
A Review on the Valorization of Macroalgal Wastes for Biomethane Production


Abstract
:1. Why Use Macroalgal Residues for Biomethane Production?
2. Chemical Composition
3. Basics of Anaerobic Digestion and Microbial Biomass Conversion
4. Biomethane Potential of Macroalgae
5. Optimizing AD and Applying Pretreatment to Improve Biodegradability
6. Growth Conditions of Macroalgae
7. Occurrence of Marine Eutrophication and Improvement of Coastal Management
8. Prediction of Macroalgal Growth Using Satellite Imagery
9. Macroalgal Biomass Obtained from Industrial Wastes
10. Economic Aspects and Relevance
11. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Armaroli, N.; Balzani, V. The Legacy of Fossil Fuels. Chemistry? Asian J. 2011, 6, 768–784. [Google Scholar] [CrossRef] [PubMed]
- Deublein, D.; Steinhauser, A. Biogas from Waste and Renewable Resources—An Introduction; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2008; p. 443. [Google Scholar]
- IEA. Key World Energy Statistics 2014. Available online: https://www.iea.org/publications/freepublications/publication/KeyWorld2014.pdf (accessed on 13 October 2015).
- Nehring, R. Traversing the Mountaintop: World Fossil Fuel Production to 2050. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 3067–3079. [Google Scholar] [CrossRef] [PubMed]
- Pacala, S.; Socolow, R. Stabilization Wedges: Solving the Climate Problem for the Next 50 Years with Current Technologies. Science 2004, 305, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Kraan, S. Mass-Cultivation of Carbohydrate Rich Macroalgae, a Possible Solution for Sustainable Biofuel Production. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 27–46. [Google Scholar] [CrossRef]
- Mol, A.P.J. Boundless Biofuels? Between Environmental Sustainability and Vulnerability. Sociol. Rural. 2007, 47, 297–315. [Google Scholar] [CrossRef]
- Grigg, J. The Health Effects of Fossil Fuel Derived Particles. Arch. Dis. Child. 2002, 86, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Machol, B.; Rizk, S. Economic Value of U.S. Fossil Fuel Electricity Health Impacts. Environ. Int. 2013, 52, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Gamborg, C.; Millar, K.; Shortall, O.; Sandøe, P. Bioenergy and Land use: Framing the Ethical Debate. J. Agric. Environ. Ethics 2012, 25, 909–925. [Google Scholar] [CrossRef]
- Fargione, J.; Hill, J.; Tilman, D.; Polasky, S.; Hawthorne, P. Land Clearing and the Biofuel Carbon Debt. Science 2008, 319, 1235–1237. [Google Scholar] [CrossRef] [PubMed]
- Crutzen, P.J.; Mosier, A.R.; Smith, K.A.; Winiwarter, W. N2O Release from Agro-Biofuel Production Negates Global Warming Reduction by Replacing Fossil Fuels. Atmos. Chem. Phys. Discuss. 2008, 8, 389–395. [Google Scholar] [CrossRef]
- Menegaki, A. Valuation for Renewable Energy: A Comparative Review. Renew. Sustain. Energy Rev. 2008, 12, 2422–2437. [Google Scholar] [CrossRef]
- Parguel, B.; Benoît-Moreau, F.; Larceneux, F. How Sustainability Ratings might Deter ‘Greenwashing’: A Closer Look at Ethical Corporate Communication. J. Bus. Ethics 2011, 102, 15–28. [Google Scholar] [CrossRef]
- Cotula, L.; Dyer, N.; Vermeulen, S. Fuelling Exclusion? The Biofuels Boom and Poor People's Access to Land; FAO and IIED: London, UK, 2008; Available online: http://pubs.iied.org/pdfs/12551IIED.pdf (accessed on 12 October 2015).
- Cotula, L. The International Political Economy of the Global Land Rush: A Critical Appraisal of Trends, Scale, Geography and Drivers. J. Peasant Stud. 2012, 39, 649–680. [Google Scholar] [CrossRef]
- Romagnoli, F.; Blumberga, D.; Gigli, E. Biogas from Marine Macroalgae: A New Environmental Technology—Life Cycle Inventory for a further LCA. Environ. Clim. Technol. 2010, 4, 97–108. [Google Scholar] [CrossRef]
- Langlois, J.; Sassi, J.; Jard, G.; Steyer, J.; Delgenes, J.; Hélias, A. Life Cycle Assessment of Biomethane from Offshore-Cultivated Seaweed. Biofuels Bioprod. Biorefining 2012, 6, 387–404. [Google Scholar] [CrossRef]
- Jung, K.A.; Lim, S.; Kim, Y.; Park, J.M. Potentials of Macroalgae as Feedstocks for Biorefinery. Bioresour. Technol. 2013, 135, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.A.; Lavoie, J. From First- to Third-Generation Biofuels: Challenges of Producing a Commodity from a Biomass of Increasing Complexity. Anim. Front. 2013, 3, 2–11. [Google Scholar] [CrossRef]
- Hannon, M.; Gimpel, J.; Tran, M.; Rasala, B.; Mayfield, S. Biofuels from Algae: Challenges and Potential. Biofuels 2010, 1, 763–784. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Quarterman, J.; Jin, Y. Marine Macroalgae: An Untapped Resource for Producing Fuels and Chemicals. Trends Biotechnol. 2013, 31, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; McKinley, K. Use of Macroalgae for Marine Biomass Production and CO2 Remediation: A Review. J. Appl. Phycol. 1994, 6, 45–60. [Google Scholar] [CrossRef]
- Park, J.B.K.; Craggs, R.J.; Shilton, A.N. Wastewater Treatment High Rate Algal Ponds for Biofuel Production. Bioresour. Technol. 2011, 102, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chynoweth, D.P.; Owens, J.M.; Legrand, R. Renewable Methane from Anaerobic Digestion of Biomass. Renew. Energy 2001, 22, 1–8. [Google Scholar] [CrossRef]
- Hughes, A.D.; Maeve, S.K.; Black, K.D.; Stanley, M.S. Biogas from Macroalgae: Is it Time to Revisit the Idea? Biotechnol. Biofuels 2012, 86. [Google Scholar] [CrossRef] [PubMed]
- Borines, M.G.; de Leon, R.L.; Cuello, J.L. Bioethanol Production from the Macroalgae Sargassum spp. Bioresour. Technol. 2013, 138, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Suganya, T.; Gandhi, N.N.; Renganathan, S. Production of Algal Biodiesel from Marine Macroalgae Enteromorpha Compressa by Two Step Process: Optimization and Kinetic Study. Bioresour. Technol. 2013, 128, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Dave, A.; Huang, Y.; Rezvani, S.; McIlveen-Wright, D.; Novaes, M.; Hewitt, N. Techno-Economic Assessment of Biofuel Development by Anaerobic Digestion of European Marine Cold-Water Seaweeds. Bioresour. Technol. 2013, 135, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.D.C. Biogas as an energy option: An overview. In The Biogas Handbook: Science, Production and Applications; Wellinger, A., Murphy, J., Baxter, D., Eds.; Woodhead Publishing: Cambridge, UK, 2013; p. 1. [Google Scholar]
- Braun, R.; Weiland, P.; Wellinger, A. Biogas from Energy Crop Digestion—IEA Bioenergy Task 37. Available online: http://www.iea-biogas.net/files/daten-redaktion/download/energycrop_def_Low_Res.pdf (accessed on 7 April 2016).
