
Comparison of Protein Phosphatase Inhibition Assay with LC-MS/MS for Diagnosis of Microcystin Toxicosis in Veterinary Cases

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
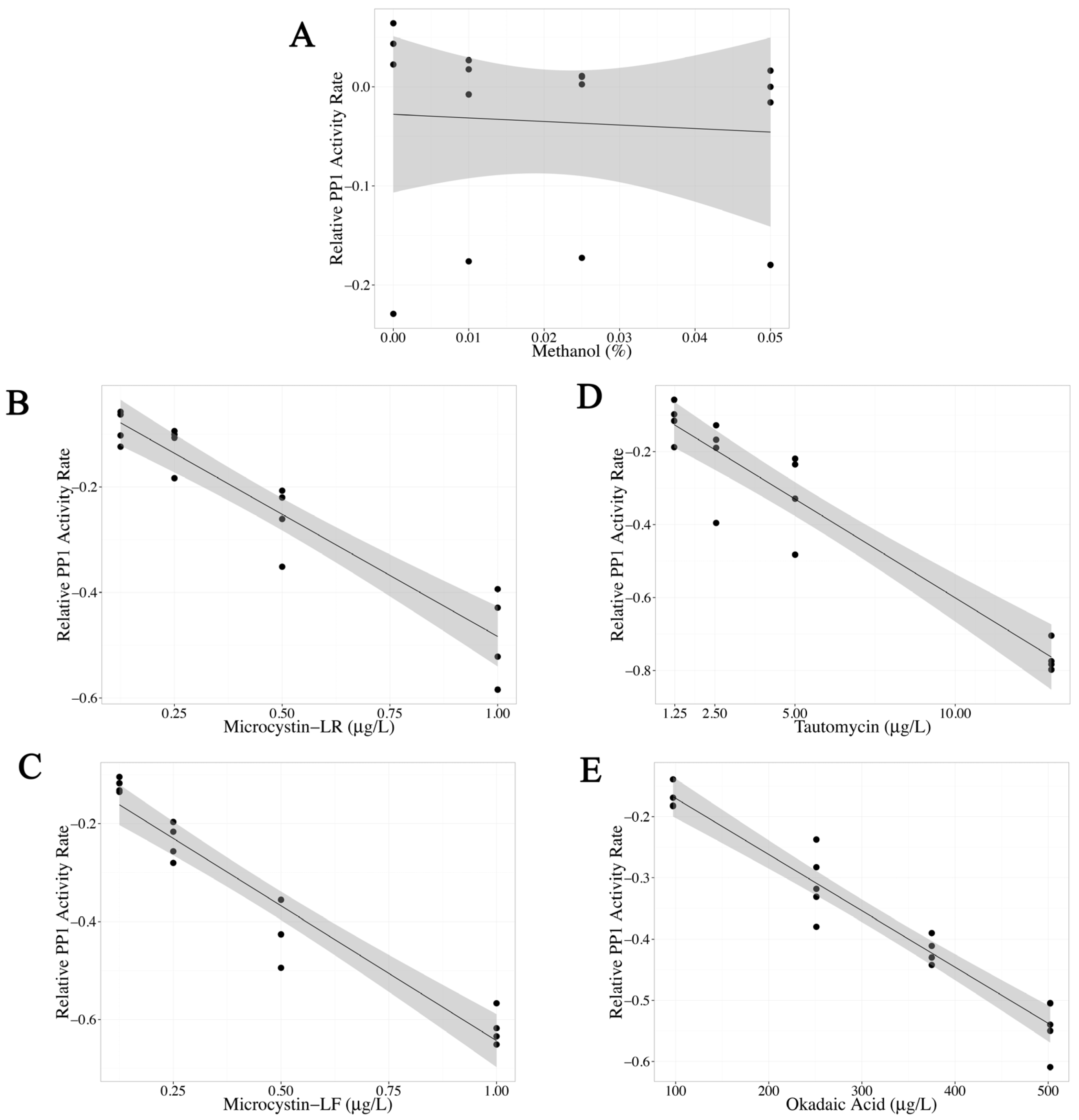
2.1. Statistical Evaluation of the PPIA with Environmental Samples and Presentation of Data
2.2. Optimization of the PPIA
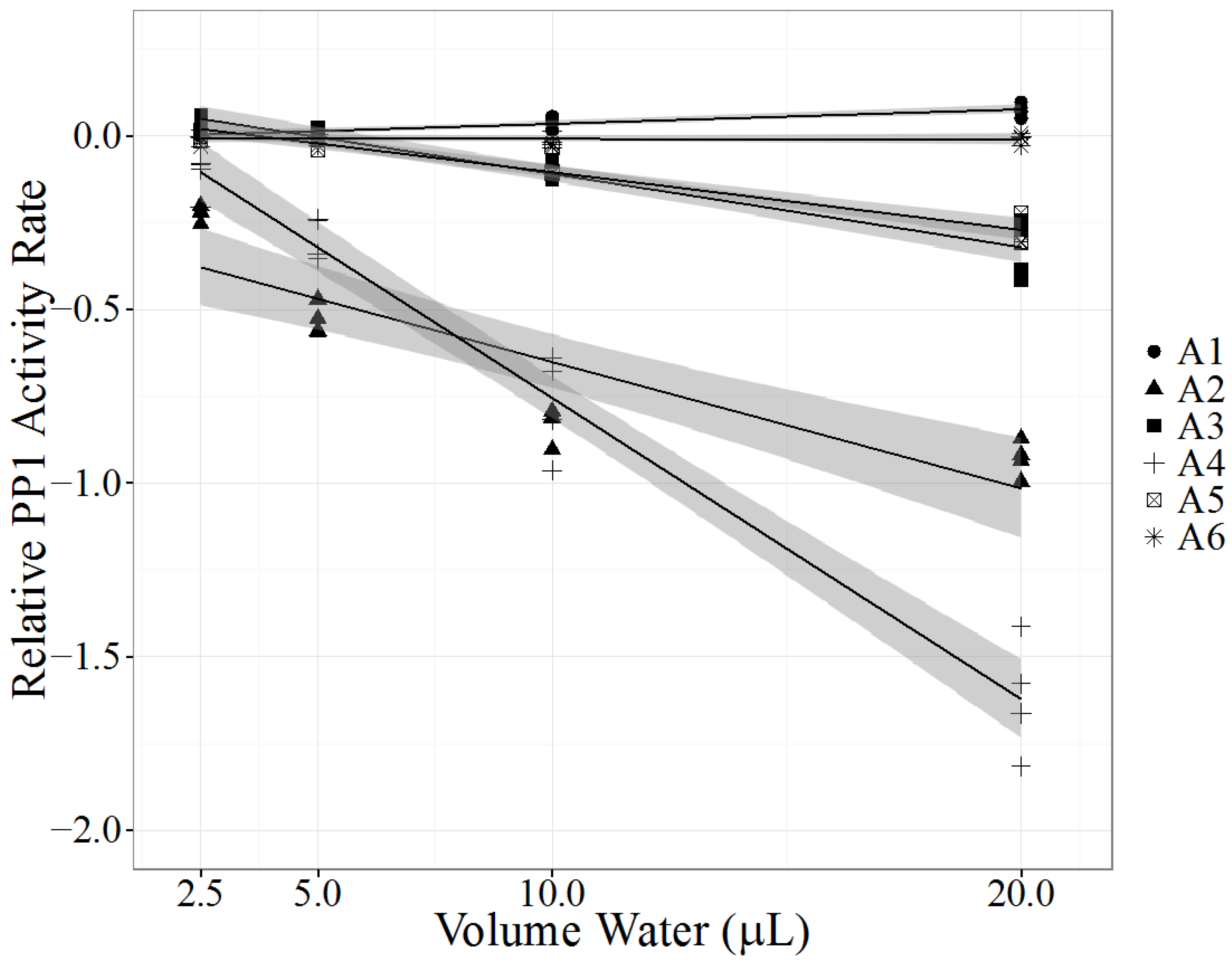
2.3. Case A: Kentucky Case
