
Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Organization and Function of the pfa Genes
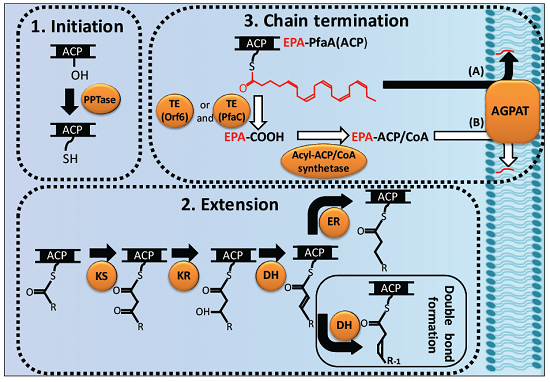
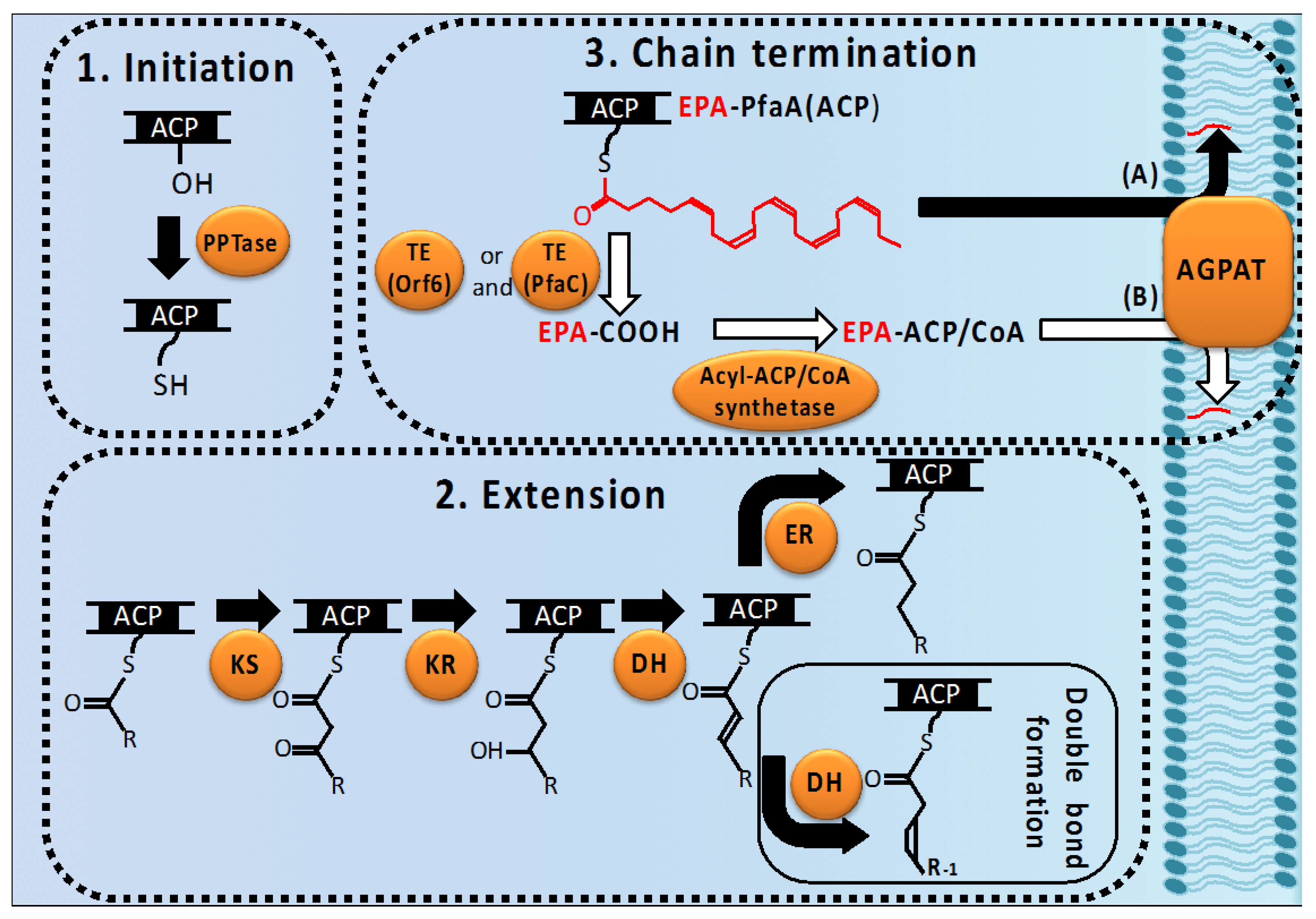
2.1. Cluster Structure and Domain Organization of the pfa Genes
2.2. Biosynthetic Process of LC-PUFAs in Bacteria
2.2.1. Initiation
2.2.2. Extension of Acyl Chains (Elongation, Double Bond Formation, and Final Product Determination)
2.2.3. Chain-Termination of LC-PUFA Biosynthesis and Their Integration into Lipids
2.3. Tertiary Structures of the Domains
3. Functions of LC-PUFAs in Bacteria and Their Practical Use
3.1. Involvement of LC-PUFAs in Cold Adaptation
3.2. Antioxidative Functions of LC-PUFAs
3.3. Involvement of LC-PUFAs in Specific Membrane Functions
3.4. Commercial Production and Use of LC-PUFAs
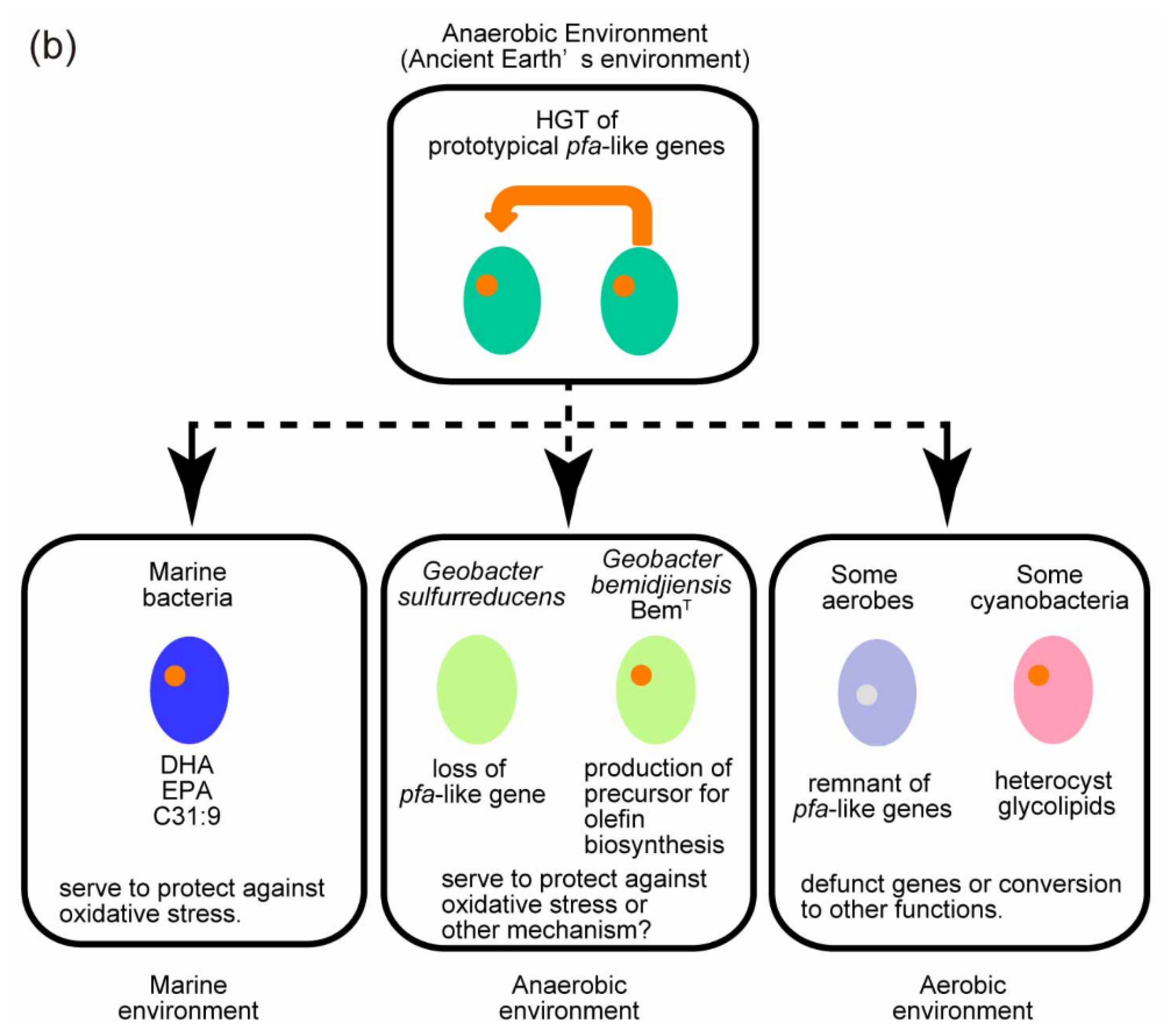
4. Evolution of C31:9 and LC-PUFA-Producing Bacteria
4.1. Possible Role of LC-HCs in Anaerobic Environments
4.2. Evolution of pfa-Like Genes
5. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Okuyama, H.; Orikasa, Y.; Nishida, T.; Watanabe, K.; Morita, N. Bacterial genes responsible for the biosynthesis of eicosapentaenoic and docosahexaenoic acids and their heterologous expression. Appl. Environ. Microbiol. 2007, 73, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, H.; Gu, Z.; Zhang, H.; Chen, W.; Chen, Y.Q. ω3 Fatty acid desaturases from microorganisms: Structure, function, evolution, and biotechnological use. Appl. Microbiol. Biotechnol. 2013, 97, 10225–10262. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabal, K.; Domergue, F.; Yamada, A.; et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.E.; Bartlett, D.H. Structure and regulation of the omega-3 polyunsaturated fatty acid synthase genes from the deep-sea bacterium Photobacterium profundum strain SS9. Microbiology 2002, 148, 1903–1913. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Tsai, S.-C. The type I fatty acid and polyketide synthases: A tale of two megasynthases. Nat. Prod. Rep. 2007, 24, 1041–1072. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48, 4688–4716. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, T.; Takata, N.; Kudo, T.; Horikoshi, K. Characteristic presence of polyunsaturated fatty acids in marine psychrophilic vibrios. FEMS Microbiol. Lett. 1995, 129, 51–56. [Google Scholar] [CrossRef]
- Niki, E. Free Radicals in Chemistry and Biochemistry. In Food and Free Radicals; Hiramatsu, M., Yoshikawa, T., Inoue, M., Eds.; Springer: New York, NY, USA, 1997; pp. 1–10. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1999; Volume 3. [Google Scholar]