- Persson, M. Evaluation of Upgrading Techniques for Biogas. 2003. Available online: https://cdm.unfccc.int/filestorage/E/6/T/E6TUR2NNQW9O83ET10CX8HTE4WXR2O/Evaluation%20of%20Upgrading%20Techniques%20for%20Biogas.pdf?t=Nld8bzVhM295fDCTzmnnYsQ5c3s1NNw1XMZf (accessed on 7 April 2016).
- Börjesson, P.; Mattiasson, B. Biogas as a Resource-Efficient Vehicle Fuel. Trends Biotechnol. 2008, 26, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Smyth, B.M.; Murphy, J.D.; O’Brien, C.M. What is the Energy Balance of Grass Biomethane in Ireland and Other Temperate Northern European Climates? Renew. Sustain. Energy Rev. 2009, 13, 2349–2360. [Google Scholar] [CrossRef]
- Orr, J.; Sarmiento, J. Potential of Marine Macroalgae as a Sink for CO2: Constraints from a 3-D General Circulation Model of the Global Ocean. Water Air Soil Pollut. 1992, 64, 405–421. [Google Scholar] [CrossRef]
- Kelly, M.S.; Dworjanyn, S. The Potential of Marine Biomass for Anaerobic Biogas Production—A Feasibility Study with Recommendations for further Research. 2008. Available online: http://www.thecrownestate.co.uk/media/5765/marine_biomass_anaerobic_biogas.pdf (accessed on 12 October 2015).
- Vanegas, C.H.; Bartlett, J. Green Energy from Marine Algae: Biogas Production and Composition from the Anaerobic Digestion of Irish Seaweed Species. Environ. Technol. 2013, 34, 2277–2283. [Google Scholar] [CrossRef] [PubMed]
- N‘Yeurt, A.D.R.; Chynoweth, D.P.; Capron, M.E.; Stewart, J.R.; Hasan, M.A. Negative Carbon Via Ocean Afforestation. Process Saf. Environ. Prot. 2012, 90, 467–474. [Google Scholar] [CrossRef]
- Filipkowska, A.; Lubecki, L.; Szymczak-Żyła, M.; Kowalewska, G.; Żbikowski, R.; Szefer, P. Utilisation of Macroalgae from the Sopot Beach (Baltic Sea). Oceanologia 2008, 50, 255–273. [Google Scholar]
- Allen, E.; Browne, J.; Hynes, S.; Murphy, J.D. The Potential of Algae Blooms to Produce Renewable Gaseous Fuel. Waste Manag. 2013, 33, 2425–2433. [Google Scholar] [CrossRef] [PubMed]
- Bucholc, K.; Szymczak-Żyła, M.; Lubecki, L.; Zamojska, A.; Hapter, P.; Tjernström, E.; Kowalewska, G. Nutrient Content in Macrophyta Collected from Southern Baltic Sea Beaches in Relation to Eutrophication and Biogas Production. Sci. Total Environ. 2014, 473, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Charlier, R.H.; Morand, P.; Finkl, C.W. How Brittany and Florida Coasts Cope with Green Tides. Int. J. Environ. Stud. 2008, 65, 191–208. [Google Scholar] [CrossRef]
- Shilton, A.; Guieysse, B. Sustainable Sunlight to Biogas is via Marginal Organics. Curr. Opin. Biotechnol. 2010, 21, 287–291. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2014. Available online: http://www.fao.org/docrep/016/i2727e/i2727e.pdf (accessed on 13 October 2015).
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G. An Overview of the Chemical Composition of Biomass. Fuel 2010, 89, 913–933. [Google Scholar] [CrossRef]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. In Seaweed: Ecology, Nutrient Composition and Medicinal Uses; Pomin, V.H., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 15–47. [Google Scholar]
- Jard, G.; Marfaing, H.; Carrère, H.; Delgenes, J.P.; Steyer, J.P.; Dumas, C. French Brittany Macroalgae Screening: Composition and Methane Potential for Potential Alternative Sources of Energy and Products. Bioresour. Technol. 2013, 144, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Morand, P.; Carpentier, B.; Charlier, R.H.; Mazé, J.; Orlandini, M.; Plunkett, B.A.; de Waart, J. Bioconversion of seaweeds. In Seaweed Resources in Europe: Uses and Potential; Guiry, M.D., Blunden, G., Eds.; John Wiley & Sons: Chichester, UK, 1991; pp. 95–148. [Google Scholar]
- Horn, S.J. Seaweed Biofuels: Production of Biogas and Bioethanol from Brown Macroalgae; VDM, Verlag Dr. Müller: Saarbrücken, Germany, 2009; p. 104. [Google Scholar]
- Vergara-Fernandez, A.; Vargas, G.; Alarcan, N.; Velasco, A. Evaluation of Marine Algae as a Source of Biogas in a Two-Stage Anaerobic Reactor System. Biomass Bioenergy 2008, 32, 338–344. [Google Scholar] [CrossRef]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential Antioxidant Capacity of Sulfated Polysaccharides from the Edible Marine Brown Seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Moen, E. Biological Degradation of Brown Seaweeds. Ph.D. Thesis, Norwegian University of Science and Technology, Trondheim, Norway, November 1997. [Google Scholar]
- Reith, J.H.; Deurwaarder, E.P.; Hemmes, K. Bio-Offshore: Grootschalige Teelt Van Zeewieren in Combinatie Met Offshore Windparken in De Noordzee; Energy Research Centre of the Netherlands: Petten, The Netherlands, 2005; p. 137. [Google Scholar]
- Park, J.I.; Wu, H.C.; Lee, J.H. Production of Bio-Energy from Marine Algae: Status and Perspectives. Korean Chem. Eng. Res. 2008, 46, 833–844. [Google Scholar]
- Roesijadi, G.; Jones, S.B.; Snowden-Swan, L.J.; Zhu, Y. Macroalgae as a Biomass Feedstock: A Preliminary Analysis (PNNL-19944). 2010. Available online: https://www.pnl.gov/main/publications/external/technical_reports/PNNL-19944.pdf (accessed on 13 October 2015). [Google Scholar]
- Jang, S.; Shirai, Y.; Uchida, M.; Wakisaka, M. Production of Mono Sugar from Acid Hydrolysis of Seaweed. Afr. J. Biotechnol. 2012, 11, 1953–1963. [Google Scholar]
- Cardoso, S.M.; Carvalho, L.G.; Silva, P.J.; Rodrigues, M.S.; Pereira, O.R.; Pereira, L. Bioproducts from Seaweeds: A Review with Special Focus on the Iberian Peninsula. Curr. Org. Chem. 2014, 18, 896–917. [Google Scholar] [CrossRef]
- Barsanti, L.; Gualtieri, P. Algae—Anatomy, Biochemistry and Biotechnology; CRC Press Taylor & Francis Group: Boca Ratom, FL, USA, 2006. [Google Scholar]
- MacArtain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional Value of Edible Seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.M.; Toop, T.A.; Donnison, I.S.; Gallagher, J.A. Seasonal Variation in Laminaria digitata and its Impact on Biochemical Conversion Routes to Biofuels. Bioresour. Technol. 2011, 102, 9976–9984. [Google Scholar] [CrossRef] [PubMed]
- Chynoweth, D.P. Review of Biomethane from Marine Biomass. 2002. Available online: http://abe.ufl.edu/chyn/download/Publications_DC/Reports/marinefinal_FT.pdf (accessed on 25 May 2016).
