
Photosynthetic Pigments in Diatoms

Abstract
:1. Introduction
2. Physical and Chemical Properties of Photosynthetic Pigments of Diatoms
2.1. Chlorophylls

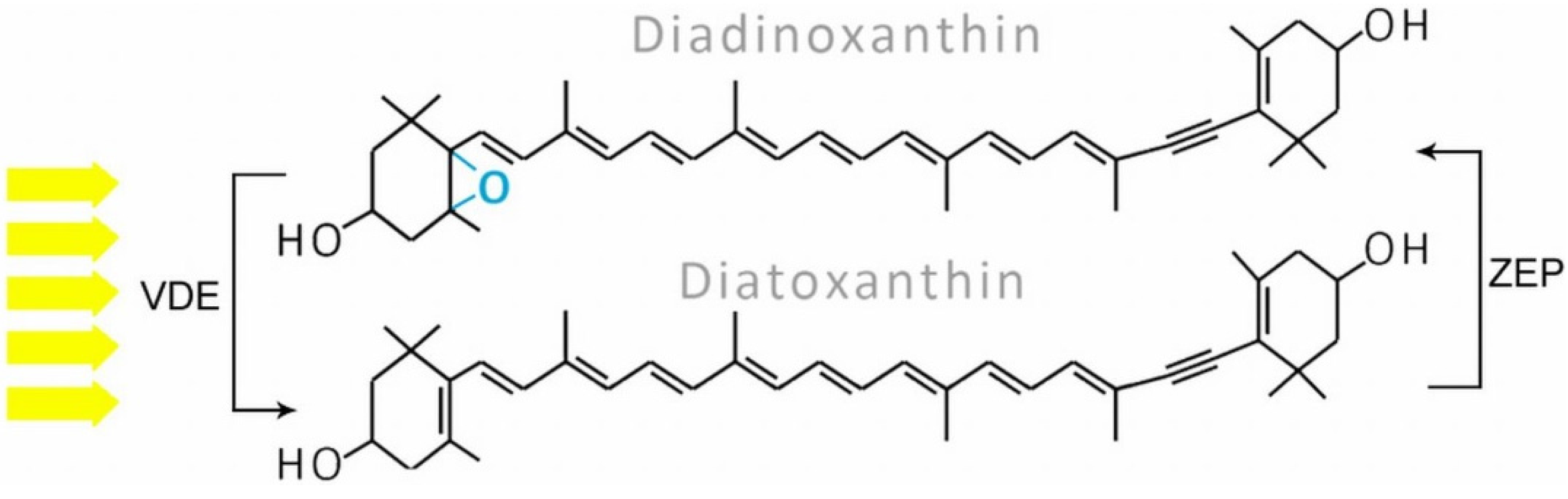
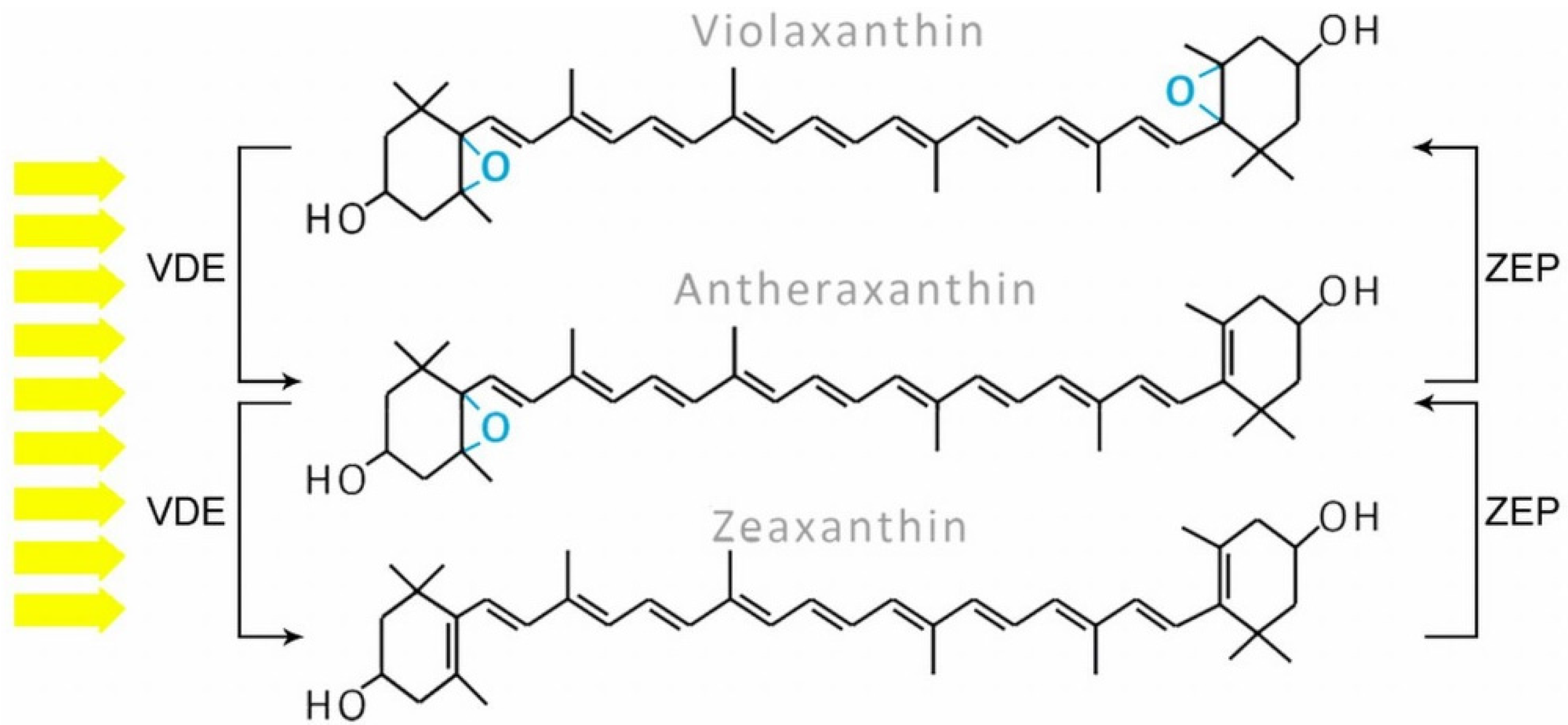
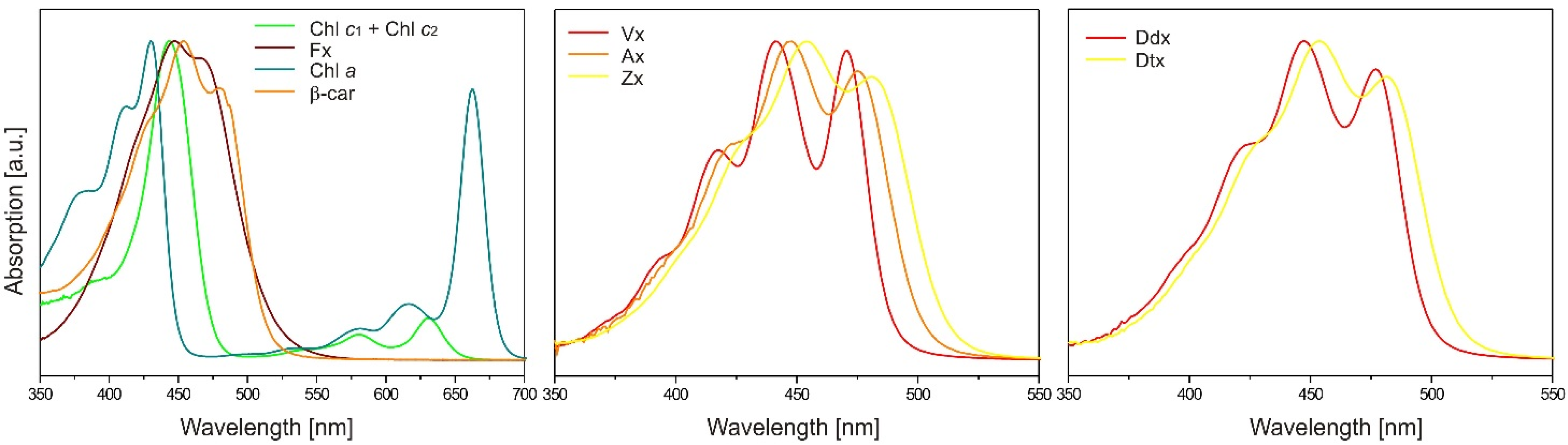
2.2. Carotenoids
3. Biosynthesis Pathways

4. Regulation of Pigment Level in the Cell

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Species | Changes in Pigment Content | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chl a | Chl c | β-Car | Fx | Ddx | Dtx | Vx | Ax | Zx | ||
| HL (140 μmol photons m−2·s−1) in comparison to LL (40 μmol photons m−2·s−1), 16 h light/8 h dark photoperiod [51] | Cyclotella meneghiniana | N/A | const | const | const | ↑ | ↑ | N/A | N/A | N/A |
| Iron-replete medium (12 μM) compared to iron-reduced medium (1 μM) in HL (140 μmol photons m−2·s−1) [52] | Cyclotella meneghiniana | ↑ | N/A | N/A | N/A | ↓ | N/A | N/A | N/A | |
| Iron-replete medium (12 μM) compared to iron-reduced medium (1 μM) in LL (40 μmol photons m−2·s−1) [52] | Cyclotella meneghiniana | ↑ | N/A | N/A | N/A | const | N/A | N/A | N/A | |
| HL (300 μmol photons m−2·s−1) compared to LL (50 μmol photons m−2·s−1), 14 h light/10 h dark photoperiod [53] | Phaeodactylum tricornutum | N/A | ↓ | ↑ | ↓ | ↑ | ↑ | N/A | N/A | N/A |
| B-HL (450 PFD) compared to BR-HL (450 PFD in R:B ratio 0.25) [43] | Pseudonitzschia multistriata | ↓ | ↑ | const | ↓ | ↓ | ↓ | N/A | N/A | N/A |
| B-LL (250 PFD) compared to BR-LL (250 PFD in R:B ratio 0.25) [43] | Pseudonitzschia multistriata | ↓ | ↑ | const | ↓ | ↓ | ↓ | N/A | N/A | N/A |
| B-LL (24 (10 absorbed) μmol photons m−2·s−1) compared to W-LL (40 (10 absorbed) μmol photons m−2·s−1) [54] | Phaeodactylum tricornutum | ↑ | const | const | const | const | N/A | ↓ | N/A | N/A |
| R-LL (41 (10 absorbed) μmol photons m−2·s−1) compared to W-LL (40 (10 absorbed) μmol photons m−2 s−1) [54] | Phaeodactylum tricornutum | ↑ | ↓ | ↑ | ↓ | ↓ | N/A | ↓ | N/A | N/A |
| HL (1250 μmol photons m−2·s−1) in comparison to LL (40 μmol photons m−2·s−1), 12 h light/12 h dark photoperiod [55] | Phaeodactylum tricornutum | ↓ | const | const | const | ↓ | ↑ | N/A | N/A | N/A |
| 6 days acclimated to shift from BL (24 (10 absorbed) μmol photons m−2·s−1) to RL (40 (10 absorbed) μmol photons m−2·s−1) [45] | Phaeodactylum tricornutum | ↑ | N/A | N/A | N/A | ↓ | N/A | N/A | N/A | N/A |
| 6 days acclimated to shift from RL (40 (10 absorbed) μmol photons m−2·s−1) to BL (24 (10 absorbed) μmol photons m−2·s−1) [45] | Phaeodactylum tricornutum | const | N/A | N/A | N/A | ↑ | N/A | N/A | N/A | N/A |
| 14 days dark storage culture [56] | Thalassiosira weissflogii | ↓ | ↓ | ↓ | ↓ | ↓ | N/A | N/A | N/A | |
| HL (700 μmol photons m−2·s−1) in comparison to LL (40 μmol photons m−2·s−1), 16 h light/8 h dark photoperiod [24] | Cyclotella meneghiniana | N/A | N/A | N/A | N/A | ↓ | ↑ | const | ↑ | ↑ |
| high nitrogen culture (18 mM) compared to low nitrogen culture (6 mM) in LL (100 μmol photons m−2·s−1) [8] | Odontella aurita | N/A | N/A | N/A | ↑ | N/A | N/A | N/A | N/A | N/A |
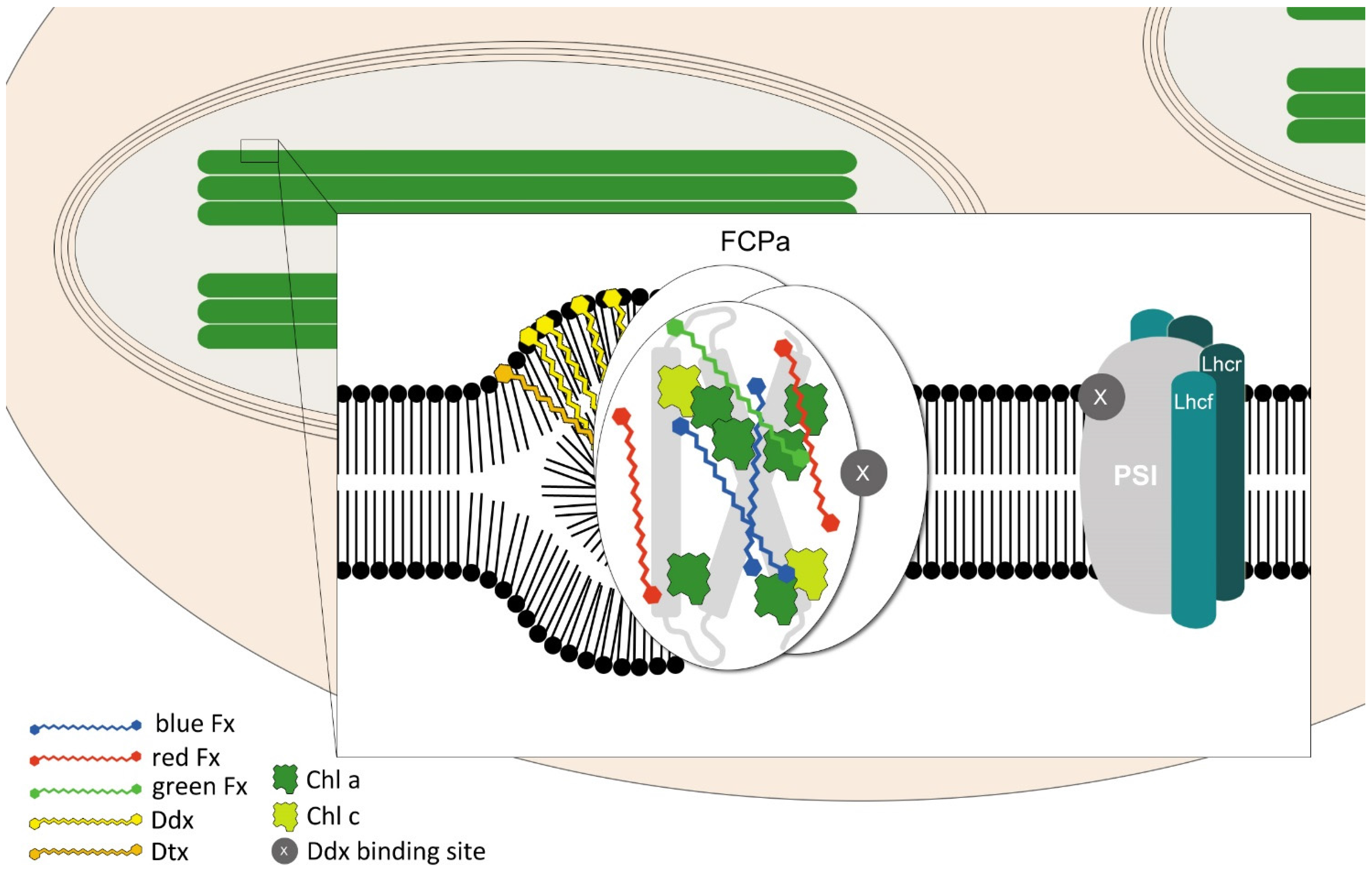
5. Localization in the Cell

6. Pigments Involved in Photosynthesis
| Species | Morphology | Colony-Forming | Lifestyle | Habitat |
|---|---|---|---|---|
| Actinella punctata | eunotioid | yes | benthic | acidic, humic lakes, and ponds |
| Actinocyclus normanii | centric | no | planktonic | coasts, brackish waters, sediment core |
| Amphora minutissima | asymmetrical biraphid | no | benthic | marine habitats, often epiphytic |
| Bacillaria paradoxa | nitzschioid | yes | benthic | marine, brackish, and freshwaters |
| Campylodiscus hibernicus | surirelloid | no | benthic | epipelon in fresh, brackish, marine waters |
| Cocconeis pediculus | monoraphid | no | benthic | planktonic, epiphytic, epilithic habitats |
| Cyclotella distinguenda | centric | no | planktonic | preferentially alkaline waters |
| Cymbella amplificata | asymmetrical biraphid | yes | benthic | oligotrophic waters |
| Diatoma vulgaris | araphid | yes | benthic | fresh and brackish water |
| Discostella stelligera | centric | no | planktonic | primarily in lakes and large rivers |
| Distrionella incognita | araphid | yes | benthic | alkaline lakes and streams |
| Epithemia turgida | epithemioid | no | benthic | epiphyte on coarse filamentous algae |
| Eucocconeis alpestris | monoraphid | no | benthic | the littoral zone of oligotrophic lakes |