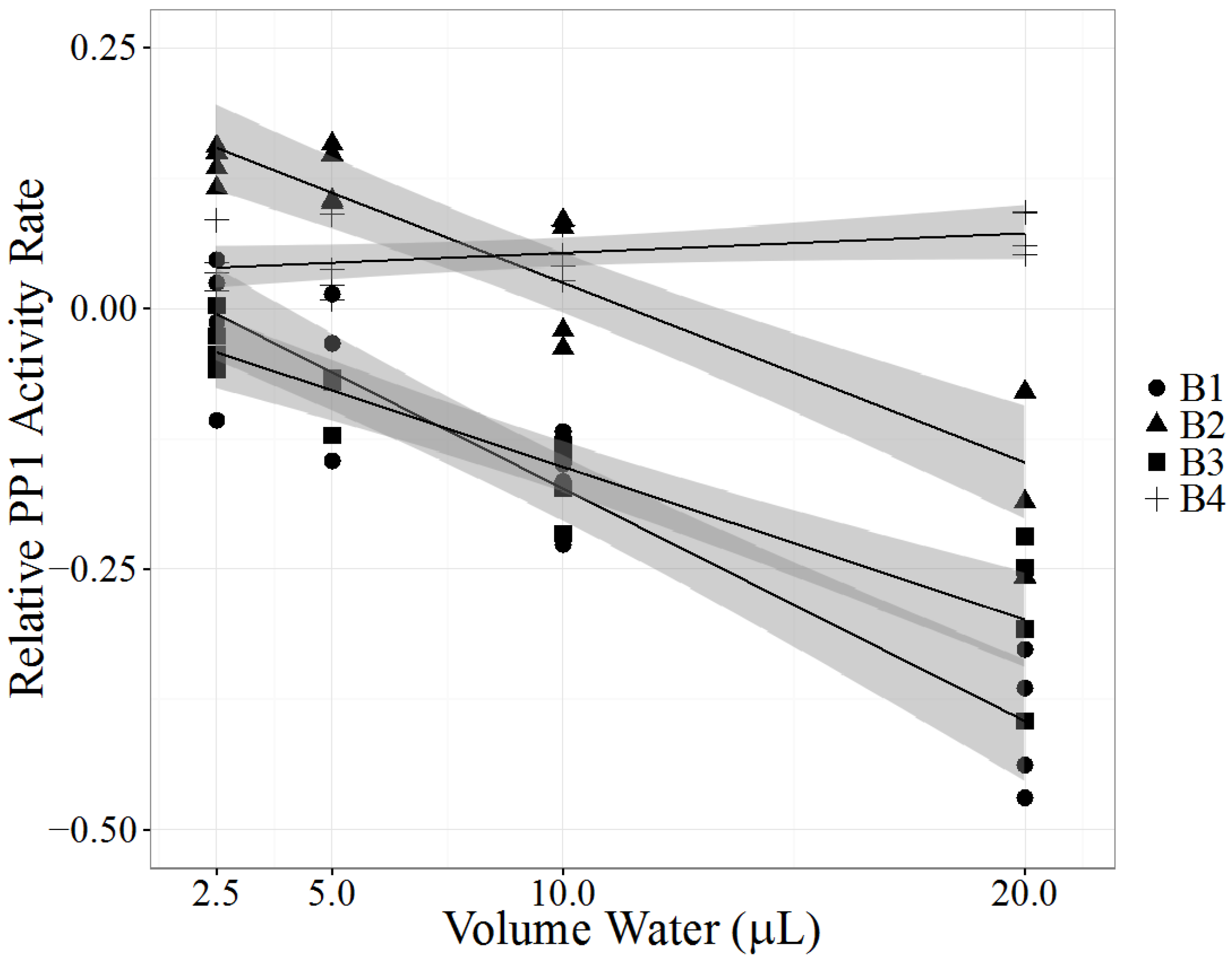
2.4. Case B: Clear Lake, California
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Sample Collection and Preparation
3.3. Liquid Chromatography Mass Spectrometry/Mass Spectrometry
3.4. Protein Phosphatase Inhibition Assay
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehr, J.D. Freshwater Algae of North America: Ecology and Classification; Academic Press: Waltham, MA, USA, 2002. [Google Scholar]
- Roegner, A.F.; Brena, B.; Gonzalez-Sapienza, G.; Puschner, B. Microcystins in potable surface waters: Toxic effects and removal strategies. J. Appl. Toxicol. 2014, 34, 441–457. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency Office of Water (4304T) Health and Ecological Criteria Division. Drinking Water Health Advisory for the Cyanobacterial Microcystin Toxins; EPA: Washington, DC, USA, 2015.