- Østgaard, K.; Indergaard, M.; Markussen, S.; Knutsen, S.; Jensen, A. Carbohydrate Degradation and Methane Production during Fermentation of Laminaria saccharina (Laminariales, Phaeophyceae). J. Appl. Phycol. 1993, 5, 333–342. [Google Scholar] [CrossRef]
- Barbot, Y.N.; Thomsen, C.; Thomsen, L.; Benz, R. Anaerobic Digestion of Laminaria japonica Waste from Industrial Production Residues in Laborarory- and Pilot-Scale. Mar. Drugs 2015, 13, 5947–5975. [Google Scholar] [CrossRef] [PubMed]
- Wellinger, A.; Murphy, J.; Baxter, D. The Biogas Handbook: Science, Production and Applications; Woodhead Publishing: Cambridge, UK, 2013; p. 512. [Google Scholar]
- Agbor, V.B.; Cicek, N.; Sparling, R.; Berlin, A.; Levin, D.B. Biomass Pretreatment: Fundamentals Toward Application. Biotechnol. Adv. 2011, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Kayembe, K.; Basosila, L.; Mpiana, P.T.; Sikulisima, P.C.; Mbuyu, K. Inhibitory Effects on Phenolic Monomers on Methanogenesis in Aaerobic Digestion. Br. Microbiol. Res. J. 2013, 3, 32–41. [Google Scholar] [CrossRef]
- Adams, J.M.; Gallagher, J.A.; Donnison, I.S. Fermentation Study on Saccharina Latissima for Bioethanol Production Considering Variable Pre-Treatments. J. Appl. Phycol. 2008, 21, 569–574. [Google Scholar] [CrossRef]
- Anastasakis, K.; Ross, A.B.; Jones, J.M. Pyrolysis Behaviour of the Main Carbohydrates of Brown Macro-Algae. Fuel 2011, 90, 598–607. [Google Scholar] [CrossRef]
- Horn, J.S.; Aasen, M.I.; Østgaard, K. Production of Ethanol from Mannitol by Zymobacter palmae. J. Ind. Microbiol. Biotechnol. 2000, 24, 51–57. [Google Scholar] [CrossRef]
- Chi, W.; Chang, Y.; Hong, S. Agar Degradation by Microorganisms and Agar-Degrading Enzymes. Appl. Microbiol. Biotechnol. 2012, 94, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Dębowski, M.; Anna, G.; Zieliński, M.; Magda, D. Efficiency of the Methane Fermentation Process of Macroalgae Biomass Originating from Puck Bay. Arch. Environ. Prot. 2013, 38, 99–107. [Google Scholar]
- Yen, H.; Brune, D.E. Anaerobic Co-Digestion of Algal Sludge and Waste Paper to Produce Methane. Bioresour. Technol. 2007, 98, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Charlier, R.H.; Morand, P.; Finkl, C.W.; Thys, A. Green Tides on the Brittany Coasts. Environ. Res. Eng. Manag. 2007, 3, 52–59. [Google Scholar]
- Migliore, G.; Alisi, C.; Sprocati, A.R.; Massi, E.; Ciccoli, R.; Lenzi, M.; Wang, A.; Cremisini, C. Anaerobic Digestion of Macroalgal Biomass and Sediments Sourced from the Orbetello Lagoon, Italy. Biomass Bioenergy 2012, 42, 69–77. [Google Scholar] [CrossRef]
- FNR. Biogas—An Introduction 2013. Available online: https://mediathek.fnr.de/media/downloadable/files/samples/b/r/brosch.biogas-2013-en-web-pdf.pdf (accessed on 13 October 2015).