| Eunotia exigua | eunotioid | N/A | benthic | moist soils, wet walls, streams, waterfalls |
| Fragilaria crotonensis | araphid | yes | planktonic | mesotrophic lakes, water column |
| Gyrosigma acuminatum | symmetrical biraphid | no | benthic | primarily an epipelic species |
| Navicula reinhardtii | symmetrical biraphid | no | benthic | fresh water, slightly brackish |
| Nitzschia regula | nitzschioid | no | benthic | cold-water, ponds, and streams |
| Phaeodactylum tricornutum | fusiform, triradiate, oval | no | planktonic | marine coastal waters |
| Pinnularia rabenhorstii | symmetrical biraphid | no | benthic | cold oligotrophic waters in the mountains |
| Pleurosira laevis | centric | yes | benthic | naturally saline or polluted waters |
| Thalassiosira weissflogii | centric | N/A | planktonic | primarily in marine waters |

7. Pigments Involved in Photoprotection


8. Methods of Pigment Analysis and Identification
8.1. Extraction of Pigments
8.2. HPLC Analysis of Pigments


| Pigment | Literature Data | HPLC Data * | ||||
|---|---|---|---|---|---|---|
| Solvent | λmax (nm) | E (L g−1·cm−1) | Reference | λmax (nm) | Retention Time (min) | |
| Chlorophyll c1 | acetone (90%) | 443 | 318 | [120] | ||
| Chlorophyll c2 | acetone (90%) | 444 | 374 | [120] | ||
| Chlorophyll c1 + c2 | N/A | N/A | N/A | N/A | 443 | 4.7 |
| Fucoxanthin | petrol ether | 449 | 165 | [121] | 447 | 7.0 |
| ethanol | 450 | 114 | [122] | |||
| acetone | 443 | 165 | [123] | |||
| Violaxanthin | N/A | 443 | N/A | [114] | 441 | 8.4 |
| Diadinoxanthin | acetone | 448 | 224 | [124] | 447 | 9.9 |
| methanol | 445 | 225 | [124] | |||
| hexane | 446 | 211 | [124] | |||
| Anteraxanthin | ethanol | 446 | 235 | [125] | 447 | 10.8 |
| Diatoxanthin | acetone | 454 | N/A | [114] | 454 | 11.3 |
| Zeaxanthin | ethanol | 452 | 254 | [126] | 453 | 11.9 |
| Chlorophyll a | acetone | 662 | 88.15 | [127] | 429 | 14.9 |
| β-carotene | etanol | 453 | 262 | [128] | 454 | 18.7 |
| hexane | 453 | 259.2 | [128] | |||
| acetone | 454 | 250 | [129] | |||
9. Significance of Diatom Pigments
10. Methods of Purification of Diatom Pigments
11. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Bozarth, A.; Maier, U.G.; Zauner, S. Diatoms in biotechnology: Modern tools and applications. Appl. Microbiol. Biotechnol. 2009, 82, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolisim in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Allen, A.E.; Dupont, C.L.; Oborník, M.; Horák, A.; Nunes-Nesi, A.; McCrow, J.P.; Zheng, H.; Johnson, D.A.; Hu, H.; Fernie, A.R.; et al. Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature 2011, 473, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef] [PubMed]
- Perl, T.M.; Bédard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.; Remis, R.S. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med. 1990, 322, 1775–1780. [Google Scholar] [CrossRef] [PubMed]
- Prestegard, S.K.; Oftedal, L.; Nygaard, G.; Skjaerven, K.H.; Knutsen, G.; Døskeland, S.O.; Coyne, R.T.; Herfindal, L. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-T. Delta 5 Fatty Acid Desaturase Upregulates the Synthesis of Polyunsaturated Fatty Acids in the Marine Diatom Phaeodactylum tricornutum. J. Agric. Food Chem. 2014, 62, 8773–8776. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.B.; Morançais, M.; Fleurence, J.; Windarto, E.; Arsad, S.; Prasetiya, F.S.; Jaouen, P.; Babin, M.; et al. Marennine, promising blue pigments from a widespread Haslea diatom species complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Davidovich, N.A.; Bardeau, J.F.; Caruso, A.; Leignel, V.; Hardivillier, Y.; Rince, Y.; Jacquette, B.; Davidovich, O.I.; Gaudin, P.; et al. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the Black Sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar] [CrossRef]
- Gelzinis, A.; Butkus, V.; Songaila, E.; Augulis, R.; Gall, A.; Büchel, C.; Robert, B.; Abramavicius, D.; Zigmantas, D.; Valkunas, L. Mapping energy transfer channels in fucoxanthin-chlorophyll protein complex. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.; Vesk, M. Introduction to marine phytoplankton and their pigment signatures. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S., Mantoura, R., Wright, S., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 37–84. [Google Scholar]
- Zapata, M.; Garrido, J.; Jeffrey, S. Chlorophyll c Pigments: Current Status. In Chlorophylls and Bacteriochlorophylls; Grimm, B., Porra, R., Rüdiger, W., Scheer, H., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2006; pp. 39–53. [Google Scholar]
