
UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium purpureum


Abstract
:1. Introduction
2. Results and Discussion
2.1. UPLC-MSE of Standard Pigments

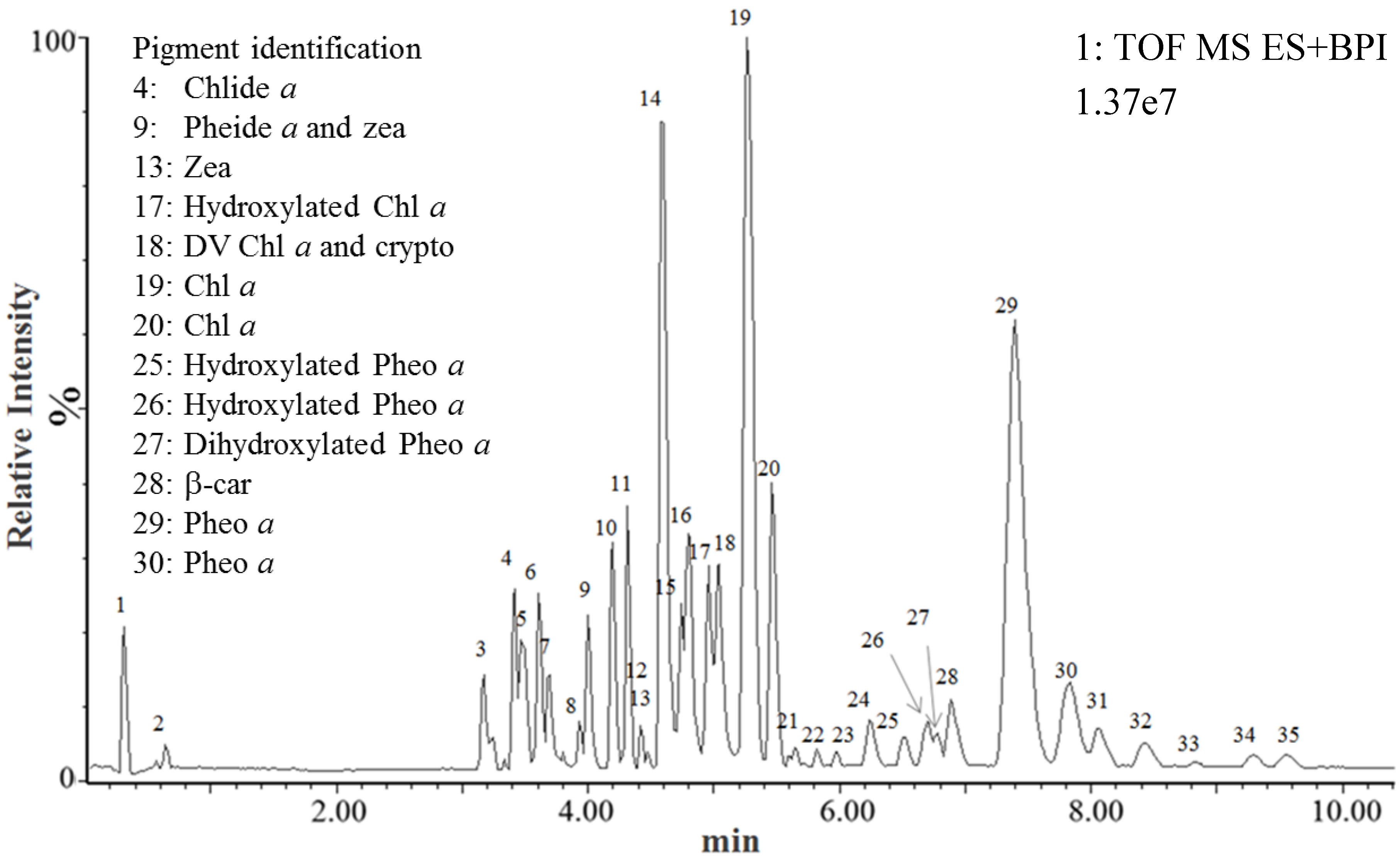{kind=link}
{kind=link}
{kind=link}
| Standard pigment | Formula | Rt (min) | Function 1 (Low Energy) | Function 2 (High Energy) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Theoretical m/z | Experimental m/z | Theoretical m/z | Experimental m/z | |||||||||||
| M●+ | [M + H]+ | [M + Na]+ | M●+ | [M + H]+ | [M + Na]+ | fragments | fragments | |||||||
| 19-Butanoyl-fucoxanthin | C46H64O8 | 3.41 | - | - | 767.4499 | - | - | 767.4491 (1.04 ppm) | 679.3999 | - | - | 679.3990 * (1.32 ppm) | - | - |
| 19-Hexanoyl-fucoxanthin | C48H68O8 | 3.61 | - | 773.4992 | 795.4812 | - | 773.4973 (2.46 ppm) | 795.4819 (0.88 ppm) | 685.4468 | 679.3999 | - | 685.4451 * (2.48 ppm) | 679.3978 * (3.09 ppm) | - |
| Alloxanthin | C40H52O2 | 3.95 | 564.3967 | - | - | 564.3965 (0.35 ppm) | - | - | 549.3733 | - | - | 549.3713 (3.64 ppm) | - | - |
| Astaxanthin | C40H52O4 | 3.75 | 596.3866 | - | 619.3763 | 596.3862 (0.67 ppm) | - | 619.3762 (0.16 ppm) | - | - | - | - | - | - |
| β-Carotene | C40H56 | 7.23 | 536.4382 | - | - | 536.4389 (1.30 ppm) | - | - | 444.3756 | - | - | 444.3755 (0.23 ppm) | - | - |
| Cryptoxanthin | C40H56O | 5.16 | 552.4331 | - | - | 552.4329 (0.36 ppm) | - | - | 460.3705 | - | - | 460.3702 (0.65 ppm) | - | - |
| Chlorophyll a | C55H72O5N4Mg | 5.28 | 892.5353 | - | 915.5251 | 892.5354 (0.11 ppm) | - | 915.5250 (0.11 ppm) | 614.2380 | 481.1879 | - | 614.2387 (1.14 ppm) | 481.1883 (0.83 ppm) | - |
| Chlorophyll a | C55H72O5N4Mg | 5.48 | 892.5353 | - | - | 892.5349 (0.45 ppm) | - | - | 614.2380 | 481.1879 | - | 614.2380 (0 ppm) | 481.1901 (4.57 ppm) | - |
| Chlorophyll b | C55H70O6N4Mg | 4.71 | 906.5146 | - | 929.5043 | 906.5149 (0.33 ppm) | - | 929.5046 (0.32 ppm) | 628.2172 | 495.1671 | - | 628.2184 (1.91 ppm) | 495.1682 (2.22 ppm) | - |
| Chlorophyll c2 | C35H28O5N4Mg | 3.62 | - | 609.1988 | 631.1808 | - | 609.1984 (0.66 ppm) | 631.1812 (0.63 ppm) | 549.1777 | - | - | 549.1765** (2.19 ppm) | - | - |
| Chlorophyllide a | C35H34O5N4Mg | 3.19 | 614.2380 | - | - | 614.2378 (0.33 ppm) | - | - | 582.2117 | 481.1879 | - | 582.2095 (3.78 ppm) | 481.1878 (0.21 ppm) | - |
| Diadinoxanthin | C40H54O3 | 3.80 | 582.4073 | - | 605.3971 | 582.4077 (0.69 ppm) | - | 605.3985 (2.31 ppm) | - | - | - | - | - | - |
| Diatoxanthin | C40H54O2 | 4,00 | 566.4124 | - | - | 566.4132 (1.41 ppm) | - | - | 119.0861 | - | - | 119.0862 (0.84 ppm) | - | - |