- Cecchi, F.; Pavan, P.; Mata-Alvarez, J. Anaerobic Co-Digestion of Sewage Sludge—Application to the Macroalgae from the Venice Lagoon. Resour. Conserv. Recycl. 1996, 17, 57–66. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Heiske, S. Anaerobic Digestion of Macroalgae: Methane Potentials, Pre-Treatment, Inhibition and Co-Digestion. Water Sci. Technol. 2011, 64, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, A.D.; Varela, J.C. Comparison of various Microbial Inocula for the Efficient Anaerobic Digestion of Laminaria hyperborea. BMC Biotechnol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Wiese, J.; König, R. From a Black-Box to a Glass-Box System: The Attempt Towards a Plant-Wide Automation Concept for Full-Scale Biogas Plants. Water Sci. Technol. 2009, 60, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Hinks, J.; Edwards, S.; Sallis, P.J.; Caldwell, G.S. The Steady State Anaerobic Digestion of Laminaria Hyperborea—Effect of Hydraulic Residence on Biogas Production and Bacterial Community Composition. Bioresour. Technol. 2013, 143, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Baek, G.; Kim, J.; Shin, S.G.; Lee, C. Mild-Temperature Thermochemical Pretreatment of Green Macroalgal Biomass: Effects on Solubilization, Methanation, and Microbial Community Structure. Bioresour. Technol. 2016, 199, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.; FitzGerald, J.; O’Shea, R.; Xia, A.; O’Kiely, P.; Murphy, J.D. Ensiling of Seaweed for a Seaweed Biofuel Industry. Bioresour. Technol. 2015, 196, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Kleemann, M.; Meliß, M. Regenerative Energiequellen; Springer Verlag: New York, NY, USA, 1993. [Google Scholar]
- Yang, P.Y. Methane Fermentation of Hawaiian Seaweeds. In Symposium Papers—Energy from Biomass and Wastes; Institute of Gas Technology: Des Plaines, IL, USA, 1981; pp. 307–327. [Google Scholar]
- Chynoweth, D.P.; Fannin, K.F.; Srivastava, V.J. Biological Gasification of Marine Algae. Dev. Aquac. Fish. Sci. 1987, 16. Available online: http://abe.ufl.edu/chyn/download/Publications_DC/Book%20Chapters/1987%20-%20Biological%20Gasification.pdf (accessed on 13 October 2015). [Google Scholar]
- Jard, G.; Dumas, C.; Delgenes, J.P.; Marfaing, H.; Sialve, B.; Steyer, J.P.; Carrère, H. Effect of Thermochemical Pretreatment on the Solubilization and Anaerobic Biodegradability of the Red Macroalga Palmaria Palmata. Biochem. Eng. J. 2013, 79, 253–258. [Google Scholar] [CrossRef]
- Oliveira, J.V.; Alves, M.M.; Costa, J.C. Design of Experiments to Assess Pre-Treatment and Co-Digestion Strategies that Optimize Biogas Production from Macroalgae Gracilaria vermiculophylla. Bioresour. Technol. 2014, 162, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dębowski, M.; Zieliński, M.; Grala, A.; Dudek, M. Algae Biomass as an Alternative Substrate in Biogas Production technologies—Review. Renew. Sustain. Energy Rev. 2013, 27, 596–604. [Google Scholar] [CrossRef]
- Habig, C.; Ryther, J.H. Methane Production from the Anaerobic Digestion of some Marine Macrophytes. Resour. Conserv. 1983, 8, 271–279. [Google Scholar] [CrossRef]
- Singh, J.; Gu, S. Commercialization Potential of Microalgae for Biofuels Production. Renew. Sustain. Energy Rev. 2010, 14, 2596–2610. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and Microalgae: A Positive Prospect for Biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, C.; Bartlett, J. Anaerobic Digestion of Laminaria Digitata: The Effect of Temperature on Biogas Production and Composition. Waste Biomass Valoriz. 2012, 4, 509–515. [Google Scholar] [CrossRef]
- Samson, R.; Leduy, A. Biogas Production from Anaerobic Digestion of Spirulina Maxima Algal Biomass. Biotechnol. Bioeng. 1982, 24, 1919–1924. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G. Methane Production from Marine, Green Macro-Algae. Resour. Conserv. 1983, 8, 185–194. [Google Scholar] [CrossRef]
- Barbot, Y.N.; Thomsen, L.; Benz, R. Thermo-Acidic Pretreatment of Beach Macroalgae from Rügen to Optimize Biomethane Production—Double Benefit with Simultaneous Bioenergy Production and Improvement of Local Beach and Waste Management. Mar. Drugs 2015, 13, 5681–5705. [Google Scholar] [CrossRef] [PubMed]
- Rigoni-Stern, S.; Rismondo, R. Anaerobic Digestion of Nitrophilic Algal Biomass from the Venice Lagoon. Biomass 1990, 23, 179–199. [Google Scholar] [CrossRef]
- Allen, E.; Wall, D.M.; Herrmann, C.; Murphy, J.D. Investigation of the Optimal Percentage of Green Seaweed that may be Co-Digested with Dairy Slurry to Produce Gaseous Biofuel. Bioresour. Technol. 2014, 170, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, J.F.; Indergaard, M.; Østgaard, K.; Bævre, O.A.; Pedersen, T.A.; Jensen, A. Anaerobic Digestion of Laminaria spp. and Ascophyllum Nodosum and Application of End Products. Biomass 1987, 14, 1–13. [Google Scholar] [CrossRef]
- Bird, K.T.; Chynoweth, D.P.; Jerger, D.E. Effects of Marine Algal Proximate Composition on Methane Yields. J. Appl. Phycol. 1990, 2, 207–213. [Google Scholar] [CrossRef]
- Briand, X.; Morand, P. Anaerobic Digestion of Ulva sp. 1. Relationship between Ulva Composition and Methanisation. J. Appl. Phycol. 1997, 9, 511–524. [Google Scholar]
- Nkemka, V.N.; Murto, M. Exploring Strategies for Seaweed Hydrolysis: Effect on Methane Potential and Heavy Metal Mobilisation. Process Biochem. 2012, 47, 2523–2526. [Google Scholar] [CrossRef]
- Barbot, Y.N.; Falk, H.M.; Benz, R. Thermo-Acidic Pretreatment of Marine Brown Algae Fucus Vesiculosus to Increase Methane Production—A Disposal Principle for Macroalgae Waste from Beaches. J. Appl. Phycol. 2014. [Google Scholar] [CrossRef]
- Vivekanand, V.; Eijsink, V.G.H.; Horn, S.J. Biogas Production from the Brown Seaweed Saccharina Latissima: Thermal Pretreatment and Codigestion with Wheat Straw. J. Appl. Phycol. 2012, 24, 1295–1301. [Google Scholar] [CrossRef]
- Ambye-Jensen, M.; Thomsen, S.; Kadar, Z.; Meyer, A. Ensiling of Wheat Straw Decreases the Required Temperature in Hydrothermal Pretreatment. Biotechnol. Biofuels 2013, 6, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedesco, S.; Barroso, T.M.; Olabi, A.G. Optimization of Mechanical Pre-Treatment of Laminariaceae Spp. Biomass-derived biogas. Renew. Energy 2014, 62, 527–534. [Google Scholar] [CrossRef]
- Vanegas, C.H.; Bartlett, J. Biogas Production from the Anaerobic Digestion of Laminaria Digitata in a 10 L Pilot-Plant with Digestate Re-use as Fertiliser. Int. J. Ambient Energy 2013. [Google Scholar] [CrossRef]
- Tyagi, V.; Lo, S. Application of Physico-Chemical Pretreatment Methods to Enhance the Sludge Disintegration and Subsequent Anaerobic Digestion: An Up to Date Review. Rev. Environ. Sci. Biotechnol. 2011, 10, 215–242. [Google Scholar] [CrossRef]
- Montgomery, L.F.R.; Bochmann, G. Pretreatment of Feedstock for Enhanced Biogas Production; IEA Bioenergy: Dublin, Ireland, 2014. [Google Scholar] [CrossRef]
- Kratky, L.; Jirout, T. Biomass Size Reduction Machines for Enhancing Biogas Production. Chem. Eng. Technol. 2011, 34, 391–399. [Google Scholar] [CrossRef]
- Bauer, A.; Bösch, P.; Friedl, A.; Amon, T. Analysis of Methane Potentials of Steam-Exploded Wheat Straw and Estimation of Energy Yields of Combined Ethanol and Methane Production. J. Biotechnol. 2009, 142, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, A.T.; Zeeman, G. Pretreatments to Enhance the Digestibility of Lignocellulosic. Biomass. Bioresour. Technol. 2009, 100, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Kim, D.; Kim, H.; Shin, H. Optimization of Combined (Acid+thermal) Pretreatment for Fermentative Hydrogen Production from Laminaria japonica using Response Surface Methodology (RSM). Int. J. Hydrog. Energy 2011, 36, 9626–9631. [Google Scholar] [CrossRef]
- Menind, A.; Normak, A. Study on Grinding Biomass as Pre-Treatment for Biogasification. Agron. Res. Available online: http://agronomy.emu.ee/vol08Spec1/p08s119.pdf (accessed on 26 October 2015).