- Stauber, J.L.; Jeffrey, S.W. Photosynthetic pigments in fifty-one species of marine diatoms. J. Phycol. 1988, 24, 158–172. [Google Scholar] [CrossRef]
- Fookes, C.; Jeffrey, S. The structure of chlorophyll c3, a novel marine photosynthetic pigment. J. Chem. Soc. Chem. Commun. 1989, 23, 1827–1828. [Google Scholar] [CrossRef]
- Britton, G. UV/visible spectroscopy. In Carotenoids: Spectroscopy, Vol 1B; Liaaen-Jensen, S., Britton, G., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; pp. 13–63. [Google Scholar]
- Zigmantas, D.; Hiller, R.G.; Sharples, F.P.; Frank, H.A.; Sundstrom, V.; Polivka, T. Effect of a conjugated carbonyl group on the photophysical properties of carotenoids. Phys. Chem. Chem. Phys. 2004, 6, 3009–3016. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Beker, B.; Raphel, D.; Baillet, G. Photodegradation of chlorophyll phytyl chain in dead phytoplanktonic cells. J. Photochem. Photobiol. A Chem. 1995, 85, 137–142. [Google Scholar] [CrossRef]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids, 1st ed.; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2004. [Google Scholar]
- Subczynski, W.K.; Markowska, E.; Sielewiesiuk, J. Effect of polar carotenoids on the oxygen diffusion-concentration product in lipid bilayers. An EPR spin label study. Biochim. Biophys. Acta Biomembr. 1991, 1068, 68–72. [Google Scholar] [CrossRef]
- Berglund, A.H.; Nilsson, R.; Liljenberg, C. Permeability of large unilamellar digalactosyldiacylglycerol vesicles for protons and glucose—Influence of α-tocopherol, β-carotene, zeaxanthin and cholesterol. Plant Physiol. Biochem. 1999, 37, 179–186. [Google Scholar] [CrossRef]
- Refvem, T.; Strand, A.; Kjeldstad, B.; Haugan, J.A.; Liaaen-Jensen, S. Stereoisomerization of Allenic Carotenoids—Kinetic, Thermodynamic and Mechanistic Aspects. Acta Chem. Scand. 1999, 53, 114–123. [Google Scholar] [CrossRef]
- Haugan, J.A.; Englert, G.; Liaaen-Jensen, S.; Frimpong-Manso, S.; Springborg, J.; Wang, D.-N.; Christensen, S.B. Algal Carotenoids. 50. Alkali Lability of Fucoxanthin—Reactions and Products. Acta Chem. Scand. 1992, 46, 614–624. [Google Scholar] [CrossRef]
- Lohr, M.; Wilhelm, C. Algae displaying the diadinoxanthin cycle also possess the violaxanthin cycle. Proc. Natl. Acad. Sci. USA 1999, 96, 8784–8789. [Google Scholar] [CrossRef] [PubMed]
- Frank, H.A.; Cogdell, R.J. Carotenoids in photosynthesis. Photochem. Photobiol. 1996, 63, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef] [PubMed]
- Lohr, M.; Wilhelm, C. Xanthophyll synthesis in diatoms: Quantification of putative intermediates and comparison of pigment conversion kinetics with rate constants derived from a model. Planta 2001, 212, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, M. Carotenoid biosynthesis in diatoms. Photosynth. Res. 2010, 106, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Coesel, S.; Oborník, M.; Varela, J.; Falciatore, A.; Bowler, C. Evolutionary origins and functions of the carotenoid biosynthetic pathway in marine diatoms. PLoS ONE 2008, 3, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Stransky, H.; Hager, A. Das Carotinoidmuster und die Verbreitung des lichtinduzierten Xanthophyllcyclus in verschiedenen Algenklassen. Arch. Mikrobiol. 1970, 73, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Montsant, A.; Allen, A.E.; Coesel, S.; Mangogna, M.; Siaut, M.; Heijde, M.; Jabbari, K.; Maheswari, U.; Rayko, E.; Vardi, A.; et al. Identification and comparative genomic analysis of signaling and regulatory components in the diatom Thalassiosira pseudonana. J. Phycol. 2007, 43, 585–604. [Google Scholar] [CrossRef]
- Dambek, M.; Eilers, U.; Bretenbach, J.; Steiger, S.; Büchel, C.; Sandmann, G. Biosynthesis of fucoxanthin and diadinoxanthin and function of initial pathway genes in Phaeodactylum tricornutum. J. Exp. Bot. 2012, 63, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K.; Hosokawa, M. Biosynthetic pathway and health benefits of fucoxanthin, an algae-specific xanthophyll in brown seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [PubMed]
- Grimm, B.; Porra, R.J.; Rüdiger, W.; Scheer, H. Chlorophylls and Bacteriochlorophylls: Biochemistry, Biophysics, Functions and Applications, 1st ed.; Grimm, B., Porra, R.J., Rüdiger, W., Scheer, H., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2006; Volume 25. [Google Scholar]