| DV chlorophyll a | C55H70O5N4Mg | 5.25 | 890.5197 | - | - | 890.5191 (0.67 ppm) | - | - | 612.2223 | - | - | 612.2227 (0.65 ppm) | - | - |
| DV chlorophyll b | C55H68O6N4Mg | 4.71 | 904.4989 | - | - | 904.4988 (0.11 ppm) | - | - | 626.2025 | - | - | 626.2015 (1.60 ppm) | - | - |
| Fucoxanthin | C42H58O6 | 3.48 | - | - | 681.4131 | - | - | 681.4130 (0.15 ppm) | 527.3161 | - | - | 527.3148 (2.47 ppm) | - | - |
| Peridinin | C39H50O7 | 3.15 | 630.3557 | - | 653.3454 | 630.3540 (2.7 ppm) | - | 653.3458 (0.61 ppm) | 635.3349 | 593.3243 | 575.3137 | 635.3348 * (0.16 ppm) | 593.3247 * (0.67 ppm) | 575.3130 * (1.22 ppm) |
| Pheophorbide a | C35H36O5N4 | 3.98 | - | 593.2764 | - | - | 593.2769 (0.84 ppm) | - | 533.2553 | - | - | 533.2557 ** (0.75 ppm) | - | - |
| Pheophytin a | C55H74O5N4 | 7.43 | - | 871.5737 | 893.5557 | - | 871.5734 (0.34 ppm) | 893.5560 (0.34 ppm) | 593.2764 | 533.2553 | - | 593.2768 ** (0.67 ppm) | 533.2554 ** (0.19 ppm) | - |
| Pheophytin a | C55H74O5N4 | 7.88 | - | 871.5737 | 893.5557 | - | 871.5739 (0.23 ppm) | 893.5555 (0.22 ppm) | 593.2764 | - | - | 593.2768 ** (0.67 ppm) | - | - |
| Prasinoxanthin | C40H56O4 | 3.65 | 600.4179 | - | 623.4076 | 600.4178 (0.17 ppm) | - | 623.4077 (0.16 ppm) | 469.3107 | - | - | 469.3098 (1.92 ppm) | - | - |
| Pyropheophorbide a | C33H34O3N4 | 4.34 | - | 535.2709 | - | - | 535.2714 (0.93 ppm) | - | 461.2341 | 447.2185 | - | 461.2341 ** (0 ppm) | 447.2191 ** (1.34 ppm) | - |
| Violaxanthin | C40H56O4 | 3.64 | 600.4179 | - | 623.4076 | 600.4173 (1.00 ppm) | - | 623.4077 (0.16 ppm) | 469.3083 | 221.1542 | - | 469.3082 * (0.21 ppm) | 221.1544 (0.90 ppm) | - |
| Zeaxanthin | C40H56O2 | 4.04 | 568.4280 | - | 591.4178 | 568.4273 (1.23 ppm) | - | 591.4186 (1.35 ppm) | 476.3654 | - | - | 476.3651 (0.63 ppm) | - | - |
| Zeaxanthin | C40H56O3 | 4.47 | 568.4280 | - | - | 568.4293 (2.29 ppm) | - | - | 476.3654 | - | - | 476.3664 (2.10 ppm) | - | - |
2.2. Identification of Pp Pigments Using the Standard Pigments Database

| Peak | Pigment identification | Formula | Rt (min) | Function 1 (Low energy) | Function 2 (High energy) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Theoretical m/z | Experimental m/z | Theoretical m/z | Experimental m/z | ||||||||||
| M●+ | [M + H]+ | [M + Na]+ | M●+ | [M + H]+ | [M + Na]+ | fragments | fragments | ||||||
| 4 | Chlorophyllide a | C35H34O5N4Mg | 3.40 | 614.2380 | - | - | 614.2380 (0 ppm) | - | - | 481.1879 | - | 481.1880 (0.21 ppm) | - |
| 9 | Pheophorbide a | C35H36N4O5 | 4.00 | - | 593.2764 | 615.2583 | - | 593.2783 (3.2 ppm) | 615.2594 (1.79 ppm) | 533.2553 | - | 533.2561** (1.50 ppm) | - |
| 9 | Zeaxanthin | C40H56O2 | 4.06 | 568.4280 | - | - | 568.4275 (0.88 ppm) | - | - | 476.3654 | - | 476.3635 (3.99 ppm) | - |