- Mshandete, A.; Björnsson, L.; Kivaisi, A.K.; Rubindamayugi, M.S.T.; Mattiasson, B. Effect of Particle Size on Biogas Yield from Sisal Fibre Waste. Renew. Energy 2006, 31, 2385–2392. [Google Scholar] [CrossRef]
- Tedesco, S.; Benyounis, K.Y.; Olabi, A.G. Mechanical Pretreatment Effects on Macroalgae-Derived Biogas Production in Co-Digestion with Sludge in Ireland. Energy 2013, 61, 27–33. [Google Scholar] [CrossRef]
- Marquez, P.B.; Takeuchi, H.; Hasegawa, T. Biogas Production of Biologically and Chemically-Pretreated Seaweed, Ulva spp., Under Different Conditions: Freshwater and Thalassic. J. Jpn. Inst. Energy 2015, 94, 1066–1073. [Google Scholar] [CrossRef]
- Vanegas, C.H.; Hernon, A.; Bartlett, J. Enzymatic and Organic Acid Pretreatment of Seaweed: Effect on Reducing Sugars Production and on Biogas Inhibition. Int. J. Ambient Energy 2015, 36, 2–7. [Google Scholar] [CrossRef]
- Jung, K.W.; Kim, D.H.; Shin, H.S. Continuous fermentative hydrogen and methane production from Laminaria japonica using a two-stage fermentation system with recycling of methane fermented effluent. Int. J. Hydrog. Energy 2012, 37, 15648–15657. [Google Scholar] [CrossRef]
- Li, H.; Kjerstadius, H.; Tjernström, E.; Davidsson, Å. Evaluation of Pretreatment Methods for Increased Biogas Production from Macro Algae. Available online: http://www.sgc.se/ckfinder/userfiles/files/SGC278.pdf (accessed on 26 October 2015).
- Demirel, B.; Scherer, P. Trace Element Requirements of Agricultural Biogas Digesters during Biological Conversion of Renewable Biomass to Methane. Biomass Bioenergy 2011, 35, 992–998. [Google Scholar] [CrossRef]
- Costa, J.C.; Gonçalves, P.R.; Nobre, A.; Alves, M.M. Biomethanation Potential of Macroalgae Ulva spp. and Gracilaria spp. and in Co-Digestion with Waste Activated Sludge. Bioresour. Technol. 2012, 114, 320–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T.; Amano, T.; Koike, Y.; Saiganji, A.; Saito, H. Methane Fermentation of Seaweed Biomass. In Sustainable Nonfuel Products/Production Systems from Biomass Resources (TE017); American Institute of Chemical Engineers (AIChE): New York, NY, USA, 2006. [Google Scholar]
- Matsui, T.; Koike, Y. Methane Fermentation of a Mixture of Seaweed and Milk at a Pilot-Scale Plant. J. Biosci. Bioeng. 2010, 110, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoek, C.; Mann, D.; Jahns, H.M. Algae: An Introduction to Phycology; Cambridge University Press: Cambridge, UK, 1996; p. 640. [Google Scholar]
- Carlsson, A.S.; Beilen, J.V.; Möller, R.; Clayton, D. Micro- and Macro-Algae: Utility for Industrial Applications. EPOBIO 2007. Available online: http://www.biofuelstp.eu/downloads/epobio_aquatic_report.pdf (accessed on 18 October 2015).
- Miller, G.T.; Spoolman, S. Living in the Environment: Principles, Connections, and Solutions; Brooks/Cole, Cengage Learning: Belmont, MA, USA, 2009; p. 828. [Google Scholar]
- Kim, N.; Li, H.; Jung, K.; Chang, H.N.; Lee, P.C. Ethanol Production from Marine Algal Hydrolysates using Escherichia Coli KO11. Bioresour. Technol. 2011, 102, 7466–7469. [Google Scholar] [CrossRef] [PubMed]
- Behrens, P.W. Photobioreactors and fermentors: The light and dark side of growing algae. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Inc.: West Boothbay Harbor, ME, USA, 2005; p. 189. [Google Scholar]
- Messyasz, B.; Rybak, A. Abiotic Factors Affecting the Development of Ulva sp. (Ulvophyceae; Chlorophyta) in Freshwater Ecosystems. Aquat. Ecol. 2011, 45, 75–87. [Google Scholar] [CrossRef]
- Burkholder, J.; Glasgow, H.; Cooke, J. Comparative Effects of Water-Column Nitrate Enrichment on Eelgrass Zostera marina, Shoalgrass Halodule wrightii and Widgeongrass Ruppia maritima. Mar. Ecol. Prog. Ser. 1994, 105, 121–138. [Google Scholar] [CrossRef]
- Schramm, W. Factors Influencing Seaweed Responses to Eutrophication: Some Results from EU-Project EUMAC. J. Appl. Phycol. 1999, 11, 69–78. [Google Scholar] [CrossRef]
- Sheader, A.; Moss, B. Effects of Light and Temperature on Germination and Growth of Ascophyllum nodosum (L.). Estuar. Coast. Mar. Sci. 1975, 3, 125–132. [Google Scholar] [CrossRef]
- Ohno, M. Effect of Temperature on the Growth Rate of Seaweeds in an Aquatron Culture System. Bull. Jpn. Soc. Phycol. 1977, 25, 257–263. [Google Scholar]
- McHugh, D.J. A Guide to the Seaweed Industry. Technical Paper 441. 2003. Available online: http://www.fao.org/3/a-y4765e.pdf (accessed on 18 October 2015).