- Guidelines for Drinking-Water Quality. Addendum to Volume 1, Recommendations, 2nd ed.; World Health Organization: Geneva, Switzerland, 1998; p. 36.
- Fawell, J.K.; Mitchell, R.E.; Everett, D.J.; Hill, R.E. The toxicity of cyanobacterial toxins in the mouse: I microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, S.D.; Poppenga, R.H. Toxicosis due to microcystin hepatotoxins in three Holstein heifers. J. Vet. Diagn. Investig. 1993, 5, 651–653. [Google Scholar] [CrossRef]
- Kerr, L.A.; McCoy, C.P.; Eaves, D. Blue-green algae toxicosis in five dairy cows. J. Am. Vet. Med. Assoc. 1987, 191, 829–830. [Google Scholar] [PubMed]
- Moore, C.E.; Puschner, B. Chapter 44: Cyanobacteria. In Small Animal Toxicology, 3rd ed.; Saunders: Philadelphia, PA, USA, 2012. [Google Scholar]
- Lurling, M.; Faassen, E.J. Dog poisonings associated with a Microcystis aeruginosa bloom in the Netherlands. Toxins 2013, 5, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Sebbag, L.; Smee, N.; van der Merwe, D.; Schmid, D. Liver failure in a dog following suspected ingestion of blue-green algae (Microcystis spp.): A case report and review of the toxin. J. Am. Anim. Hosp. Assoc. 2013, 49, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, D.; Sebbag, L.; Nietfeld, J.C.; Aubel, M.T.; Foss, A.; Carney, E. Investigation of a Microcystis aeruginosa cyanobacterial freshwater harmful algal bloom associated with acute microcystin toxicosis in a dog. J. Vet. Diagn. Investig. 2012, 24, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.R.; Lund, J.C.; Schumacher, D.G.; Brakhage, P.A.; McManus, B.C.; Miller, J.D.; Augustine, M.M.; Carney, J.J.; Holland, R.S.; Hoagland, K.D.; et al. Chapter 6: Nebraska experience. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; Volume 619, pp. 139–152. [Google Scholar]
- Wood, S.A.; Heath, M.W.; Holland, P.T.; Munday, R.; McGregor, G.B.; Ryan, K.G. Identification of a benthic microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- DeVries, S.E.; Galey, F.D.; Namikoshi, M.; Woo, J.C. Clinical and pathologic findings of blue-green algae (Microcystis aeruginosa) intoxication in a dog. J. Vet. Diagn. Investig. 1993, 5, 403–408. [Google Scholar] [CrossRef]
- Craig, M.; Luu, H.A.; McCready, T.L.; Williams, D.; Andersen, R.J.; Holmes, C.F. Molecular mechanisms underlying he interaction of motuporin and microcystins with type-1 and type-2A protein phosphatases. Biochem. Cell Biol. 1996, 74, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, M.; Berndt, N.; Kong, S.M.; Lee, E.Y.; Zhang, L. In vivo and in vitro binding of microcystin to protein phosphatases 1 and 2A. Biochem. Biophys. Res. Commun. 1995, 216, 162–169. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- An, J.; Carmichael, W.W. Use of a colorimetric protein phosphatase inhibition assay and enzyme linked immunosorbent assay for the study of microcystins and nodularins. Toxicon 1994, 32, 1495–1507. [Google Scholar] [CrossRef]
