Google Scholar] [CrossRef]
- Gademann, K.; Portmann, C. Secondary Metabolites from Cyanobacteria: Complex Structures and Powerful Bioactivities. Curr. Org. Chem. 2008, 12, 326–341. [Google Scholar] [CrossRef]
- Blaha, L.; Babica, P.; Marsalek, B. Toxins produced in cyanobacterial water blooms—Toxicity and risks. Interdisc. Toxicol. 2009, 2, 36–41. [Google Scholar] [CrossRef]
- Sedmak, B.; Elersek, T. Microcystins Induce Morphological and Physiological Changes in Selected Representative Phytoplanktons. Microb. Ecol. 2005, 50, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Bladt, T.T.; Kalifa-Aviv, S.; Larsen, T.O.; Carmeli, S. Micropeptins from Microcystis sp. collected in Kabul Reservoir, Israel. Tetrahedron 2014, 70, 936–943. [Google Scholar] [CrossRef]
- Raveh, A.; Carmeli, S. Aeruginazole A, a Novel Thiazole-Containing Cyclopeptide from the Cyanobacterium Microcystis sp. Org. Lett. 2010, 12, 3536–3539. [Google Scholar] [CrossRef] [PubMed]
- Adiv, S.; Aharonv-Nadborny, R.; Carmeli, S. New aeruginazoles, a group of thiazole-containing cyclic peptides from Microcystis aeruginosa blooms. Tetrahedron 2012, 68, 1376–1383. [Google Scholar] [CrossRef]
- Valls, N.; Lopez-Canet, M.; Vallribera, M.; Bonjoch, J. First Total Syntheses of Aeruginosin 298-A and Aeruginosin 298-B, Based on a Stereocontrolled Route to the New Amino Acid 6-Hydroxyoctahydroindole-2-carboxylic Acid. Chem. Eur. J. 2001, 7, 3446–3460. [Google Scholar] [CrossRef] [PubMed]
- Adiv, S.; Carmeli, S. Protease Inhibitors from Microcystis aeruginosa Bloom Material Collected from the Dalton Reservoir, Israel. J. Nat. Prod. 2013, 76, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Suzuki, S.; Ishida, K.; Murakami, M. Anabaenopeptins G and H, Potent Carboxypeptidase A inhibitors from the cyanobacterium Oscillatoria agardhii (NIES-595). Bioor. Med. Chem. Lett. 1999, 9, 1243–1246. [Google Scholar] [CrossRef]
- Okumura, H.S.; Philmus, B.; Portmann, C.; Hemscheidt, T.K. Homotyrosine-Containing Cyanopeptolins 880 and 960 and Anabaenopeptins 908 and 915 from Planktothrix agardhii CYA 126/8. J. Nat. Prod. 2009, 72, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Gesner-Apter, S.; Carmeli, S. Protease Inhibitors from a Water Bloom of the Cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2009, 72, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Zafrir-Ilan, E.; Carmeli, S. Eight novel serine proteases inhibitors from a water bloom of the cyanobacterium Microcystis sp. Tetrahedron 2010, 66, 9194–9202. [Google Scholar] [CrossRef]
- Jakobi, C.; Oberer, L.; Quiquerez, C.; Konig, W.A.; Weckesser, J. Cyanopeptolin S, a sulfate-containing depsipeptide from a water bloom of Microcystis sp. FEMS Microbiol. Lett. 1995, 129, 129–133. [Google Scholar] [PubMed]
- Jakobi, C.; Rinehart, K.L.; Neuber, R.; Mez, K.; Weckesser, J. Cyanopeptolin SS, a disulphated depsipeptide from a water bloom: Structural elucidation and biological activities. Phycologia 1996, 35, 111–116. [Google Scholar] [CrossRef]
- Zainuddin, E.N.; Mentel, R.; Wray, V.; Jansen, R.; Nimtz, M.; Lalk, M.; Mundt, S. Cyclic Depsipeptides, Ichthyopeptins A and B, from Microcystis ichthyoblabe. J. Nat. Prod. 2007, 70, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Krishanamurthy, T.; Caarmichael, W.W.; Sarver, E.W. Toxic peptides from freshwater cyanobacteria (blue-green algae). I. Isolation, purification and characterization of peptides from Microcystis aeruginosa and Anabaena flos-aquae. Toxicon 1986, 24, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Lifshits, M.; Zafrir-Ilan, E.; Raveh, A.; Carmeli, S. Protease inhibitors from three fishpond water blooms of Microcystis spp. Tetrahedron 2011, 67, 4017–4024. [Google Scholar] [CrossRef]
- Vegman, R.; Carmeli, S. Eight micropeptins from a Microcystis spp. bloom collected from a fishpond near Kibbutz Lehavot HaBashan, Israel. Tetrahedron 2013, 69, 10108–10115. [Google Scholar] [CrossRef]