- Lüning, K. Temperature Tolerance and Biogeography of Seaweeds: The Marine Algal Flora of Helgoland (North Sea) as an Example. Helgol. Mar. Res. 1984, 38, 305–317. [Google Scholar] [CrossRef]
- Lobban, C.S.; Harrison, P.J. Seaweed Ecology and Physiology; Cambridge University Press: Cambridge, UK, 1997; pp. 563–568. [Google Scholar]
- Clark, J.S.; Poore, A.G.B.; Ralph, P.J.; Doblin, M.A. Potential for Adaptation in Response to Thermal Stress in an Intertidal Macroalga. J. Phycol. 2013, 49, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Fortes, M.D.; Lüning, K. Growth Rates of North Sea Macroalgae in Relation to Temperature, Irradiance and Photoperiod. Helgol. Meeresunters. 1980, 34, 15–29. [Google Scholar] [CrossRef]
- Lobban, C.S.; Wynne, M.J. The Biology of Seaweeds; University of California Press: Berkley/Los Angeles, CA, USA, 1981; p. 786. [Google Scholar]
- Glöckner, F.O.; Fuchs, B.M.; Amann, R. Bacterioplankton Compositions of Lakes and Oceans: A First Comparison Based on Fluorescence in Situ Hybridization. Appl. Environ. Microbiol. 1999, 65, 3721–3726. [Google Scholar] [PubMed]
- Raven, J.; Taylor, R. Macroalgal Growth in Nutrient-Enriched Estuaries: A Biogeochemical and Evolutionary Perspective. Water Air Soil Pollut. Focus 2003, 3, 7–26. [Google Scholar] [CrossRef]
- Kaandorp, J.A.; Kübler, J.E. The Algorithmic Beauty of Seaweeds, Sponges and Corals; Springer-Verlag New York, LLC: New York, NY, USA, 2001; p. 205. [Google Scholar]
- Trancoso, A.R.; Saraiva, S.; Fernandes, L.; Pina, P.; Leitão, P.; Neves, R. Modelling Macroalgae using a 3D Hydrodynamic-Ecological Model in a Shallow, Temperate Estuary. Ecol. Model. 2005, 187, 232–246. [Google Scholar] [CrossRef]
- Greve, T.; Binzer, T. Which factors regulate seagrass growth and distribution. In European Seagrasses: An Introduction to Monitoring & Management; Borum, J., Duarte, C.M., Krause-Jensen, D., Greve, T.M., Eds.; EU Project M & S: Luxembourg City, Luxembourg, 2004; pp. 19–23. Available online: http://www.seagrasses.org/handbook/european_seagrasses_high.pdf (accessed on 18 October 2015).
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Mossbauer, M.; Haller, I.; Dahlke, S.; Schernewski, G. Management of Stranded Eelgrass and Macroalgae Along the German Baltic Coastline. Ocean Coast. Manag. 2012, 57, 1–9. [Google Scholar] [CrossRef]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of Excess Nutrient Inputs on Freshwater, Marine, and Terrestrial Ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Biber, P.D. Hydrodynamic Transport of Drifting Macroalgae through a Tidal Cut. Estuar. Coast. Shelf Sci. 2007, 74, 565–569. [Google Scholar] [CrossRef]
- Valiela, I.; McClelland, J.; Hauxwell, J.; Behr, P.J.; Hersh, D.; Foreman, K. Macroalgal Blooms in Shallow Estuaries: Controls and Ecophysiological and Ecosystem Consequences. Am. Soc. Limnol. Oceanogr. 1997, 42, 1105–1118. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gobler, C.J. The Green Macroalga, Ulva lactuca, Inhibits the Growth of Seven Common Harmful Algal Bloom Species via Allelopathy. Harmful Algae 2011, 10, 480–488. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and Harmful Algal Blooms: A Scientific Consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Morand, P.; Briand, X. Excessive Growth of Macroalgae—A Symptom of Environmental Disturbance. Bot. Mar. 1996, 39, 491–516. [Google Scholar] [CrossRef]
- Aleksandrov, S.V. Biological Production and Eutrophication of Baltic Sea Estuarine Ecosystems: The Curonian and Vistula Lagoons. Mar. Pollut. Bull. 2010, 61, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Smetacek, V.; Zingone, A. Green and Golden Seaweed Tides on the Rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalewska, G.; Lubecki, L.; Szymczak-Żyła, M.; Bucholc, K.; Filipkowska, A.; Gogacz, R.; Zamojska, A. Eutrophication Monitoring System Near the Sopot Beach (Southern Baltic). Ocean Coast. Manag. 2014, 98, 51–61. [Google Scholar] [CrossRef]
- Filipkowska, A.; Lubecki, L.; Szymczak-Żyła, M.; Łotocka, M.; Kowalewska, G. Factors Affecting the Occurrence of Algae on the Sopot Beach (Baltic Sea). Oceanologia 2009, 51, 233–262. [Google Scholar] [CrossRef]
- Knee, K.L.; Street, J.H.; Grossman, E.E.; Boehm, A.B.; Paytan, A. Nutrient Inputs to the Coastal Ocean from Submarine Groundwater Discharge in a Groundwater-Dominated System: Relation to Land use (Kona Coast, Hawaii, U.S.A.). Limnol. Oceanogr. 2010, 55, 1105–1122. [Google Scholar] [CrossRef]
- Kersen, P.; Orav-Kotta, H.; Kotta, J.; Kukk, H. Effect of Abiotic Environment on the Distribution of the Attached and Drifting Red Algae Furcellaria lumbricalis in the Estonian Coastal Sea. Estonian J. Ecol. 2009, 58, 245–258. [Google Scholar] [CrossRef]
- Yan, T.; Zhou, M.J. Environmental and Health Effects Associated with Harmufl Algal Blooms and Marine Algal Toxins in China. Biomed. Environ. Sci. 2004, 17, 165–176. [Google Scholar] [PubMed]
- Roberts, D.A.; de Nys, R.; Paul, N.A. The Effect of CO2 on Algal Growth in Industrial Waste Water for Bioenergy and Bioremediation Applications. PLoS ONE 2013, 8, e81631. [Google Scholar] [CrossRef] [PubMed]
- Risén, E.; Tatarchenko, O.; Gröndahl, F.; Malmström, M.E. Harvesting of Drifting Filamentous Macroalgae in the Baltic Sea: An Energy Assessment. J. Renew. Sustain. Energy 2014, 6. [Google Scholar] [CrossRef]
- Nash, C.E. The History of Aquaculture, 1st ed.; Wiley-Blackwell: Ames, IA, USA, 2011; p. 244. [Google Scholar]
- Lorentsen, S.; Sjøtun, K.; Grémillet, D. Multi-Trophic Consequences of Kelp Harvest. Biol. Conserv. 2010, 143, 2054–2062. [Google Scholar] [CrossRef]