- Nishida, T.; Orikasa, Y.; Ito, Y.; Yu, R.; Yamada, A.; Watanabe, K.; Okuyama, H. Escherichia coli engineered to produce eicosapentaenoic acid becomes resistant against oxidative damages. FEBS Lett. 2006, 580, 2731–2735. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D. A new n-C31:9 polyene hydrocarbon from Antarctic bacteria. FEMS Microbiol. Lett. 1995, 125, 281–285. [Google Scholar] [CrossRef]
- Sugihara, S.; Hori, R.; Nakanowatari, H.; Takada, Y.; Yumoto, I.; Morita, N.; Yano, Y.; Watanabe, K.; Okuyama, H. Possible biosynthetic pathways for all cis-3,6,9,12,15,19,22,25,28-hentriacontanonaene in bacteria. Lipids 2010, 45, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Sukovich, D.J.; Seffernick, J.L.; Richman, J.E.; Hunt, K.A.; Gralnick, J.A.; Wackett, L.P. Structure, function, and insights into the biosynthesis of a head-to-head hydrocarbon in Shewanella oneidensis strain MR-1. Appl. Environ. Microbiol. 2010, 76, 3842–3849. [Google Scholar] [CrossRef] [PubMed]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Oyola-Robles, D.; Gay, D.C.; Trujillo, U.; Sanchez-Pares, J.M.; Bermudez, M.L.; Rivera-Diaz, M.; Carballeira, N.M.; Baerga-Ortiz, A. Identification of novel protein domains required for the expression of an active dehydratase fragment from a polyunsaturated fatty acid synthase. Protein Sci. 2013, 22, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Shulse, C.N.; Allen, E.E. Widespread occurrence of secondary lipid biosynthesis potential in microbial lineages. PLoS ONE 2011, 6, e20146. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Jeong, Y.-S.; Kim, D.-K.; Seo, J.-W.; Hur, B.-K. Eicosapentaenoic acid (EPA) biosynthetic gene cluster of Shewanella oneidensis MR-1: Cloning, heterologous expression, and effects of temperature and glucose on the production of EPA in Escherichia coli. Biotechnol. Bioprocess Eng. 2006, 11, 510–516. [Google Scholar] [CrossRef]
- Hosoya, S.; Arunpairojana, V.; Suwannachart, C.; Kanjana-Opas, A.; Yokota, A. Aureispira marina gen. nov., sp. nov., a gliding, arachidonic acid-containing bacterium isolated from the southern coastline of Thailand. Int. J. Syst. Evol. Microbiol. 2006, 56, 2931–2935. [Google Scholar] [CrossRef] [PubMed]
- Ujihara, T.; Nagano, M.; Wada, H.; Mitsuhashi, S. Identification of a novel type of polyunsaturated fatty acid synthase involved in arachidonic acid biosynthesis. FEBS Lett. 2014, 588, 4032–4036. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; McCammon, S.A.; Lewis, T.; Skerratt, J.H.; Brown, J.L.; Nichols, D.S.; McMeekin, T.A. Psychroflexus torquis gen. nov., sp. nov., a psychrophilic species from Antarctic sea ice, and reclassification of Flavobacterium gondwanense (Dobson et al. 1993) as Psychroflexus gondwanense gen. nov., comb. nov. Microbiology 1998, 144, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Gemperlein, K.; Rachid, S.; Garcia, R.O.; Wenzel, S.C.; Müller, R. Polyunsaturated fatty acid biosynthesis in myxobacteria: Different PUFA synthases and their product diversity. Chem. Sci. 2014, 5, 1733–1741. [Google Scholar] [CrossRef]