- Hunsperger, H.M.; Randhawa, T.; Cattolico, R.A. Extensive horizontal gene transfer, duplication, and loss of chlorophyll synthesis genes in the algae. BMC Evol. Biol. 2015, 15, 1–19. [Google Scholar]
- Disch, A.; Schwender, J.; Muller, C.; Lichtenthaler, H.K.; Rohmer, M. Distribution of the mevalonate and glyceraldehyde phosphate/pyruvate pathways for isoprenoid biosynthesis in unicellular algae and the cyanobacterium Synechocystis PCC 6714. Biochem. J. 1998, 333, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Ohtsuka, T.; Tanaka, A. Conversion of chlorophyll b to chlorophyll a via 7-hydroxymethyl chlorophyll. J. Biol. Chem. 1996, 271, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J. Recent Progress in Porphyrin and Chlorophyll Biosynthesis. Photochem. Photobiol. 1997, 65, 492–516. [Google Scholar] [CrossRef]
- Vidoudez, C.; Pohnert, G. Comparative metabolomics of the diatom Skeletonema marinoi in different growth phases. Metabolomics 2012, 8, 654–669. [Google Scholar] [CrossRef]
- Sánchez-Saavedra, M.P.; Voltolina, D. Effect of photon fluence rates of white and blue-green light on growth efficiency and pigment content of three diatom species in batch cultures. Cienc. Mar. 2002, 28, 273–279. [Google Scholar]
- Brunet, C.; Chandrasekaran, R.; Barra, L.; Giovagnetti, V.; Corato, F.; Ruban, A.V. Spectral radiation dependent photoprotective mechanism in the diatom Pseudo-nitzschia multistriata. PLoS ONE 2014, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lepetit, B.; Goss, R.; Jakob, T.; Wilhelm, C. Molecular dynamics of the diatom thylakoid membrane under different light conditions. Photosynth. Res. 2012, 111, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Jungandreas, A.; Costa, B.S.; Jakob, T.; von Bergen, M.; Baumann, S.; Wilhelm, C. The acclimation of Phaeodactylum tricornutum to blue and red light does not influence the photosynthetic light reaction but strongly disturbs the carbon allocation pattern. PLoS ONE 2014, 9, 1–14. [Google Scholar]
- Veuger, B.; van Oevelen, D. Long-term pigment dynamics and diatom survival in dark sediment. Limnol. Oceanogr. 2011, 56, 1065–1074. [Google Scholar] [CrossRef]
- Järup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, R.; Niwa-Kubota, M.; Matsui, H.; Matsuda, Y. Characterization of iron-responsive promoters in the marine diatom Phaeodactylum tricornutum. Mar. Genom. 2014, 16, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; Laroche, J.; Maheswari, U.; Lommer, M.; Schauer, N.; Lopez, P.J.; Finazzi, G.; Fernie, A.R.; Bowler, C. Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation. Proc. Natl. Acad. Sci. USA 2008, 105, 10438–10443. [Google Scholar] [CrossRef] [PubMed]
- Sivaci, R.E.; Sivaci, A.; Eroglu, S. Changes in photosynthetic pigments and species diversity of epiphytic diatoms on Myriophyllum triphyllum exposed to cadmium. ScienceAsia 2013, 39, 100–104. [Google Scholar] [CrossRef]
- Alexandre, M.; Gundermann, K.; Pascal, A.; van Grondelle, R.; Büchel, C.; Robert, B. Probing the carotenoid content of intact Cyclotella cells by resonance Raman spectroscopy. Photosynth. Res. 2014, 119, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Beer, A.; Juhas, M.; Büchel, C. Influence of different light intensities and different iron nutrition on the photosynthetic apparatus in the diatom Cyclotella meneghiniana (bacillariophyceae). J. Phycol. 2011, 47, 1266–1273. [Google Scholar] [CrossRef]
- Brotas, V.; Plante-Cuny, M.R. The use of HPLC pigment analysis to study microphytobenthos communities. Acta Oecol. 2003, 24, 109–115. [Google Scholar] [CrossRef]
- Costa, B.S.; Jungandreas, A.; Jakob, T.; Weisheit, W.; Mittag, M.; Wilhelm, C. Blue light is essential for high light acclimation and methylation and chromatin patterning photoprotection in the diatom Phaeodactylum tricornutum. J. Exp. Bot. 2013, 64, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Domingues, N.; Matos, A.R.; da Silva, J.M.; Cartaxana, P. Response of the Diatom Phaeodactylum tricornutum to photooxidative stress resulting from high light exposure. PLoS ONE 2012, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Murata, A.; Taguchi, S. Responses of pigment composition of the marine diatom Thalassiosira weissflogii to silicate availability during. Plankt. Benthos Res. 2011, 6, 1–11. [Google Scholar] [CrossRef]
- Lavaud, J. Fast regulation of photosynthesis in diatoms: Mechanisms, evolution and ecophysiology. Funct. Plant Sci. Biotechnol. 2007, 1, 267–287. [Google Scholar]