| 13 | Zeaxanthin | C40H56O2 | 4.47 | 568.4280 | - | - | 568.4282 (0.35 ppm) | - | - | 476.3654 | - | 476.3652 (1.89 ppm) | - |
| 18 | DV-Chlorphyll a | C55H70O5N4Mg | 5.09 | 890.5197 | - | - | 890.5206 (1.01 ppm) | - | - | 612.2223 | - | 612.2216 (1.01 ppm) | - |
| 18 | Cryptoxanthin | C40H56O | 5.17 | 552.4331 | - | - | 552.4346 (2.72 ppm) | - | - | 460.3705 | - | 460.3695 (2.17 ppm) | - |
| 19 | Chlorophyll a | C55H72O5N4Mg | 5.26 | 892.5353 | - | - | 892.5368 (1.68 ppm) | - | - | 614.2380 | 481.1879 | 614.2390 (1.63 ppm) | 481.1879 (0 ppm) |
| 20 | Chlorophyll a | C55H72O5N4Mg | 5.46 | 892.5353 | - | - | 892.5389 (4.03 ppm) | - | - | 614.2380 | 481.1879 | 614.2406 (4.23 ppm) | 481.1891 (2.49 ppm) |
| 28 | β-Carotene | C40H56 | 7.18 | 536.4382 | - | - | 536.4384 (0.37 ppm) | - | - | 444.3756 | - | 444.3752 (0.9 ppm) | - |
| 29 | Pheophytin a | C55H74O5N4 | 7.40 | - | 871.5737 | 893.5557 | - | 871.5723 (1.61 ppm) | 893.5549 (0.9 ppm) | 593.2768 | 533.2554 | 593.2755 ** (2.19 ppm) | 533.2550** (0.75 ppm) |
| 30 | Pheophytin a | C55H74O5N4 | 7.83 | - | 871.5737 | 893.5557 | - | 871.5741 (0.46 ppm) | 893.5568 (1.23 ppm) | 593.2768 | 533.2554 | 593.2770 ** (0.34 ppm) | 533.2556** (0.38 ppm) |


2.3. Identification of Pp Pigments Derivatives Using the Metabolynx Software
| Peak | Pigment identification | Formula | Rt (min) | Function 1 (Low energy) | Function 2 (High energy) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Theoretical m/z | Experimental m/z | Theoretical m/z | Experimental m/z | ||||||||||
| M●+ | [M + H]+ | [M + Na]+ | M●+ | [M + H]+ | [M + Na]+ | fragments | fragments | ||||||
| 17 | Hydroxylated chlorophyll a | C55H72O6N4Mg | 4.97 | 908.5302 | - | - | 908.5333 (3.41 ppm) | - | - | 630.2329 | - | 614.2340 (1.75 ppm) | - |
| 25 | Hydroxylated pheophytin a | C55H74O6N4 | 6.55 | - | 887.5687 | 909.5506 | - | 887.5695 (0.9 ppm) | 909.5506 (0 ppm) | 609.2713 | 549.2502 | 609.2710 ** (0.49 ppm) | 549.2491 ** (2.0 ppm) |
| 26 | Hydroxylated pheophytin a | C55H74O6N4 | 6.70 | - | 887.5687 | 909.5506 | - | 887.5676 (1.24 ppm) | 909.5495 (1.21 ppm) | 609.2713 | 549.2502 | 609.2705 ** (1.31 ppm) | 549.248 ** (4.01 ppm) |
| 27 | Dihydroxylated pheophytin a | C55H74O7N4 | 6.80 | - | 903.5636 | - | - | 903.5627 (1.00 ppm) | - | 625.2662 | 565.2451 | 625.2655 ** (1.12 ppm) | 565.2436 ** (2.65 ppm) |
2.4. Tentative Identification of Other Metabolites in the Pp Ethanol Extract
3. Experimental Section
3.1. Chemicals
| Peak | Rt (min) | Major ions ( m/z) | Empirical formula | Tentative identification | Mass error (ppm) | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | ||