- Ward, C.J.; Beattie, K.A.; Lee, E.Y.; Codd, G.A. Colorimetric protein phosphatase inhibition assay of laboratory strains and natural blooms of cyanobacteria: Comparisons with high-performance liquid chromatographic analysis for microcystins. FEMS Microbiol. Lett. 1997, 153, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, R.E.; Codispoti, B.A.; Tse, K.; Boynton, A.L.; Honkanan, R.E. Characterization of natural toxins with inhibitory activity against serine/threonine protein phosphatases. Toxicon 1994, 32, 339–350. [Google Scholar] [CrossRef]
- Honkanen, R.E.; Zwiller, J.; Moore, R.E.; Daily, S.L.; Khatra, B.S.; Dukelow, M.; Boynton, A.L. Characterization of microcystin-LR, a potent inhibitor of type 1 and type 2A protein phosphatases. J. Biol. Chem. 1990, 265, 19401–19404. [Google Scholar] [PubMed]
- Ash, C.; Mackintosh, C.; Mackintosh, R. Development of a Colorimetric Protein-Phosphorylation Assay for Detecting Cyanobacterial Toxins. Water Sci. Technol. 1995, 31, 47–49. [Google Scholar] [CrossRef]
- Bouaicha, N.; Maatouk, I.; Vincent, G.; Levi, Y. A colorimetric and fluorometric microplate assay for the detection of microcystin-LR in drinking water without preconcentration. Food Chem. Toxicol. 2002, 40, 1677–1683. [Google Scholar] [CrossRef]
- Rios, V.; Moreno, I.; Prieto, A.I.; Soria-Diaz, M.E.; Frias, J.E.; Camean, A.M. Comparison of Microcystis aeruginosa (PCC7820 and PCC7806) growth and intracellular microcystins content determined by liquid chromatography-mass spectrometry, enzyme-linked immunosorbent assay anti-Adda and phosphatase bioassay. J. Water Health 2014, 12, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Feurstein, D.; Kleinteich, J.; Heussner, A.H.; Stemmer, K.; Dietrich, D.R. Investigation of microcystin congener-dependent uptake into primary murine neurons. Environ. Health Perspect. 2010, 118, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; An, J. Using an enzyme linked immunosorbent assay (ELISA) and a protein phosphatase inhibition assay (PPIA) for the detection of microcystins and nodularins. Nat. Toxins 1999, 7, 377–385. [Google Scholar] [CrossRef]
- Heresztyn, T.; Nicholson, B.C. Determination of cyanobacterial hepatotoxins directly in water using a protein phosphatase inhibition assay. Water Res. 2001, 35, 3049–3056. [Google Scholar] [CrossRef]
- Takai, A.; Sasaki, K.; Nagai, H.; Mieskes, G.; Isobe, M.; Isono, K.; Yasumoto, T. Inhibition of specific binding of okadaic acid to protein phosphatase 2A by microcystin-LR, calyculin-A and tautomycin: Method of analysis of interactions of tight-binding ligands with target protein. Biochem. J. 1995, 306, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Reguera, B.; Riobo, P.; Rodriguez, F.; Diaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ju, J.; Osada, H.; Shen, B. Utilization of the methoxymalonyl-acyl carrier protein biosynthesis locus for cloning of the tautomycin biosynthetic gene cluster from Streptomyces spiroverticillatus. J. Bacteriol. 2006, 188, 4148–4152. [Google Scholar] [CrossRef] [PubMed]
- Van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Chorus, E.I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring And Management; WHO: Geneva, Switzerland, 1999. [Google Scholar]
- Molinspiration Software. Available online: http://www.molinspiration.com/services/logp.html (accessed on 21 July 2015).