- Sano, T.; Usui, T.; Ueda, K.; Osada, H.; Kaya, K. Isolation of New Protein Phosphatase Inhibitors from Two Cyanobacteria Species, Planktothrix spp. J. Nat. Prod. 2001, 64, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Lifshits, M.; Carmeli, S. Metabolites of Microcystis aeruginosa Bloom Material from Lake Kinneret, Israel. J. Nat. Prod. 2012, 75, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Marfey, P. Determination of d-amino acids. II. Use of a bifunctional reagent, 1,5-difluoro-2,4-dinitrobenzene. Carlsberg Res. Commun. 1984, 49, 591–596. [Google Scholar] [CrossRef]
- Ishida, K.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Kawaguchipeptin A, a novel cyclic undecapeptide from cyanobacterium Microcystis aeruginosa (NIES-88). Tetrahedron 1996, 52, 9025–9030. [Google Scholar] [CrossRef]
- Murakami, M.; Itou, Y.; Ishida, K.; Shin, H.J. Prenylagaramides A and B, new cyclic peptides from two strained of Oscillatoria agarghii. J. Nat. Prod. 1999, 62, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Matsuda, H.; Okita, Y.; Murakami, M. Aeruginoguanidines 98-A-98-C: Cytoxic unusual peptides from the cyanobacterium Microcystis aeruginosa. Tetrahedron 2002, 58, 7645–7652. [Google Scholar] [CrossRef]
- Liang, Z.; Sorrobas, A.; Sulzmaier, F.J.; Jimenez, J.I.; Wang, X.; Sauvage, T.; Yoshida, W.Y.; Wang, G.; Ramos, J.W.; Williams, P.G. Stictamides A–C, MMP12 inhibitors containing 4-amino-3-hydroxy-5-phenylpentanoic acid subunits. J. Org. Chem. 2011, 76, 3635–3643. [Google Scholar] [CrossRef] [PubMed]
- Gesner-Apter, S.; Carmeli, S. Three novel metabolites from a bloom of the cyanobacterium Microcystis sp. Tetrahedron 2008, 64, 6628–6634. [Google Scholar] [CrossRef]
- Baumann, H.I.; Keller, S.; Wolter, F.E.; Nicholson, G.J.; Jung, G.; Sussmuth, R.D.; Juttner, F. Planktocyclin, a cyclooctapeptide protease inhibitor produced by the freshwater cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2007, 70, 1611–1615. [Google Scholar] [CrossRef] [PubMed]
- Leikoski, N.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Rouhiainen, L.; Sivonen, K. Highly diverse cyanobactins in strains of the genus Anabaena. Appl. Environ. Microbiol. 2010, 76, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Zafrir, E.; Carmeli, S. Micropeptins from an Israeli Fishpond Water Bloom of the Cyanobacterium Microcystis sp. J. Nat. Prod. 2010, 73, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a Potent Cytotoxin and Chymotrypsin Inhibitor from the Marine Cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Okino, T.; Qi, S.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, Elastase Inhibitors from the Cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Sussmuth, R.D.; Juttner, F. Oscillapeptin J, a New Grazer Toxin of the Freshwater Cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Adiv, S.; Aharonov-Nadborny, R.; Carmeli, S. Micropeptins from Microcystis aeruginosa collected in Dalton reservoir, Israel. Tetrahedron 2010, 66, 7429–7436. [Google Scholar] [CrossRef]
- Fujii, K.; Ikai, Y.; Mayumi, T.; Oka, H.; Suzuki, M.; Harada, K.-I. A Nonempirical Method Using LC/MS for Determination of the Absolute Configuration of Constituent Amino Acids in a Peptide: Elucidation of Limitations of Marfey’s Method and of Its Separation Mechanism. Anal. Chem. 1997, 69, 3346–3352. [Google Scholar] [CrossRef]
- Ersmark, K.; del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkobi-Peer, S.; Carmeli, S. New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel. Mar. Drugs 2015, 13, 2347-2375. https://doi.org/10.3390/md13042347
Elkobi-Peer S, Carmeli S. New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel. Marine Drugs. 2015; 13(4):2347-2375. https://doi.org/10.3390/md13042347
Chicago/Turabian StyleElkobi-Peer, Shira, and Shmuel Carmeli. 2015. "New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel" Marine Drugs 13, no. 4: 2347-2375. https://doi.org/10.3390/md13042347
APA StyleElkobi-Peer, S., & Carmeli, S. (2015). New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel. Marine Drugs, 13(4), 2347-2375. https://doi.org/10.3390/md13042347