| 1 | 0.32 | 277.0908 | - | C9H18O8Na | - | C9H18O8Na [M + Na]+ | - | 3.25 | - |
| 2 | 0.64 | 376.2599 | 398.2419 | C21H34N3O3 | C21H33N3O3Na | C21H34N3O3 [M + H]+ | C21H34N3O3Na [M + Na]+ | 0.27 | 0.25 |
| 3 | 3.17 | 377.2693 | 399.2515 | C23H37O4 | C23H36O4Na | Butylundecylphthalate [M + H]+ | Butylundecylphthalate [M + Na]+ | 0.27 | 1.00 |
| 4 | 3.42 | 379.2845 | 401.2672 | C23H39O4 | C23H38O4Na | 2-Arachidonoylglycerol [M + H]+a | 2-Arachidonoylglycerol [M + Na]+a | 0.79 | 1.00 |
| 5 | 3.46 | 303.2329 | 325.2136 | C20H31O2 | C20H30O2Na | Eicosapentenoic acid [M + H]+a | Eicosapentenoic acid [M + Na]+a | 1.65 | 2.15 |
| 6 | 3.61 | 353.2668 | - | C17H33N6O2 | - | C17H32N6O2 [M + H]+ | - | 0.85 | - |
| 7 | 3.68 | 305.2482 | 327.2288 | C20H33O2 | C20H32O2Na | Arachidonoic acid [M + H]+a | Arachidonoic acid [M + Na]+a | 0.33 | 3.67 |
| 8 | 3.80 | 413.2671 | - | C24H38O4Na | - | Diisooctyl phthalate [M + H]+b | - | 2.18 | - |
| 10 | 4.20 | 338.3425 | 360.3244 | C22H44NO | C22H43NONa | Erucamide [M + H]+b | Erucamide [M+Na]+b | 0.59 | 0.56 |
| 11 | 4.31 | 1007.5737 | - | C55H84O15Na | - | Digalactosyldiacylglycerol [M + Na]+ (40:5) [37,38,39,40] | - | 2.88 | - |
| 12 | 4.42 | 1009.5861 | - | C55H86O15Na | - | Digalactosyldiacylglycerol [M + Na]+ (40:9) [37,38,39,40] | - | 0.30 | - |
| 13 | 4.47 | 947.5701 | - | C50H84O15Na | - | Digalactosyldiacylglycerol [M + Na]+ (35:5) [37,38,39,40] | - | 0.74 | - |
| 14 | 4.61 | 961.5868 | - | C51H86O15Na | - | Digalactosyldiacylglycerol [M + Na]+ (36:5) [37,38,39,40] | - | 0.42 | - |
| 15 | 4.74 | 963.6005 | - | C51H88O15Na | - | Digalactosyldiacylglycerol [M + Na]+ (36:4) [37,38,39,40] | - | 1.66 | - |
| 16 | 4.80 | 685.4784 | 939.5999 | C39H62N6O3Na | C49H88O15Na | C39H62N6O3Na [M + Na]+ | Digalactosyldiacylglycerol [M + Na]+ (34:3) [37,38,39,40] | 0.44 | 2.34 |
| 17 | 4.97 | 687.4990 | - | C43H68O5Na | - | Diacylglycerol [M + Na]+ (40:8)a [38,39] | - | 3.78 | - |
| 18 | 5.03 | 637.4825 | - | C39H66O5Na | - | Diacylglycerol [M + Na]+ (40:8)a [38,39] | - | 2.67 | - |
| 21 | 5.64 | 591.4996 | - | C37H67O5 | - | Diacylglycerol [M + H]+ (34:3)a [38,39] | - | 1.18 | - |
| 22 | 5.82 | 726.6608 | 748.6443 | C44H88NO6 | C44H87NO6Na | Gracilamide B (+ OH -H + CH2) [M + H]+ [41] | Gracilamide B (+ OH -H + CH2) [M+Na]+ [41] | 0.69 | 1.60 |
| 23 | 5.98 | 696.6515 | 718.6335 | C43H86NO5 | C43H85NO5Na | Gracilamide B [M + H]+ [41] | Gracilamide B [M+Na]+ [41] | 1.29 | 1.39 |
| 24 | 6.24 | 710.6680 | 732.6506 | C44H88NO5 | C44H87NO5Na | Gracilamide B (+ CH2) [M + H]+ [41] | Gracilamide B (+ CH2) [M + Na]+ [41] | 2.53 | 3.28 |