- Garibo, D.; Flores, C.; Ceto, X.; Prieto-Simon, B.; del Valle, M.; Caixach, J.; Diogene, J.; Campas, M. Inhibition equivalency factors for microcystin variants in recombinant and wild-type protein phosphatase 1 and 2A assays. Environ. Sci. Pollut. Res. Int. 2014, 21, 10652–10660. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Vesterkvist, P.S.; Misiorek, J.O.; Spoof, L.E.; Toivola, D.M.; Meriluoto, J.A. Comparative cellular toxicity of hydrophilic and hydrophobic microcystins on Caco-2 cells. Toxins 2012, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed]
- Monks, N.R.; Liu, S.; Xu, Y.; Yu, H.; Bendelow, A.S.; Moscow, J.A. Potent cytotoxicity of the phosphatase inhibitor microcystin LR and microcystin analogues in OATP1B1- and OATP1B3-expressing HeLa cells. Mol. Cancer Ther. 2007, 6, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Puschner, B.; Galey, F.D.; Johnson, B.; Dickie, C.W.; Vondy, M.; Francis, T.; Holstege, D.M. Blue-green algae toxicosis in cattle. J. Am. Vet. Med. Assoc. 1998, 213, 1605–1607. [Google Scholar] [PubMed]
- Galey, F.D.; Beasley, V.R.; Carmichael, W.W.; Kleppe, G.; Hooser, S.B.; Haschek, W.M. Blue-green algae (Microcystis aeruginosa) hepatotoxicosis in dairy cows. Am. J. Vet. Res. 1987, 48, 1415–1420. [Google Scholar] [PubMed]
- Cattle Die From Toxic Algae. Available online: http://beefmagazine.com/health/cattle-die-toxic-algae (accessed on 1 August 2015).
- Blue-Green Algae Also a Threat to People, Other Animals. Available online: http://beefmagazine.com/parasites/blue-green-algae-also-threat-people-other-animals (accessed on 2 August 2015).
- Beasley, V.R.; Dahlem, A.M.; Cook, W.O.; Valentine, W.M.; Lovell, R.A.; Hooser, S.B.; Harada, K.; Suzuki, M.; Carmichael, W.W. Diagnostic and clinically important aspects of cyanobacterial (blue-green algae) toxicoses. J. Vet. Diagn. Investig. 1989, 1, 359–365. [Google Scholar] [CrossRef]
- Mioni, C.E.; Kudela, R.M.; Baxa, D.V. Harmful Cyanobacteria Blooms and their toxins in Clear Lake and the Sacramento-San Joaquin Delta (California). In Surface Water Ambient Monitoring Program (10-058-150); Central Valley Regional Water Quality Control Board: Rancho Cordova, CA, USA, 2011. [Google Scholar]
- Craig, M.; McCready, T.L.; Luu, H.A.; Smillie, M.A.; Dubord, P.; Holmes, C.F. Identification and characterization of hydrophobic microcystins in Canadian freshwater cyanobacteria. Toxicon 1993, 31, 1541–1549. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; E & FN Spon: London, UK; New York, NY, USA, 1999. [Google Scholar]
- Anderson, G. Clear Lake Algae Suspected in Death of Dog. Available online: http://www.pressdemocrat.com/csp/mediapool/sites/PressDemocrat/News/story.csp?cid=2212492&sid=555&fid=181 (accessed on 2 August 2015).
- Clear Lake Water Quality Investigation; California Department of Water Resources: Sacramento, CA, USA, 1966.