- Orikasa, Y.; Nishida, T.; Hase, A.; Watanabe, K.; Morita, N.; Okuyama, H. A phosphopantetheinyl transferase gene essential for biosynthesis of n-3 polyunsaturated fatty acids from Moritella marina strain MP-1. FEBS Lett. 2006, 580, 4423–4429. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Pistorius, D.; Stadler, M.; Müller, R. Fatty acid-related phylogeny of myxobacteria as an approach to discover polyunsaturated omega-3/6 fatty acids. J. Bacteriol. 2011, 193, 1930–1942. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.I.; Vogel, H.J. Current understanding of fatty acid biosynthesis and the acyl carrier protein. Biochem. J. 2010, 430, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, S.; Orikasa, Y.; Okuyama, H. The Escherichia coli highly expressed entD gene complements the pfaE deficiency in a pfa gene clone responsible for the biosynthesis of long-chain n-3 polyunsaturated fatty acids. FEMS Microbiol. Lett. 2010, 307, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Orikasa, Y.; Tanaka, M.; Sugihara, S.; Hori, R.; Nishida, T.; Ueno, A.; Morita, N.; Yano, Y.; Yamamoto, K.; Shibahara, A.; et al. pfaB products determine the molecular species produced in bacterial polyunsaturated fatty acid biosynthesis. FEMS Microbiol. Lett. 2009, 295, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-N.; Kasai, W.; Kawamoto, J.; Esaki, N.; Kurihara, T. Characterization of 1-acyl-sn-glycerol-3-phosphate acyltransferase from a polyunsaturated fatty acid-producing bacterium, Shewanella livingstonensis Ac10. Trace Nutr. Res. 2012, 29, 92–99. [Google Scholar]
- Rodríguez-Guilbe, M.; Oyola-Robles, D.; Schreiter, E.R.; Baerga-Ortiz, A.; Rodriguez-Guilbe, M.; Oyola-Robles, D.; Schreiter, E.R.; Baerga-Ortiz, A. Structure, activity, and substrate selectivity of the Orf6 thioesterase from Photobacterium profundum. J. Biol. Chem. 2013, 288, 10841–10848. [Google Scholar] [CrossRef] [PubMed]
- Orikasa, Y.; Yamada, A.; Yu, R.; Ito, Y.; Nishida, T.; Yumoto, I.; Watanabe, K.; Okuyama, H. Characterization of the eicosapentaenoic acid biosynthesis gene cluster from Shewanella sp. strain SCRC-2738. Cell. Mol. Biol. 2004, 50, 625–630. [Google Scholar] [PubMed]