- Gibbs, S. The comparative ultrastructure of the algal chloroplast. Ann. N. Y. Acad. Sci. 1970, 175, 454–473. [Google Scholar] [CrossRef]
- Pyszniak, A.; Gibbs, S. Immunocytochemical localization of photosystem I and the fucoxanthinchlorophyll a/c light-harvesting complex in the diatom Phaeodactylum tricornutum. Protoplasma 1992, 166, 208–217. [Google Scholar] [CrossRef]
- Gundermann, K.; Büchel, C. The Structural Basis of Biological Energy Generation, 1st ed.; Hohmann-Marriott, M., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Papagiannakis, E.; van Stokkum, I.H.M.; Fey, H.; Büchel, C.; van Grondelle, R. Spectroscopic characterization of the excitation energy transfer in the fucoxanthinchlorophyll protein of diatoms. Photosynth. Res. 2005, 86, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Premvardhan, L.; Sandberg, D.J.; Fey, H.; Birge, R.R.; Büchel, C.; van Grondelle, R. The charge-transfer properties of the S2 state of fucoxanthin in solution and in fucoxanthin chlorophyll-a/c2 protein (FCP) based on stark spectroscopy and molecular-orbital theory. J. Phys. Chem. B 2008, 112, 11838–11853. [Google Scholar] [CrossRef] [PubMed]
- Premvardhan, L.; Bordes, L.; Beer, A.; Büchel, C.; Robert, B. Carotenoid structures and environments in trimeric and oligomeric fucoxanthin chlorophyll a/c2 proteins from resonance raman spectroscopy. J. Phys. Chem. B 2009, 113, 12565–12574. [Google Scholar] [CrossRef] [PubMed]
- Premvardhan, L.; Réfrégiers, M.; Büchel, C. Pigment organization effects on energy transfer and Chl a emission imaged in the diatoms C. meneghiniana and P. tricornutum in vivo: A confocal laser scanning fluorescence (CLSF) microscopy and spectroscopy study. J. Phys. Chem. B 2013, 117, 11272–11281. [Google Scholar] [CrossRef] [PubMed]
- Gundermann, K.; Schmidt, M.; Weisheit, W.; Mittag, M.; Büchel, C. Identification of several sub-populations in the pool of light harvesting proteins in the pennate diatom Phaeodactylum tricornutum. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Lepetit, B.; Volke, D.; Gilbert, M.; Wilhelm, C.; Goss, R. Evidence for the existence of one antenna-associated, lipid-dissolved and two protein-bound pools of diadinoxanthin cycle pigments in diatoms. Plant Physiol. 2010, 154, 1905–1920. [Google Scholar] [CrossRef] [PubMed]
- Eppard, M.; Rhiel, E. The genes encoding light-harvesting subunits of Cyclotella cryptica (Bacillariophyceae) constitute a complex and heterogeneous family. Mol. Gen. Genet. 1998, 260, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.; Chang, W. Crystal structure of spinach major light-harvesting complex at 2.72 A resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G. Biogeochemical Controls and Feedbacks on Ocean Primary Production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.J.; Peterson, C.G. Variation in benthic diatom (Bacillariophyceae) immigration with habitat characteristics and cell morphology. J. Phycol. 1989, 25, 120–129. [Google Scholar] [CrossRef]
- Padisák, J.; Soróczki-Pintér, É.; Rezner, Z. Sinking properties of some phytoplankton shapes and the relation of form resistance to morphological diversity of plankton—An experimental study. Hydrobiologia 2003, 500, 243–257. [Google Scholar] [CrossRef]
- Verleyen, E.; Hodgson, D.A.; Leavitt, P.R.; Sabbe, K.; Vyverman, W. Quantifying habitat-specific diatom production: A critical assessment using morphological and biogeochemical markers in Antarctic marine and lake sediments. Limnol. Oceanogr. 2004, 49, 1528–1539. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Knoll, A.H. Evolution of Primary Producers in the Sea. In Evolution of Primary Producers in the Sea; Falkowski, P., Knoll, A.H., Eds.; Academic Press: New York, NY, USA, 2007; p. 456. [Google Scholar]
- Haxo, F.T.; Blinks, L.R. Photosynthetic action spectra of marine algae. J. Gen. Physiol. 1950, 33, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Nicklisch, A. Growth and light absorption of some planktonic cyanobacteria, diatoms and Chlorophyceae under simulated natural light fluctuations. J. Plankton Res. 1998, 20, 105–119. [Google Scholar] [CrossRef]
- Spaulding, S.A.; Lubinski, D.J.; Potapova, M. Diatoms of the United States. Available online: http://westerndiatoms.colorado.edu (accessed on 6 July 2015).
- Johansen, J.R. Morphological variability and cell wall composition of Phaeodactylum tricornutum (Bacillariophyceae). Great Basin Nat. 1991, 51, 310–315. [Google Scholar]