- Puschner, B.; Hoff, B.; Tor, E.R. Diagnosis of anatoxin-a poisoning in dogs from North America. J. Vet. Diagn. Investig. 2008, 20, 89–92. [Google Scholar] [CrossRef]
- Hill, H. Dog Deaths in Humboldt and Dog Deaths in Humboldt and Mendocino County Water Mendocino County Water Bodies Possibly Related to Bodies Possibly Related to Cyanobacterial Toxicity Cyanobacterial Toxicity. Available online: http://www.swrcb.ca.gov/water_issues/programs/bluegreen_algae/docs/workgroup110805/harriethill.pdf (accessed on 2 August 2015).
- Backer, L.C.; Landsberg, J.H.; Miller, M.; Keel, K.; Taylor, T.K. Canine cyanotoxin poisonings in the United States (1920s–2012): Review of suspected and confirmed cases from three data sources. Toxins 2013, 5, 1597–1628. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, L. After Boy Sickened, Pets Killed, Minnesota Warns of Toxic Blue-Green Algae in Lakes. Available online: http://www.startribune.com/vacationers-should-avoid-contact-with-blue-green-algae-mpca-warns/311618101/ (accessed on 2 August 2015).
- McDonough, B. New Brighton Family Offers Warning after Puppy Dies from Blue-Green Algae Exposure. Available online: http://kstp.com/article/stories/s3825778.shtml (accessed on 2 August 2015).
- Cuff, D. Dogs Die from East Bay Lake Toxic Algae Blooming Caused by Drought. Available online: http://www.mercurynews.com/drought/ci_27433372/dogs-die-from -lake -chabot- toxic- algae-blooming (accessed on 2 August 2015).
- Costantini, A. Dog Dies from Blue-Green Algae Poisoning. Available online: http://www.kare11.com/story/news/local/2015/06/08/blue-green-algae-dog-death-douglas-county-heiskary-brutlag-poison-helpline/28705241/ (accessed on 2 August 2015).
- Moulton, K.; Mims, B. Dog Dies after Swimming in Utah Lake; Toxin Warning Issued. Available online: http://www.sltrib.com/info/staff/1681075–155/utah-algae-lake-department-health-green (accessed on 2 August 2015).
- Francis, G. Poisonous Australian Lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef]
- Su, X.; Xue, Q.; Steinman, A.D.; Zhao, Y.; Xie, L. Spatiotemporal Dynamics of Microcystin Variants and Relationships with Environmental Parameters in Lake Taihu, China. Toxins 2015, 7, 3224–3244. [Google Scholar] [CrossRef] [PubMed]
- Messina, I. Microcystin Detected in RAW LAKE Erie Water, Toledo Water Deemed Safe to Drink. Available online: http://www.toledoblade.com/local/2015/07/27/Microcystin-detected-in-raw-Lake-Erie-water-Toledo-water-deemed-safe-to-drink.html (accessed on 2 August 2015).