- Oyola-Robles, D.; Rullan-Lind, C.; Carballeira, N.M.; Baerga-Ortiz, A. Expression of dehydratase domains from a polyunsaturated fatty acid synthase increases the production of fatty acids in Escherichia coli. Enzym. Microb. Technol. 2014, 55, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yang, J.; Xian, M.; Xu, X.; Liu, W. Increasing unsaturated fatty acid contents in Escherichia coli by coexpression of three different genes. Appl. Microbiol. Biotechnol. 2010, 87, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.C.; Bateman, A. The Hotdog fold: Wrapping up a superfamily of thioesterases and dehydratases. BMC Bioinform. 2004, 5, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, U.; Vazquez-Rosa, E.; Oyola-Robles, D.; Stagg, L.J.; Vassallo, D.A.; Vega, I.E.; Arold, S.T.; Baerga-Ortiz, A. Solution structure of the tandem acyl carrier protein domains from a polyunsaturated fatty acid synthase reveals beads-on-a-string configuration. PLoS ONE 2013, 8, e57859. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zirkle, R.; Metz, J.G.; Braun, L.; Richter, L.; van Lanen, S.G.; Shen, B. The role of tandem acyl carrier protein domains in polyunsaturated fatty acid biosynthesis. J. Am. Chem. Soc. 2008, 130, 6336–6337. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Eisman, E.B.; Dutta, S.; Franzmann, T.M.; Walter, S.; Gerwick, W.H.; Skiniotis, G.; Sherman, D.H. Tandem acyl carrier proteins in the curacin biosynthetic pathway promote consecutive multienzyme reactions with a synergistic effect. Angew. Chem. Int. Ed. Engl. 2011, 50, 2795–2798. [Google Scholar] [CrossRef] [PubMed]
- Mertens, H.D.T.; Svergun, D.I. Structural characterization of proteins and complexes using small-angle X-ray solution scattering. J. Struct. Biol. 2010, 172, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Weissman, K.J. Uncovering the structures of modular polyketide synthases. Nat. Prod. Rep. 2015, 32, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Kato, C.; Nogi, Y. Correlation between phylogenetic structure and function: Examples from deep-sea Shewanella. FEMS Microbiol. Ecol. 2001, 35, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Valentine, R.C.; Valentine, D.L. Omega-3 fatty acids in cellular membranes: A unified concept. Prog. Lipid Res. 2004, 43, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.E.; Facciotti, D.; Bartlett, D.H. Monounsaturated but not polyunsaturated fatty acids are required for growth of the deep-sea bacterium Photobacterium profundum SS9 at high pressure and low temperature. Appl. Environ. Microbiol. 1999, 65, 1710–1720. [Google Scholar] [PubMed]
- Motoigi, T.; Okuyama, H.; Morita, N.; Hokkaido University, Sapporo, Japan. Unpublished work. 2013.