- Wagner, A.H.; Jakob, T.; Wilhelm, C.; Wagner, H. Balancing the energy captured under fluctuating light conditions to. New Biotechnol. 2005, 169, 95–108. [Google Scholar]
- Holdsworth, E.S. Effect of growth factors and light quality on the growth, pigmentation and photosynthesis of two diatoms, Thalassiosira gravida and Phaeodactylum tricornutum. Mar. Biol. 1985, 86, 253–262. [Google Scholar] [CrossRef]
- Darley, W.; Volcani, B. Role of silicon in diatom metabolism. A silicon requirement for deoxyribonucleic acid synthesis in the diatom Cylindrotheca fusiformis Reimann and Lewin. Exp. Cell Res. 1969, 58, 334–342. [Google Scholar] [CrossRef]
- Lombardi, A.; Wangersky, P. Influence of phosphorus and silicon on lipia class production by the marine diatom Chaetoceros gracilis grown in turbidostat cage cultures. Mar. Ecol. Prog. Ser. 1991, 77, 39–47. [Google Scholar] [CrossRef]
- Brzezinski, M.A.; Baines, S.B.; Balch, W.M.; Beucher, C.P.; Chai, F.; Dugdale, R.C.; Krause, J.W.; Landry, M.R.; Marchi, A.; Measures, C.I.; et al. Co-limitation of diatoms by iron and silicic acid in the equatorial Pacific. Deep. Res. II Top. Stud. Oceanogr. 2011, 58, 493–511. [Google Scholar] [CrossRef]
- Lippemeier, S.; Hartig, P.; Colijn, F. Direct impact of silicate on the photosynthetic performance of the diatom Thalassiosira weissflogii assessed by on- and off-line PAM fluorescence measurements. J. Plankton Res. 1999, 21, 269–283. [Google Scholar] [CrossRef]
- Roberts, K.; Granum, E.; Leegood, R.C.; Raven, J.A. C3 and C4 pathways of photosynthetic carbon assimilation in marine diatoms are under genetic, not environmental, control. Plant Physiol. 2007, 145, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M.M. Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.; Lepetit, B. Biodiversity of NPQ. J. Plant Physiol. 2014, 172, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Lavaud, J.; Rousseau, B.; Etienne, A.L. In diatoms, a transthylakoid proton gradient alone is not sufficient to induce a non-photochemical fluorescence quenching. FEBS Lett. 2002, 523, 163–166. [Google Scholar] [CrossRef]
- Goss, R.; Ann Pinto, E.; Wilhelm, C.; Richter, M. The importance of a highly active and ΔpH-regulated diatoxanthin epoxidase for the regulation of the PS II antenna function in diadinoxanthin cycle containing algae. J. Plant Physiol. 2006, 163, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- García-Plazaola, J.I.; Matsubara, S.; Osmond, C.B. The lutein epoxide cycle in higher plants: Its relationships to other xanthophyll cycles and possible functions. Funct. Plant Biol. 2007, 34, 759–773. [Google Scholar] [CrossRef]
- Mewes, H.; Richter, M. Supplementary ultraviolet-B radiation induces a rapid reversal of the diadinoxanthin cycle in the strong light-exposed diatom Phaeodactylum tricornutum. Plant Physiol. 2002, 130, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Jakob, T.; Goss, R.; Wilhelm, C. Unusual pH-dependence of diadinoxanthin de-epoxidase activation causes chlororespiratory induced accumulation of diatoxanthin in the diatom Phaeodactylum tricornutum. J. Plant Physiol. 2001, 158, 383–390. [Google Scholar] [CrossRef]
- Grouneva, I.; Jakob, T.; Wilhelm, C.; Goss, R. Influence of ascorbate and pH on the activity of the diatom xanthophyll cycle-enzyme diadinoxanthin de-epoxidase. Physiol. Plant. 2006, 126, 205–211. [Google Scholar] [CrossRef]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef] [PubMed]
- Schumann, A.; Goss, R.; Jakob, T.; Wilhelm, C. Investigation of the quenching efficiency of diatoxanthin in cells of Phaeodactylum tricornutum (Bacillariophyceae) with different pool sizes of xanthophyll cycle pigments. Phycologia 2007, 46, 113–117. [Google Scholar] [CrossRef]
- Pinckney, M.; Micheli, F. Microalgae on seagrass mimics: Does epiphyte community structure differ from live seagrasses. J. Exp. Mar. Biol. Ecol. 1998, 221, 59–70. [Google Scholar] [CrossRef]
- Schagerl, M.; Künzl, G. Chlorophyll a extraction from freshwater algae—A reevaluation. Biol. Bratisl 2007, 62, 270–275. [Google Scholar] [CrossRef]
- Strickland, J.; Parsons, T. A Practical Handbook of Seawater Analysis, 2nd ed.; Stevenson, J.C., Ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; Volume 167. [Google Scholar]