- Tor, E.R.; Holstege, D.M.; Galey, F.D. Determination of cholinesterase activity in brain and blood samples using a plate reader. J. AOAC Int. 1994, 77, 1308–1313. [Google Scholar] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | LRS | SE | p Value | PP1 Ki (µg/L) |
|---|---|---|---|---|
| Methanol | −0.359 | 1.29 | 0.785 | - |
| MC-LR | −0.463 | 0.0423 | <0.0001 | 0.04 |
| MC-LF | −0.550 | 0.0402 | <0.0001 | - |
| Tautomycin | −0.0542 | 0.00473 | <0.0001 | 0.33 |
| Okadaic Acid | −0.000919 | 0.0000582 | <0.0001 | 118 |
| PPIA | |||||
|---|---|---|---|---|---|
| Sample Type | LC-MS/MS (µg/L) | LRS | SE | p Value | |
| A1 | Pond Water | N.D. | 0.00419 | 0.000497 | <0.0001 |
| A2 | Pond Water | LR 5.7 | −0.0364 | 0.00536 | <0.0001 |
| A3 | Rumen Contents | LR 1.3 | −0.0211 | 0.00169 | <0.0001 |
| A4 | Rumen Contents | LR 4.3 | −0.0867 | 0.00420 | <0.0001 |
| A5 | Rumen Contents | LR 1 | −0.0166 | 0.00116 | <0.0001 |
| A6 | Rumen Contents, Control | N.D. | −0.000120 | 0.000629 | 0.851 |
| Sample Collection | PPIA | |||||
|---|---|---|---|---|---|---|
| Location | Week | LC-MS/MS (µg/L) | LRS | SE | p Value | |
| B1 | Near Shore | 1 | LA: 2.1 | −0.0223 | 0.00216 | <0.0001 |
| LR: 0.4 | ||||||
| B2 | Near Shore | 4 | LA: 1.7 | −0.0173 | 0.00201 | <0.0001 |
| LR: 0.6 | ||||||
| LF: 50 | ||||||
| RR: 0.5 | ||||||
| B3 | 200 ft from shore | 4 | LR: 0.6 | −0.0147 | 0.00168 | <0.0001 |
| LF: 11 | ||||||
| RR: 0.2 | ||||||
| B4 | Near Shore | 5 | LR: 0.3 | 0.00188 | 0.000971 | 0.0727 |
| LF: 2.6 | ||||||
| RR: 0.20 | ||||||
| ID | Molecular Weight (Da) | LOD/LOQ (µg/L) | Retention Time (min) | Precursor Ion | Product Ion | Collision Energy |
|---|---|---|---|---|---|---|
| MC-RR | 1038.2 | 0.2/0.6 | 3.19 | 519.7 | 135.0 | 34 |
| MC-YR | 1045.2 | 0.3/0.7 | 3.95 | 523.6 | 135.6 | 30 |
| 523.6 | 507.8 | 30 | ||||
| 523.6 | 456.7 | 30 | ||||
| MC-LR | 995.2 | 0.2/0.5 | 4.30 | 498.4 | 135.1 | 20 |
| 498.4 | 430.5 | 20 | ||||
| 498.4 | 482.1 | 10 | ||||
| MC-LA | 910.0 | 1.3/4.2 | 6.70 | 910.0 | 372.8 | 30 |
| 910.0 | 399.5 | 30 | ||||
| 455.5 | 172.4 | 60 | ||||
| MC-LF | 986.2 | 1.9/6.4 | 7.63 | 1008.0 | 510.5 | 74 |
| 1008.0 | 504.2 | 73 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, C.E.; Juan, J.; Lin, Y.; Gaskill, C.L.; Puschner, B. Comparison of Protein Phosphatase Inhibition Assay with LC-MS/MS for Diagnosis of Microcystin Toxicosis in Veterinary Cases. Mar. Drugs 2016, 14, 54. https://doi.org/10.3390/md14030054
Moore CE, Juan J, Lin Y, Gaskill CL, Puschner B. Comparison of Protein Phosphatase Inhibition Assay with LC-MS/MS for Diagnosis of Microcystin Toxicosis in Veterinary Cases. Marine Drugs. 2016; 14(3):54. https://doi.org/10.3390/md14030054
Chicago/Turabian StyleMoore, Caroline E., Jeanette Juan, Yanping Lin, Cynthia L. Gaskill, and Birgit Puschner. 2016. "Comparison of Protein Phosphatase Inhibition Assay with LC-MS/MS for Diagnosis of Microcystin Toxicosis in Veterinary Cases" Marine Drugs 14, no. 3: 54. https://doi.org/10.3390/md14030054
APA StyleMoore, C. E., Juan, J., Lin, Y., Gaskill, C. L., & Puschner, B. (2016). Comparison of Protein Phosphatase Inhibition Assay with LC-MS/MS for Diagnosis of Microcystin Toxicosis in Veterinary Cases. Marine Drugs, 14(3), 54. https://doi.org/10.3390/md14030054