- Vodegel, S.; Kristin, A.; Thomsen, C. Strandalgen Als Energierohstoff—Eine Verwertungsmöglichkeit? Müll Abfall 2011, 7, 319–321. [Google Scholar]
- Liu, D.; Keesing, J.K.; Xing, Q.; Shi, P. World’s Largest Macroalgal Bloom Caused by Expansion of Seaweed Aquaculture in China. Mar. Pollut. Bull. 2009, 58, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Long, T.; Meng, G.; Wu, L.; Zhang, X.; Guo, W. Numerical Simulation for Effects of Hydrodynamic Condition on Algae Growth in Chongqing Reaches of Jialing River. Ecol. Model. 2011, 222, 112–119. [Google Scholar] [CrossRef]
- Cui, T.; Zhang, J.; Sun, L.; Jia, Y.; Zhao, W.; Wang, Z.; Meng, J. Satellite Monitoring of Massive Green Macroalgae Bloom (GMB): Imaging Ability Comparison of Multi-Source Data and Drifting Velocity Estimation. Int. J. Remote Sens. 2012, 33, 5513–5527. [Google Scholar] [CrossRef]
- Stephens, M.P.; Samuels, G.; Olson, D.B.; Fine, R.A.; Takahashi, T. Sea-Air Flux of CO2 in the North Pacific using Shipboard and Satellite Data. J. Geophys. Res. Oceans 1995, 100, 13571–13583. [Google Scholar] [CrossRef]
- McCain, C.; Hooker, S.; Feldman, G.; Bontempi, P. Satellite Data for Ocean Biology, Biogeochemistry, and Climate Research. Eos Trans. Am. Geophys. Union 2006, 87, 337–343. [Google Scholar] [CrossRef]
- Filippi, J.; Komatsu, T.; Tanaka, K. Simulation of Drifting Seaweeds in East China Sea. Ecol. Inf. 2010, 5, 67–72. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Mouritsen, J.D.; Johansen, M. Seaweed: Edible, Available, and Sustainable; University of Chicago Press: Chicago, IL, USA, 2013; p. 272. [Google Scholar]
- Buck, B.H.; Buchholz, C.M. The Offshore-Ring: A New System Design for the Open Ocean Aquaculture of Macroalgae. J. Appl. Phycol. 2004, 16, 355–368. [Google Scholar] [CrossRef]
- Mulbry, W.W.; Wilkie, A.C. Growth of Benthic Freshwater Algae on Dairy Manures. J. Appl. Phycol. 2001, 13, 301–306. [Google Scholar] [CrossRef]
- López-Mosquera, M.E.; Fernández-Lema, E.; Villares, R.; Corral, R.; Alonso, B.; Blanco, C. Composting Fish Waste and Seaweed to Produce a Fertilizer for use in Organic Agriculture. Procedia Environ. Sci. 2011, 9, 113–117. [Google Scholar] [CrossRef]
- Aderhold, D.; Williams, C.J.; Edyvean, R.G.J. The Removal of Heavy-Metal Ions by Seaweeds and their Derivatives. Bioresour. Technol. 1996, 58, 1–6. [Google Scholar] [CrossRef]
- Rodrigueza, M.R.C.; Montaño, M.N.E. Bioremediation Potential of Three Carrageenophytes Cultivated in Tanks with Seawater from Fish Farms. J. Appl. Phycol. 2007, 19, 755–762. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Azevedo, C.A.A.; Trigueiro, T.G.; Pereira, D.C.; Carneiro, M.A.A.; Camara, M.R. Bioremediation of Aquaculture Wastewater using Macroalgae and Artemia. Int. Biodeterior. Biodegrad. 2011, 65, 253–257. [Google Scholar] [CrossRef]
- Edyvean, R.G.J.; Stanley, I.M.; Stanley, S.O. Biogas Production from Seaweed Waste Following Alginate Extraction. In Biodeterioration 7; Houghton, D.R., Smith, R.N., Eggins, H.O.W., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 819–824. [Google Scholar] [CrossRef]
- Kerner, K.N.; Hanssen, J.F.; Pedersen, T.A. Anaerobic Digestion of Waste Sludges from the Alginate Extraction Process. Bioresour. Technol. 1991, 37, 17–24. [Google Scholar] [CrossRef]
- Carpentier, B.; Festino, C.; Aubart, C. Anaerobic Digestion of Flotation Sludges from the Alginic Acid Extraction Process. Biol. Wastes 1988, 23, 269–278. [Google Scholar] [CrossRef]
- Ferrera-Lorenzo, N.; Fuente, E.; Suárez-Ruiz, I.; Gil, R.R.; Ruiz, B. Pyrolysis Characteristics of a Macroalgae Solid Waste Generated by the Industrial Production of Agar–Agar. J. Anal. Appl. Pyrolysis 2014, 105, 209–216. [Google Scholar] [CrossRef]
- Zhao, H.; Yan, H.; Dong, S.; Zhang, Y.; Sun, B.; Zhang, C.; Ai, Y.; Chen, B.; Liu, Q.; Sui, T.; et al. Thermogravimetry Study of the Pyrolytic Characteristics and Kinetics of Macro-Algae Macrocystis Pyrifera Residue. J. Therm. Anal. Calorim. 2013, 111, 1685–1690. [Google Scholar] [CrossRef]
- Manara, P.; Francavilla, M.; Monteleone, M.; Zabaniotou, A. Thermogravimetric Characteristics and Pyrolysis of Red Seaweed Gracilaria Gracilis Residues. Available online: http://athens2014.biowaste.gr/pdf/manara_et_al.pdf (accessed on 18 October 2015).
- Park, J.; Yoon, J.; Park, H.; Lim, D.J.; Kim, S. Anaerobic Digestibility of Algal Bioethanol Residue. Bioresour. Technol. 2012, 113, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Clarke, W.; Pratt, S. Composting of Waste Algae: A Review. Waste Manag. 2014, 34, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Buffiere, P.F.; Loisel, D.; Bernet, N.; Delgenes, J. Towards New Indicators for the Prediction of Solid Waste Anaerobic Digestion Properties. Water Sci. Technol. 2006, 53, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Couillard, D.; Li, J.F. Assessment of Manure-Application Effects upon the Runoff Water Quality by Algal Assays and Chemical Analyses. Environ. Pollut. 1993, 80, 273–279. [Google Scholar] [CrossRef]
- Tambone, F.; Scaglia, B.; D’Imporzano, G.; Schievano, A.; Orzi, V.; Salati, S.; Adani, F. Assessing Amendment and Fertilizing Properties of Digestates from Anaerobic Digestion through a Comparative Study with Digested Sludge and Compost. Chemosphere 2010, 81, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Makádi, M.; Tomócsik, A.; Orosz, V. Digestate: A New Nutrient Source—Review. In Biogas; Kumar, S., Ed.; InTechOpen: Rijeka, Croatia, 2012; p. 295. Available online: http://cdn.intechopen.com/pdfs/31331.pdf (accessed on 27 May 2016).