| 28 | 6.87 | 967.6772 | - | C63H92O6Na | - | Triacylglycerol [M + Na]+ (60:15)a [39] | - | 2.07 | - |
| 31 | 8.04 | 971.7145 | - | C63H96O6Na | - | Triacylglycerol [M + Na]+ (60:13)a [39] | - | 4.12 | - |
| 32 | 8.44 | 921.6916 | - | C59H94O6Na | - | Triacylglycerol [M + Na]+ (56:10) a [39] | - | 3.47 | - |
| 33 | 8.81 | 973.7232 | - | C63H98O6Na | - | Triacylglycerol [M + Na]+ (60:12) a [39] | - | 2.98 | - |
| 34 | 9.29 | 923.7145 | - | C59H96O6Na | - | Triacylglycerol [M + Na]+ (56:9) a [39] | - | 4.33 | - |
| 35 | 9.52 | 771.6071 | - | C53H80O2Na | - | Plastoquinone [M + Na]+ | - | 1.94 | - |
3.2. Microalgae
3.3. Pigment Extraction
3.4. UPLC-MSE
3.4.1. Equipment and Analytical Conditions
3.4.2. Software
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Abbreviations
| 19′ But-fuco | 19′ Butanoyloxy-fucoxanthin |
| 19′ Hexan-fuco | 19′ Hexanoyloxy-fucoxanthin |
| Allo | Alloxanthin |
| Asta | Astaxanthin |
| Chl | Chlorophyll |
| Chlide a | Chlorophyllide a |
| Diadino | Diadinoxanthin |
| Diato | Diatoxanthin |
| DV chl | Divinylchlorophyll |
| Fuco | Fucoxanthin |
| HPLC | High Performance Liquid Chromatography |
| MSE | High-resolution mass spectrometry with simultaneous acquisition of accurate mass at high and low collision energy |
| Perid | Peridinin |
| Pheide a | Pheophorbide a |
| Phein a | Pheophytin a |
| Pp | Porphyridium purpureum |
| Prasino | Prasinoxanthin |
| Pyropheide a | Pyropheophorbide a |
| UPLC | Ultra-Performance Liquid Chromatography |
| UV-vis | Ultraviolet-Visible |
| Viola | Violaxanthin |
| Zea | zeaxanthin |
| β-car | β,β-carotene |
| crypto | cryptoxanthin |
Conflicts of Interest
References
- Wright, S.W.; Jeffrey, S.W. Pigment markers for phytoplankton production. In Marine Organic Matter: Biomarkers, Isotopes and DNA; Volkman, J.K., Ed.; The Handbook of Environmental Chemistry; Springer: Berlin Heidelberg, Germany, 2006; pp. 71–104. [Google Scholar]
- Jeffrey, S.W.; Wright, S.W.; Zapata, M. Recent advances in HPLC pigment analysis of phytoplankton. Mar. Freshw. Res. 1999, 50, 879–896. [Google Scholar] [CrossRef]
- Medlin, L.K.; Lange, M.; Nothig, E.M. Genetic diversity in the marine phytoplankton: A review and a consideration of Antarctic phytoplankton. Antarct. Sci. 2000, 12, 325–333. [Google Scholar] [CrossRef]
- Barra, L.; Chandrasekaran, R.; Corato, F.; Brunet, C. The challenge of ecophysiological biodiversity for biotechnological applications of marine microalgae. Mar. Drugs 2014, 12, 1641–1675. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of high added-value compounds—A brief review of recent work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef]