- Kawamoto, J.; Kurihara, T.; Yamamoto, K.; Nagayasu, M.; Tani, Y.; Mihara, H.; Hosokawa, M.; Baba, T.; Sato, S.B.; Esaki, N. Eicosapentaenoic acid plays a beneficial role in membrane organization and cell division of a cold-adapted bacterium, Shewanella livingstonensis Ac10. J. Bacteriol. 2009, 191, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Kato, C.; Horikoshi, K. Taxonomic studies of deep-sea barophilic Shewanella strains and description of Shewanella violacea sp. nov. Arch. Microbiol. 1998, 170, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Usui, K.; Hiraki, T.; Kawamoto, J.; Kurihara, T.; Nogi, Y.; Kato, C.; Abe, F. Eicosapentaenoic acid plays a role in stabilizing dynamic membrane structure in the deep-sea piezophile Shewanella violacea: A study employing high-pressure time-resolved fluorescence anisotropy measurement. Biochim. Biophys. Acta 2012, 1818, 574–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Wang, P.; Chen, M.; Xiao, X. Isolation of extremophiles with the detection and retrieval of Shewanella strains in deep-sea sediments from the west Pacific. Extremophiles 2004, 8, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xiao, X.; Ou, H.-Y.; Gai, Y.; Wang, F. Role and regulation of fatty acid biosynthesis in the response of Shewanella piezotolerans WP3 to different temperatures and pressures. J. Bacteriol. 2009, 191, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Lippmeier, J.C.; Crawford, K.S.; Owen, C.B.; Rivas, A.A.; Metz, J.G.; Apt, K.E. Characterization of both polyunsaturated fatty acid biosynthetic pathways in Schizochytrium sp. Lipids 2009, 44, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Semchyshyn, H.; Bagnyukova, T.; Storey, K.; Lushchak, V. Hydrogen peroxide increases the activities of soxRS regulon enzymes and the levels of oxidized proteins and lipids in Escherichia coli. Cell Biol. Int. 2005, 29, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Orikasa, Y.; Watanabe, K.; Okuyama, H. The cell membrane-shielding function of eicosapentaenoic acid for Escherichia coli against exogenously added hydrogen peroxide. FEBS Lett. 2006, 580, 6690–6694. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Morita, N.; Yano, Y.; Orikasa, Y.; Okuyama, H. The antioxidative function of eicosapentaenoic acid in a marine bacterium, Shewanella marinintestina IK-1. FEBS Lett. 2007, 581, 4212–4216. [Google Scholar] [CrossRef] [PubMed]
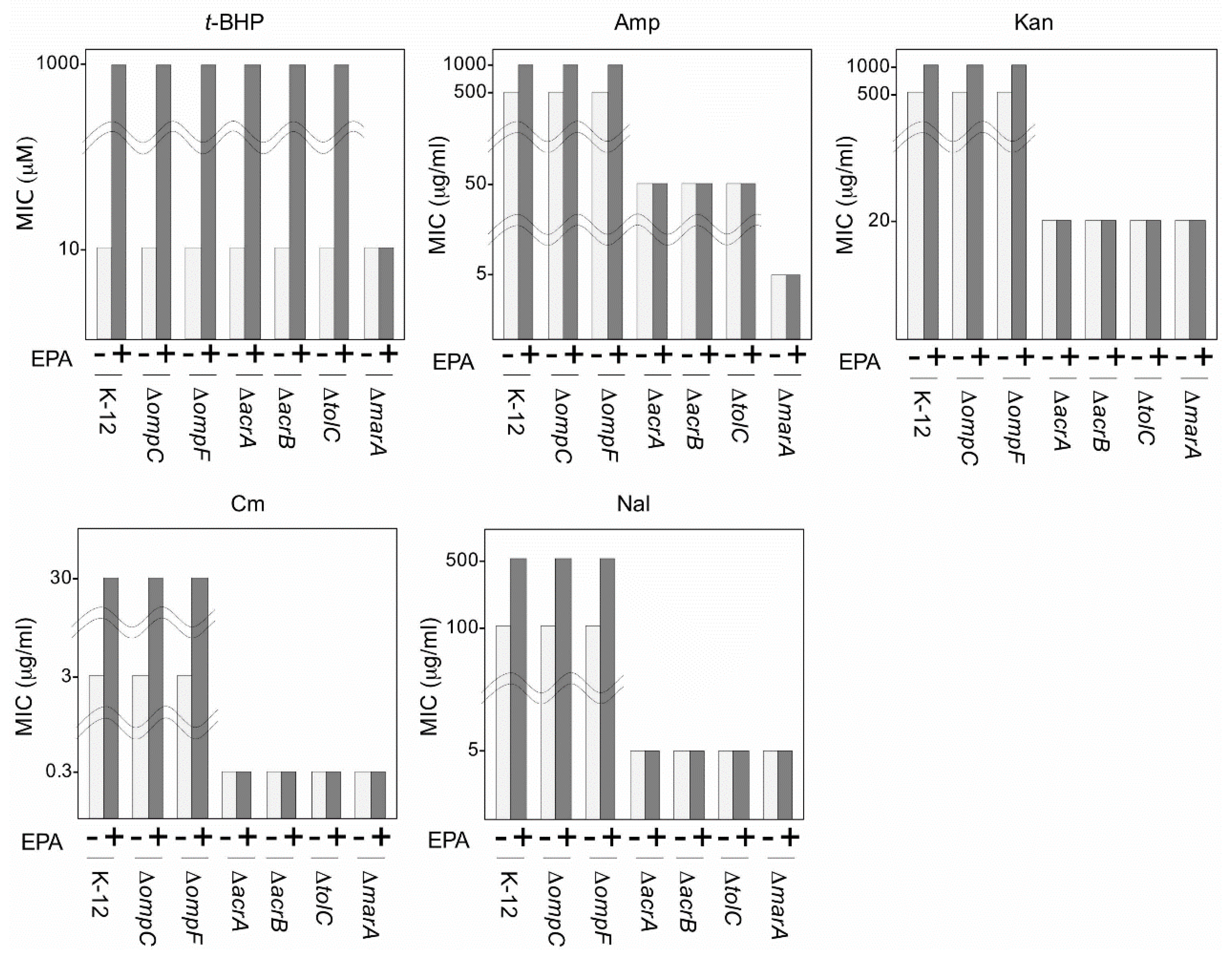
- Nishida, T.; Hori, R.; Morita, N.; Okuyama, H. Membrane eicosapentaenoic acid is involved in the hydrophobicity of bacterial cells and affects the entry of hydrophilic and hydrophobic compounds. FEMS Microbiol. Lett. 2010, 306, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, H.; Orikasa, Y.; Nishida, T. Significance of antioxidative functions of eicosapentaenoic and docosahexaenoic acids in marine microorganisms. Appl. Environ. Microbiol. 2008, 74, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Tilay, A.; Annapure, U. Novel simplified and rapid method for screening and isolation of polyunsaturated fatty acids producing marine bacteria. Biotechnol. Res. Int. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Rajamoorthi, K.; Petrache, H.I.; McIntosh, T.J.; Brown, M.F. Packing and viscoelasticity of polyunsaturated ω-3 and ω-6 lipid bilayers as seen by 2H NMR and X-ray diffraction. J. Am. Chem. Soc. 2005, 127, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- Hori, R.; Nishida, T.; Okuyama, H. Hydrophilic and hydrophobic compounds antithetically affect the growth of eicosapentaenoic acid-synthesizing Escherichia coli recombinants. Open Microbiol. J. 2011, 5, 114–118. [Google Scholar] [PubMed]
- Sato, S.; Kawamoto, J.; Sato, S.B.; Watanabe, B.; Hiratake, J.; Esaki, N.; Kurihara, T. Occurrence of a bacterial membrane microdomain at the cell division site enriched in phospholipids with polyunsaturated hydrocarbon chains. J. Biol. Chem. 2012, 287, 24113–24121. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.-Z.; Kawamoto, J.; Sato, S.B.; Esaki, N.; Kurihara, T. Eicosapentaenoic acid facilitates the folding of an outer membrane protein of the psychrotrophic bacterium, Shewanella livingstonensis Ac10. Biochem. Biophys. Res. Commun. 2012, 425, 363–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikaido, H.; Rosenberg, E.Y.; Foulds, J. Porin channels in Escherichia coli: Studies with beta-lactams in intact cells. J. Bacteriol. 1983, 153, 232–240. [Google Scholar] [PubMed]