- Holm-Hansen, O.; Lorenzen, C.; Holmes, R.; Strickland, J. Fluorometric determination of chlorophyll. J. Cons. Perm. Int. Explor. Mer 1965, 30, 3–15. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C. Evaluation of methods and solvents for pigment extraction. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Wright, S.W., Jeffrey, S.W., Mantoura, R.F.C., Eds.; UNESCO: Paris, France, 1997; pp. 261–282. [Google Scholar]
- Chen, N.; Bianchi, T.; Bland, J. Novel decomposition products of chlorophyll-a in continental shelf (Louisiana shelf) sediments: Formation and transformation of carotenol chlorine esters. Geochim. Cosmochim. Acta 2003, 67, 2027–2042. [Google Scholar] [CrossRef]
- Jeffrey, S. Profiles of photosynthetic pigments in the ocean using thin-layer chromatography. Mar. Biol. 1974, 26, 101–110. [Google Scholar] [CrossRef]
- Mantoura, R.; Llewellyn, C. The rapid determination of algal chlorophyll and carotenoid pigments and their breakdown products in natural waters by reverse-phase high-performance liquid chromatography. Anal. Chim. Acta 1983, 151, 297–314. [Google Scholar] [CrossRef]
- Bowles, N.; Paerl, H.; Tucker, J. Effective solvents and extraction periods employed in phytoplankton carotenoi d and chlorophyll determination. Can. J. Fish. Aquat. Sci. 1985, 42, 1127–1131. [Google Scholar] [CrossRef]
- Shoaf, W.T.; Lium, B.W. Improved extraction of chlorophyll a and b from algae using dimethyl sulfoxide. Limnol. Oceanogr. 1976, 21, 926–928. [Google Scholar] [CrossRef]
- Stock, R.; Rice, C.B.F. Chromatographic Methods, 1st ed.; Springer US: New York, NY, USA, 1974. [Google Scholar]
- Buffan-Dubau, E.; Carman, K.R. Extraction of benthic microalgal pigments for HPLC analyses. Mar. Ecol. Prog. Ser. 2000, 204, 293–297. [Google Scholar] [CrossRef]
- Jeffrey, S.; Hallegraeff, G. Chlorophyllase distribution in ten classes of phytoplankton: A problem for chlorophyll analysis. Mar. Ecol. Prog. Ser. 1987, 35, 293–304. [Google Scholar] [CrossRef]
- Barrett, J.; Jeffrey, S.W. Chlorophyllase and formation of an atypical chlorophyllide in marine algae. Plant Physiol. 1964, 39, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Draziewicz, M. Chlorophyllase: Occurrence, functions, mechanisms of action, effects of external and internal factor. Photosynthesis 1994, 30, 321–331. [Google Scholar]
- Rowan, K. Photosynthetic Pigments of Algae; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Jeffrey, S.W.; Mantoura, R.F.C.; Wright, S.W. Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S., Mantoura, R., Wright, S., Eds.; UNESCO Publishing: Paris, France, 1997. [Google Scholar]
- Jeffrey, S.; Wright, S.; Zapata, M. Recent advances in HPLC pigment analysis of phytoplankton. Mar. Freshw. Res. 1999, 50, 879–896. [Google Scholar] [CrossRef]
- Wright, S.; Shearer, J. Rapid extraction and HPLC of chlorophylls and carotenoids from marine phytoplankton. J. Chrom. 1984, 294, 281–295. [Google Scholar] [CrossRef]
- Wright, S.W. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar] [CrossRef]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Thomas, C. Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Kraay, G.W.; Zapata, M.; Veldhuis, M.J.W. Separation of chlorophylls c1c2, and c3 of marine phytoplankton by reversed-phase-C18-high-performance liquid chromatography. J. Phycol. 1992, 28, 708–712. [Google Scholar] [CrossRef]
- Mendes, C.R.; Cartaxana, P.; Brotas, V. HPLC determination of phytoplankton and microphytobenthos pigments: Comparing resolution and sensitivity of a C18 and a C8 method. Limnol. Oceanogr. Methods 2007, 5, 363–370. [Google Scholar] [CrossRef]
- Jeffrey, S.; Mantoura, R.; Bjørnland, T. Data for the identification of 47 key phytoplankton pigments. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S., Mantour, R., Wright, S., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 447–559. [Google Scholar]
- Jeffrey, S.W. Preparation and some properties of crystalline chlorophyll c1 and c2 from marine algae. Biochim. Biophys. Acta 1972, 279, 15–33. [Google Scholar] [CrossRef]
- Jensen, A. Algal carotenoids, I. Fucoxanthin monoacetate. Acta Chem. Scand. 1961, 15, 1604–1628. [Google Scholar] [CrossRef]
- Antia, N.J. The optical activity of fucoxanthin. Can. J. Chem. 1965, 43, 302–303. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Improved isolation procedure for fucoxanthin. Phytochemistry 1989, 28, 2797–2798. [Google Scholar] [CrossRef]
- Johansen, J.E.; Svec, W.A.; Liaaen-Jensen, S. Carotenoids of the Dinophyceae. Phytochemistry 1974, 13, 2261–2271. [Google Scholar] [CrossRef]