- Szymczak-Żyła, M.; Kowalewska, G.; Louda, J.W. Chlorophyll-a and derivatives in recent sediments as indicators of productivity and depositional conditions. Mar. Chem. 2011, 125, 39–48. [Google Scholar] [CrossRef]
- Reuss, N.; Conley, D.J.; Bianchi, T.S. Preservation conditions and the use of sediment pigments as a tool for recent ecological reconstruction in four Northern European estuaries. Mar. Chem. 2005, 95, 283–302. [Google Scholar] [CrossRef]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.-P.; Morant-Manceau, A.; Schoefs, B. The potential of microalgae for the production of bioactive molecules of pharmaceutical interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef] [PubMed]
- Gagez, A.-L.; Thiery, V.; Pasquet, V.; Cadoret, J.-P.; Picot, L. Epoxycarotenoids and Cancer. Review. Curr. Bioact. Compd. 2012, 8, 109–141. [Google Scholar] [CrossRef]
- Pasquet, V.; Morisset, P.; Ihammouine, S.; Chepied, A.; Aumailley, L.; Berard, J.-B.; Serive, B.; Kaas, R.; Lanneluc, I.; Thiery, V.; et al. Antiproliferative activity of violaxanthin isolated from bioguided fractionation of Dunaliella tertiolecta extracts. Mar. Drugs 2011, 9, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Baudelet, P.-H.; Gagez, A.-L.; Bérard, J.-B.; Juin, C.; Bridiau, N.; Kaas, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. Antiproliferative activity of Cyanophora paradoxa pigments in melanoma, breast and lung cancer cells. Mar. Drugs 2013, 11, 4390–4406. [Google Scholar] [CrossRef] [PubMed]
- Juin, C.; Chérouvrier, J.-R.; Thiéry, V.; Gagez, A.-L.; Bérard, J.-B.; Joguet, N.; Kaas, R.; Cadoret, J.-P.; Picot, L. Microwave-assisted extraction of phycobiliproteins from Porphyridium purpureum. Appl. Biochem. Biotechnol. 2015, 175, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A marine carotenoid exerting anti-cancer effects by affecting multiple mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; D’Orazio, N.; Franceschelli, S.; Speranza, L. Marine carotenoids and cardiovascular risk markers. Mar. Drugs 2011, 9, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; de Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.W.; Wright, S.W.; Zapata, M. Microalgal classes and their signature pigments. In Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Roy, S., Llewellyn, C., Egeland, E.S., Johnsen, G., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 3–77. [Google Scholar]
- Phytoplankton Pigments : Characterization, Chemotaxonomy and Applications in Oceanography; Roy, S.; Llewellyn, C.; Egeland, E.; Johnsen, G. (Eds.) Cambridge University Press: Cambridge, UK, 2011.