- Tikhonova, E.B.; Zgurskaya, H.I. AcrA, AcrB, and TolC of Escherichia coli form a stable intermembrane multidrug efflux complex. J. Biol. Chem. 2004, 279, 32116–32124. [Google Scholar] [CrossRef] [PubMed]
- Hino, A.; Adachi, H.; Toyomasu, K.; Yoshida, N.; Enomoto, M.; Hiratsuka, A.; Hirai, Y.; Satoh, A.; Imaizumi, T. Very long chain N-3 fatty acids intake and carotid atherosclerosis: An epidemiological study evaluated by ultrasonography. Atherosclerosis 2004, 176, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Jicha, G.A.; Markesbery, W.R. Omega-3 fatty acids: Potential role in the management of early Alzheimer’s disease. Clin. Interv. Aging 2010, 5, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Kunisawa, J.; Arita, M.; Hayasaka, T.; Harada, T.; Iwamoto, R.; Nagasawa, R.; Shikata, S.; Nagatake, T.; Suzuki, H.; Hashimoto, E.; et al. Dietary ω3 fatty acid exerts anti-allergic effect through the conversion to 17,18-epoxyeicosatetraenoic acid in the gut. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, L.; Garssen, J.; Willemsen, L. Long chain N-3 polyunsaturated fatty acids in the prevention of allergic and cardiovascular disease. Curr. Pharm. Des. 2012, 18, 2375–2392. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.-J.; Zhou, J.-D.; Dong, J.-Y.; Ding, W.-Q.; Wu, J.-C. Dietary intake of n-3 fatty acids and colorectal cancer risk: A meta-analysis of data from 489,000 individuals. Br. J. Nutr. 2012, 108, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.-S.; Hu, X.-J.; Zhao, Y.-M.; Yang, J.; Li, D. Intake of fish and marine n-3 polyunsaturated fatty acids and risk of breast cancer: Meta-analysis of data from 21 independent prospective cohort studies. BMJ 2013, 346. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Martins, D.A.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Fujita, Y. Isolation of enhanced eicosapentaenoic acid producing mutants of Nannochloropsis oculata ST-6 using ethyl methane sulfonate induced mutagenesis techniques and their characterization at mRNA transcript level. Phycol. Res. 2006, 54, 208–219. [Google Scholar] [CrossRef]
- Meiser, A.; Schmid-Staiger, U.; Trösch, W. Optimization of eicosapentaenoic acid production by Phaeodactylum tricornutum in the flat panel airlift (FPA) reactor. J. Appl. Phycol. 2004, 16, 215–225. [Google Scholar] [CrossRef]
- Sakuradani, E.; Ando, A.; Shimizu, S.; Ogawa, J. Metabolic engineering for the production of polyunsaturated fatty acids by oleaginous fungus Mortierella alpina 1S-4. J. Biosci. Bioeng. 2013, 116, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, H.; Shimizu, S.; Shinmen, Y. Production of arachidonic acid by Mortierella elongata 1S-5. Agric. Biol. Chem. 1987, 51, 785–790. [Google Scholar] [CrossRef]
- Tavares, S.; Grotkjær, T.; Obsen, T.; Haslam, R.P.; Napier, J.A.; Gunnarsson, N. Metabolic engineering of Saccharomyces cerevisiae for production of eicosapentaenoic acid, using a novel ∆5-desaturase from Paramecium tetraurelia. Appl. Environ. Microbiol. 2011, 77, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Perveen, Z.; Ando, H.; Ueno, A.; Ito, Y.; Yamamoto, Y.; Yamada, Y.; Takagi, T.; Kaneko, T.; Kogame, K.; Okuyama, H. Isolation and characterization of a novel thraustochytrid-like microorganism that efficiently produces docosahexaenoic acid. Biotechnol. Lett. 2006, 28, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-J.; Ji, X.-J.; Huang, H.; Qu, L.; Feng, Y.; Tong, Q.-Q.; Ouyang, P.-K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Quilodrán, B.; Hinzpeter, I.; Hormazabal, E.; Quiroz, A.; Shene, C. Docosahexaenoic acid (C22:6n-3, DHA) and astaxanthin production by Thraustochytriidae sp. AS4-A1 a native strain with high similitude to Ulkenia sp.: Evaluation of liquid residues from food industry as nutrient sources. Enzym. Microb. Technol. 2010, 47, 24–30. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, F. Effects of medium glucose concentration and pH on docosahexaenoic acid content of heterotrophic Crypthecodinium cohnii. Process Biochem. 2000, 35, 1205–1209. [Google Scholar] [CrossRef]