- Hager, A.; Meyer-Bertenrath, T. Die Isolierung und quantitative Bestimmung der Carotinoide und Chlorophylle von Blättern, Algen und isolierten Chloroplasten mit Hilfe dünnschichtchromatographischer Methoden. Planta 1966, 69, 198–217. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.H. Carotenoids. In Chemistry and Biochemistry of Plant Pigments Vol. II; Goodwin, T., Ed.; Academic Press: London, UK, 1976; pp. 38–165. [Google Scholar]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar]
- Isler, O.; Lindlar, H.; Montavon, M.; Rüegg, R.; Zeller, P. Die technische Synthese von beta-Carotin. Helv. Chim. Acta 1956, 39, 249–259. [Google Scholar] [CrossRef]
- Hiyama, T.; Nishimura, M.; Chance, B. Determination of carotenes by thin-layer chromatography. Anal. Biochem. 1969, 29, 339–342. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Palozza, P.; Krinsky, N.I. β-Carotene and α-tocopherol are synergistic antioxidants. Arch. Biochem. Biophys. 1992, 297, 184–187. [Google Scholar] [CrossRef]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy trial. J. Natl. Cancer Inst. 1997, 89, 325–326. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Landrum, J.T.; Bone, R.A. Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu. Rev. Nutr. 2003, 23, 171–201. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, V.; Morisset, P.; Ihammouine, S.; Chepied, A.; Aumailley, L.; Berard, J.B.; Serive, B.; Kaas, R.; Lanneluc, I.; Thiery, V.; et al. Antiproliferative activity of violaxanthin isolated from bioguided fractionation of Dunaliella tertiolecta extracts. Mar. Drugs 2011, 9, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-J.; Ko, S.-C.; Kang, S.-M.; Kang, H.-S.; Kim, J.-P.; Kim, S.-H.; Lee, K.-W.; Cho, M.-G.; Jeon, Y.-J. Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage. Eur. Food Res. Technol. 2008, 228, 145–151. [Google Scholar] [CrossRef]
- Kim, K.-N.; Heo, S.-J.; Yoon, W.-J.; Kang, S.-M.; Ahn, G.; Yi, T.-H.; Jeon, Y.-J. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Wanezaki, S.; Miyauchi, K.; Kurihara, H.; Kohno, H.; Kawabata, J.; Odashima, S.; Takahashi, K. Apoptosis-Inducing Effect of Fucoxanthin on Human Leukemia Cell Line HL-60. Food Sci. Technol. Res. 1999, 5, 243–246. [Google Scholar] [CrossRef]
- Asai, A.; Sugawara, T.; Ono, H.; Nagao, A. Biotransformation of fucoxanthinol into amarouciaxanthin a in mice and HepG2 cells: Formation and cytotoxicity of fucoxanthin metabolites. Drug Metab. Dispos. 2004, 32, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The effects of XanthigenTM in the weight management of obese premenopausal women with non-alcoholic fatty liver disease and normal liver fat. Diabetes Obes. Metab. 2010, 12, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Jaswir, I.; Dedi, N.; Salleh, H.M.; Taher, M.; Miyashita, K.; Ramli, N. Analysis of fucoxanthin content and purification of all-trans-fucoxanthin from Turbinaria turbinata and Sargassum plagyophyllum by SiO2 open column chromatography and reversed phase HPLC. J. Liq. Chromatogr. 2013, 36, 1340–1354. [Google Scholar]
- Kim, S.M.; Shang, Y.F.; Um, B.H. A preparative method for isolation of fucoxanthin from Eisenia bicyclis by centrifugal partition chromatography. Phytochem. Anal. 2011, 22, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Rebolloso-Fuentes, M.M.; Navarro-Pérez, A.; García-Camacho, F.; Ramos-Miras, J.J.; Guil-Guerrero, J.L. Biomass nutrient profiles of the microalga Nannochloropsis. J. Agric. Food Chem. 2001, 49, 2966–2972. [Google Scholar] [CrossRef] [PubMed]
- Gómez-loredo, A.; Benavides, J.; Rito-palomares, M. Partition behavior of fucoxanthin in ethanol-potassium phosphate two-phase systems. J. Chem. Technol. Biotechnol. 2014, 89, 1637–1645. [Google Scholar] [CrossRef]
- Kim, S.; Kang, S.-W.; Kwon, O.-N.; Chung, D.; Pan, C.-H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic Pigments in Diatoms. Mar. Drugs 2015, 13, 5847-5881. https://doi.org/10.3390/md13095847
Kuczynska P, Jemiola-Rzeminska M, Strzalka K. Photosynthetic Pigments in Diatoms. Marine Drugs. 2015; 13(9):5847-5881. https://doi.org/10.3390/md13095847
Chicago/Turabian StyleKuczynska, Paulina, Malgorzata Jemiola-Rzeminska, and Kazimierz Strzalka. 2015. "Photosynthetic Pigments in Diatoms" Marine Drugs 13, no. 9: 5847-5881. https://doi.org/10.3390/md13095847
APA StyleKuczynska, P., Jemiola-Rzeminska, M., & Strzalka, K. (2015). Photosynthetic Pigments in Diatoms. Marine Drugs, 13(9), 5847-5881. https://doi.org/10.3390/md13095847