- Wright, S.W. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Lewitus, J.; Kana, T.M.; Craft, N.E. Improved separations of phytoplankton pigments using temperature controlled high performance liquid chromatography. Mar. Ecol. Prog. Ser. 1994, 114, 303–313. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Lewitus, A.J.; Kana, T.M.; Craft, N.E. High-performance liquid chromatography of phytoplankton pigments using a polymeric reversed-phase C18 column. J. Phycol. 1992, 28, 867–872. [Google Scholar] [CrossRef]
- Zapata, M.; Rodriguez, F.; Garrido, J. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Bonjean, K.; De Pauw-Gillet, M.C.; Defresne, M.P.; Colson, P.; Houssier, C.; Dassonneville, L.; Bailly, C.; Greimers, R.; Wright, C.; Quetin-Leclercq, J.; et al. The DNA intercalating alkaloid cryptolepine interferes with topoisomerase II and inhibits primarily DNA synthesis in B16 melanoma cells. Biochemistry 1998, 37, 5136–5146. [Google Scholar] [CrossRef] [PubMed]
- Gentili, A.; Caretti, F. Evaluation of a method based on liquid chromatography-diode array detector-tandem mass spectrometry for a rapid and comprehensive characterization of the fat-soluble vitamin and carotenoid profile of selected plant foods. J. Chromatogr. A 2011, 1218, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Raghunath Reddy, R.L.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. Characterization of microalgal carotenoids by mass spectrometry and their bioavailability and antioxidant properties elucidated in rat model. J. Agric. Food Chem. 2010, 58, 8553–8559. [Google Scholar] [CrossRef] [PubMed]
- Ranga, R.; Sarada, A.R.; Baskaran, V.; Ravishankar, G.A. Identification of carotenoids from green alga Haematococcus pluvialis by HPLC and LC-MS (APCI) and their antioxidant properties. J. Microbiol. Biotechnol. 2009, 19, 1333–1341. [Google Scholar] [PubMed]
- Rivera, S.; Vilaró, F.; Canela, R. Determination of carotenoids by liquid chromatography/mass spectrometry: Effect of several dopants. Anal. Bioanal. Chem. 2011, 400, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry. Mass Spectrom. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, Y.; Zhao, C.; Zhou, P.; Yu, L. Analysis and identification of astaxanthin and its carotenoid precursors from Xanthophyllomyces dendrorhous by high-performance liquid chromatography. Z. Naturforsch. C. 2010, 65, 489–94. [Google Scholar] [PubMed]
- Fu, W.; Magnúsdóttir, M.; Brynjólfson, S.; Palsson, B.Ø.; Paglia, G. UPLC-UV-MS(E) analysis for quantification and identification of major carotenoid and chlorophyll species in algae. Anal. Bioanal. Chem. 2012, 404, 3145–3154. [Google Scholar] [CrossRef]
- Khachik, F.; Bernstein, P.S.; Garland, D.L. Identification of lutein and zeaxanthin oxidation products in human and monkey retinas. Invest. Ophthalmol. Vis. Sci. 1997, 38, 1802–1811. [Google Scholar] [PubMed]
- Kopecký, J.; Lukavská, A.; Verboviková, E.; Pfündel, E. Changes in the photosynthetic pigment patterns during the synchronous life cycle of Porphyridium purpureum. Arch. Hydrobiol. Suppl. Algol. Stud. 2004, 114, 121–132. [Google Scholar]