- Luy, M.; Rusing, M.; Kiy, T. PUFA-PKS Gene from Ulkenia. US Patent 20090093033 A1, 9 April 2009. [Google Scholar]
- Liu, Y.; Tang, J.; Li, J.; Daroch, M.; Cheng, J.J. Efficient production of triacylglycerols rich in docosahexaenoic acid (DHA) by osmo-heterotrophic marine protists. Appl. Microbiol. Biotechnol. 2014, 98, 9643–9652. [Google Scholar] [CrossRef] [PubMed]
- Konagai, C.; Yanagimoto, K.; Hayamizu, K.; Han, L.; Tsuji, T.; Koga, Y. Effects of krill oil containing n-3 polyunsaturated fatty acids in phospholipid form on human brain function: A randomized controlled trial in healthy elderly volunteers. Clin. Interv. Aging 2013, 8, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Bunea, R.; El Farrah, K.; Deutsch, L. Evaluation of the effects of Neptune Krill oil on the clinical course of hyperlipidemia. Altern. Med. Rev. 2004, 9, 420–428. [Google Scholar] [PubMed]
- Yurko-Mauro, K.; Kralovec, J.; Bailey-Hall, E.; Smeberg, V.; Stark, J.G.; Salem, N. Similar eicosapentaenoic acid and docosahexaenoic acid plasma levels achieved with fish oil or krill oil in a randomized double-blind four-week bioavailability study. Lipids Health Dis. 2015, 14, 99. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, H.; Orikasa, Y.; Nishida, T. In vivo conversion of triacylglycerol to docosahexaenoic acid-containing phospholipids in a thraustochytrid-like microorganism, strain 12B. Biotechnol. Lett. 2007, 29, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Iskandar Bin Haji Mohd Taha; Sato, M.; Tojo, M.; Kanda, K.; Kimoto, T.; Kaneda, T.; Okuyama, H. Fermentative production of long-chain polyunsaturated fatty acid-containing phospholipids by the two-step cultivation method using microorganisms. New Food Ind. 2012, 54, 1–8. [Google Scholar]
- Stamey, J.A.; Shepherd, D.M.; de Veth, M.J.; Corl, B.A. Use of algae or algal oil rich in n-3 fatty acids as a feed supplement for dairy cattle. J. Dairy Sci. 2012, 95, 5269–5275. [Google Scholar] [CrossRef] [PubMed]
- Sardi, L.; Martelli, G.; Lambertini, L.; Parisini, P.; Mordenti, A. Effects of a dietary supplement of DHA-rich marine algae on Italian heavy pig production parameters. Livest. Sci. 2006, 103, 95–103. [Google Scholar] [CrossRef]
- Park, J.H.; Upadhaya, S.D.; Kim, I.H. Effect of dietary marine microalgae (Schizochytrium) powder on egg production, blood lipid profiles, egg quality, and fatty acid composition of egg yolk in layers. Asian-Australas. J. Anim. Sci. 2015, 28, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. How obligatory is anaerobiosis? Mol. Microbiol. 2008, 68, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Santos, W.G.D.; Pacheco, I.; Liu, M.; Teixeira, M.; Xavier, A.V.; LeGall, J. Purification and characterization of an iron superoxide dismutase and a catalase from the sulfate-reducing bacterium Desulfovibrio gigas. J. Bacteriol. 2000, 182, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Brioukhanov, A.L.; Thauer, R.K.; Netrusov, A.I. Catalase and superoxide dismutase in the cells of strictly anaerobic microorganisms. Microbiology 2002, 71, 281–285. [Google Scholar] [CrossRef]
- Jenney, F.E.; Verhagen, M.F.; Cui, X.; Adams, M.W. Anaerobic microbes: Oxygen detoxification without superoxide dismutase. Science 1999, 286, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Lombard, M.; Fontecave, M.; Touati, D.; Nivière, V. Reaction of the desulfoferodoxin from Desulfoarculus baarsii with superoxid anion. Evidence for a superoxide reductase activity. J. Biol. Chem. 2000, 275, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, L.B. Bacterial defenses against oxidants: Mechanistic features of cysteine-based peroxidases and their flavoprotein reductases. Arch. Biochem. Biophys. 2005, 433, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Wildschut, J.D.; Lang, R.M.; Voordouw, J.K.; Voordouw, G. Rubredoxin: Oxygen oxidoreductase enhances survival of Desulfovibrio vulgaris hildenborough under microaerophilic conditions. J. Bacteriol. 2006, 188, 6253–6260. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Coppi, M.V.; Lovley, D.R. Geobacter sulfurreducens can grow with oxygen as a terminal electron acceptor. Appl. Environ. Microbiol. 2004, 70, 2525–2528. [Google Scholar] [PubMed]