- Cunningham, F.X.; Dennenberg, R.J.; Jursinic, P.A.; Gantt, E. Growth under red light enhances photosystem II relative to photosystem I and phycobilisomes in the red alga Porphyridium cruentum. Plant Physiol. 1990, 93, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Bouman, H.A.; Ulloa, O.; Scanlan, D.J.; Zwirglmaier, K.; Li, W.K.W.; Platt, T.; Stuart, V.; Barlow, R.; Leth, O.; Clementson, L.; et al. Oceanographic basis of the global surface distribution of Prochlorococcus ecotypes. Science 2006, 312, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Partensky, F.; La Roche, J.; Wyman, K.; Falkowski, P.G. The divinyl-chlorophyll a/b-protein complexes of two strains of the oxyphototrophic marine prokaryote Prochlorococcus—Characterization and response to changes in growth irradiance. Photosynth. Res. 1997, 51, 209–222. [Google Scholar] [CrossRef]
- Barlow, R.G.; Cummings, D.G.; Gibb, S.W. Improved resolution of mono- and divinyl chlorophylls a and b and zeaxanthin and lutein in phytoplankton extracts using reverse phase C-8 HPLC. Mar. Ecol. Prog. Ser. 1997, 161, 303–307. [Google Scholar] [CrossRef]
- Dahmen, J.; Khadka, M.; Dodson, J.; Leblond, J. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. VI. Biochemical and genomic comparison of galactolipid biosynthesis between Chromeravelia (Chromerida), a photosynthetic alveolate with a red algal plastid ancestry, to the dinoflagellate, Lingulodinium polyedrum. Eur. J. Phycol. 2013, 48, 268–277. [Google Scholar] [CrossRef]
- Botté, C.Y.; Yamaryo-Botté, Y.; Janouskovec, J.; Rupasinghe, T.; Keeling, P.J.; Crellin, P.; Coppel, R.L.; Maréchal, E.; McConville, M.J.; McFadden, G.I. Identification of plant-like galactolipids in Chromeravelia, a photosynthetic relative of malaria parasites. J. Biol. Chem. 2011, 286, 29893–29903. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.K.; Gautam, K.; Jueppner, J.; Giavalisco, P.; Rihko-Struckmann, L.; Pareek, A.; Sundmacher, K. UPLC-MS analysis of Chlamydomonas reinhardtii and Scenedesmus obliquus lipid extracts and their possible metabolic roles. J. Appl. Phycol. 2014. [Google Scholar] [CrossRef]
- Leblond, J.D.; Timofte, H.I.; Roche, S.A.; Porter, N.M. Mono- and digalactosyldiacylglycerol composition of glaucocystophytes (Glaucophyta): A modern interpretation using positive-ion electrospray ionization/mass spectrometry/mass spectrometry. Phycol. Res. 2010, 58, 222–229. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, Y.; Liu, K.; Hua, H.; Zhu, H.; Pei, Y. Gracilarioside and gracilamides from the red alga Gracilaria asiatica. J. Nat. Prod. 2006, 69, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Walne, P. Studies on the Food Value of Nineteen Genera of Algae to Juvenile Bivalves of the Genera Ostrea, Crassostrea, Mercenaria and Mytilus; H.M.S.O.: London, UK, 1970. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juin, C.; Bonnet, A.; Nicolau, E.; Bérard, J.-B.; Devillers, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium purpureum. Mar. Drugs 2015, 13, 2541-2558. https://doi.org/10.3390/md13042541
Juin C, Bonnet A, Nicolau E, Bérard J-B, Devillers R, Thiéry V, Cadoret J-P, Picot L. UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium purpureum. Marine Drugs. 2015; 13(4):2541-2558. https://doi.org/10.3390/md13042541
Chicago/Turabian StyleJuin, Camille, Antoine Bonnet, Elodie Nicolau, Jean-Baptiste Bérard, Romain Devillers, Valérie Thiéry, Jean-Paul Cadoret, and Laurent Picot. 2015. "UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium purpureum" Marine Drugs 13, no. 4: 2541-2558. https://doi.org/10.3390/md13042541
APA StyleJuin, C., Bonnet, A., Nicolau, E., Bérard, J. -B., Devillers, R., Thiéry, V., Cadoret, J. -P., & Picot, L. (2015). UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium purpureum. Marine Drugs, 13(4), 2541-2558. https://doi.org/10.3390/md13042541