- Nunez, C.; Adams, L.; Childers, S.; Lovley, D.R. The RpoS sigma factor in the dissimilatory Fe(III)-reducing bacterium Geobacter sulfurreducens. J. Bacteriol. 2004, 186, 5543–5546. [Google Scholar] [CrossRef] [PubMed]
- Sukovich, D.J.; Seffernick, J.L.; Richman, J.E.; Gralnick, J.A.; Wackett, L.P. Widespread head-to-head hydrocarbon biosynthesis in bacteria and role of OleA. Appl. Environ. Microbiol. 2010, 76, 3850–3862. [Google Scholar] [CrossRef] [PubMed]
- Burdige, D.J. The biogeochemistry of manganese and iron reduction in marine sediments. Earth-Sci. Rev. 1993, 35, 249–284. [Google Scholar] [CrossRef]
- Lovley, D.R. Dissimilatory Fe(III) and Mn(IV) reduction. Microbiol. Rev. 1991, 55, 259–287. [Google Scholar] [PubMed]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Roden, E.E.; Sobolev, D.; Glazer, B.; Luther, G.W. Potential for microscale bacterial Fe redox cycling at the aerobic-anaerobic interface. Geomicrobiol. J. 2004, 21, 379–391. [Google Scholar] [CrossRef]
- Pham, A.N.; Waite, T.D. Oxygenation of Fe(II) in natural waters revisited: Kinetic modeling approaches, rate constant estimation and the importance of various reaction pathways. Geochim. Cosmochim. Acta 2008, 72, 3616–3630. [Google Scholar] [CrossRef]
- Millero, F.J.; Sotolongo, S.; Izaguirre, M. The oxidation kinetics of Fe(II) in seawater. Geochim. Cosmochim. Acta 1987, 51, 793–801. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Manganese inhibition of microbial iron reduction in anaerobic sediments. Geomicrobiol. J. 1988, 6, 145–155. [Google Scholar] [CrossRef]
- Myers, C.R.; Nealson, K.H. Microbial reduction of manganese oxides: Interactions with iron and sulfur. Geochim. Cosmochim. Acta 1988, 52, 2727–2732. [Google Scholar] [CrossRef]
- Polz, M.F.; Alm, E.J.; Hanage, W.P. Horizontal gene transfer and the evolution of bacterial and archaeal population structure. Trends Genet. 2013, 29, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Letzel, A.C.; Pidot, S.J.; Hertweck, C. A genomic approach to the cryptic secondary metabolome of the anaerobic world. Nat. Prod. Rep. 2013, 30, 392–428. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Cohen, M.F.; Meeks, J.C. A polyketide-synthase-like gene is involved in the synthesis of heterocyst glycolipids in Nostoc punctiforme strain ATCC 29133. Arch. Microbiol. 1997, 167, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Sarma, T.A. Handbook of Cyanobacteria; CRC Press: New York, NY, USA, 2012. [Google Scholar]
- Bauersachs, T.; Speelman, E.N.; Hopmans, E.C.; Reichart, G.J.; Schouten, S.; Damste, J.S. Fossilized glycolipids reveal past oceanic N2 fixation by heterocystous cyanobacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 19190–19194. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.G.; Kuner, J.; Rosenzweig, B.; Lippmeier, J.C.; Roessler, P.; Zirkle, R. Biochemical characterization of polyunsaturated fatty acid synthesis in Schizochytrium: Release of the products as free fatty acids. Plant Physiol. Biochem. 2009, 47, 472–478. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, K.; Hashimoto, M.; Hori, R.; Adachi, T.; Okuyama, H.; Orikasa, Y.; Nagamine, T.; Shimizu, S.; Ueno, A.; Morita, N. Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use. Mar. Drugs 2016, 14, 94. https://doi.org/10.3390/md14050094
Yoshida K, Hashimoto M, Hori R, Adachi T, Okuyama H, Orikasa Y, Nagamine T, Shimizu S, Ueno A, Morita N. Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use. Marine Drugs. 2016; 14(5):94. https://doi.org/10.3390/md14050094
Chicago/Turabian StyleYoshida, Kiyohito, Mikako Hashimoto, Ryuji Hori, Takumi Adachi, Hidetoshi Okuyama, Yoshitake Orikasa, Tadashi Nagamine, Satoru Shimizu, Akio Ueno, and Naoki Morita. 2016. "Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use" Marine Drugs 14, no. 5: 94. https://doi.org/10.3390/md14050094
APA StyleYoshida, K., Hashimoto, M., Hori, R., Adachi, T., Okuyama, H., Orikasa, Y., Nagamine, T., Shimizu, S., Ueno, A., & Morita, N. (2016). Bacterial Long-Chain Polyunsaturated Fatty Acids: Their Biosynthetic Genes, Functions, and Practical Use. Marine Drugs, 14(5), 94. https://doi.org/10.3390/md14050094