- Massi, E.; McPhail, S.J. Biomethane and Natural Gas. In Fuel Cells in the Waste-to-Energy Chain; Springer: London, UK, 2012; pp. 165–176. [Google Scholar] [CrossRef]
- Hengeveld, E.J.; Bekkering, J.; van Gemert, W.J.T.; Broekhuis, A.A. Biogas Infrastructures from Farm to Regional Scale, Prospects of Biogas Transport Grids. Biomass Bioenergy 2016, 86, 43–52. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-Value Products from Macroalgae: The Potential Uses of the Invasive Brown Seaweed, Sargassum muticum. Rev. Environ. Sci. Biol. Technol. 2015, 15, 67–88. [Google Scholar] [CrossRef]
- Taelman, S.E.; Champenois, J.; Edwards, M.D.; de Meester, S.; Dewulf, J. Comparative Environmental Life Cycle Assessment of Two Seaweed Cultivation Systems in North West Europe with a Focus on Quantifying Sea Surface Occupation. Algal Res. 2015, 11, 173–183. [Google Scholar] [CrossRef]
- Pomin, V.H. Seaweed: Ecology, Nutrient Composition, and Medicinal Uses; Nova Science Publishers: New York, NY, USA, 2011; p. 248. [Google Scholar]
- Alvarado-Morales, M.; Boldrin, A.; Karakashev, D.B.; Holdt, S.L.; Angelidaki, I.; Astrup, T. Life Cycle Assessment of Biofuel Production from Brown Seaweed in Nordic Conditions. Bioresour. Technol. 2013, 129, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Barbot, Y. Anaerobic Fermentation of Marine Algae Biomass. Ph.D. Thesis, Jacobs University Bremen, Bremen, Germany, November 2010. [Google Scholar]
- Cordell, D.; Drangert, J.; White, S. The Story of Phosphorus: Global Food Security and Food for Thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Compound | Green Algae | Red Algae | Brown Algae | Reference |
|---|---|---|---|---|
| Water content (from fresh mass) | 70%–85% | 70%–80% | 79%–90% | [54] |
| Ash | 18%–53% | 26%–48% | 33%–55% | [47] |
| Total organic | 47%–82% | 52%–74% | 44%–66% | [47] |
| Carbohydrate | 25%–50% | 30%–60% | 30%–50% | [19] |
| Polysaccharide | Alginate Cellulose Mannan Starch Ulvan | Agar Alginate Carrageenan Cellulose Lignin | Agar Alginate Carrageenan Cellulose Fucoidan Laminarin Mannitol | [19,55,56] |
| Protein | 12%–13% | 10%–16% | 7%–12% | [47] |
| Lipid | 2%–3% | 0%–3% | 0%–2% | [47] |
| Industrial extracts | Sulfated galactan, vitamins (e.g., C), antiviral and anticoagulating agents | Sulfated galactan, vitamins (e.g., C, B), mineral nutrients (e.g., iodine), agar, phycobiliproteins | Fucoidan, fucan hydrocolloids (alginate, carrageenan, agar-agar) polyphenols, mineral nutrients (e.g., iodine), pigments | [19,47,53,57,58] |
| Industrial use | Human food, food supplement, medicinal use | Human food, pet food production, thickener, emulsifier and gelling agent in industrial and lab use and for cosmetics | Human food, animal feed, alginate for textile printing, medical fiber, paper industry, cosmetics, agar as a laxative in the pharmaceutical industry, fermentative production of organic acids | [19,47,53,57,58] |
| Seaweed Species | Methane Yield (mL·g−1 VS) | Reference |
|---|---|---|
| Ascophyllum | 110 | [98] |
| Gracilaria | 280–400 | [90,91] |
| Laminaria sp. | 180–300 | [53,92] |
| Macrocystis pyrifera | 180–430 | [85,88] |
| Sargassum | 120–190 | [99] |
| Ulva lactuca | 200–480 | [53,100] |
| Pretreatment Method | Technique | Description | Examples | Increase of BMP | Reference |
|---|---|---|---|---|---|
| Physical | Mechanical | Substrate fragmentation using manual knife mills, shredders or automatic hammer mills | Laminaria digitata, Laminaria saccharina, Laminaria hyperborea | +20%–50% | [105,109,113,114,115] |
| Thermal | Heating at 125 °C–190 °C under pressure for up to an hour | Saccharina latissima | n.s. | [86,103,108] | |
| Chemical | Alkaline | Alkali pretreatment, e.g., sodium hydroxide | Ulva spp. | +27% | [116] |
| Acidic | Pretreatment with organic acids (citric acid, lactic acid, acetic acid, oxalic acid) or inorganic acids (e.g., hydrochloric acid, sulfuric acid) | Laminaria digitata, Saccharina latissima, Fucus vesiculosus | +4% | [102,117] | |
| Biological | Microbial digestion | Aerobic microbial digestion (e.g., polysaccharide hydrolyzing bacteria, methanogenic archaea) or anaerobic digestion in one- or two-stage bioreactors | Laminaria japonica, Laminaria hyperborean, Saccharina latissima | n.s. | [63,77,118] |
| Enzymatic digestion | Co-digestion with individual enzymes (e.g., pectinase, cellulase, hemicellulase, alginate lyase or protease) or with enzyme mixtures | Furcellaria lumbricalis, Fucus vesiculosus, Palmaria palmate Laminaria digitata, Saccharina latissima | +2% | [117,119] | |
| Combined processes | Steam explosion | Thermal pretreatment at 160 °C–220 °C combined with a sudden drop in pressure. | Saccharina latissima | +20% | [103,110] |
| Thermo-chemical | Thermal treatment (60 °C–220 °C) combined with the addition of different kinds of acidic or alkali reagents. | Palmaria palmate, Fucus vesiculosus | +10%–140% | [86,102,103] | |
| Biochemical | Both acidic (2.5% citric acid) and enzymatic (cellulase) pretreatments are applied to the substrate. | Laminaria digitata | +7% | [117] |
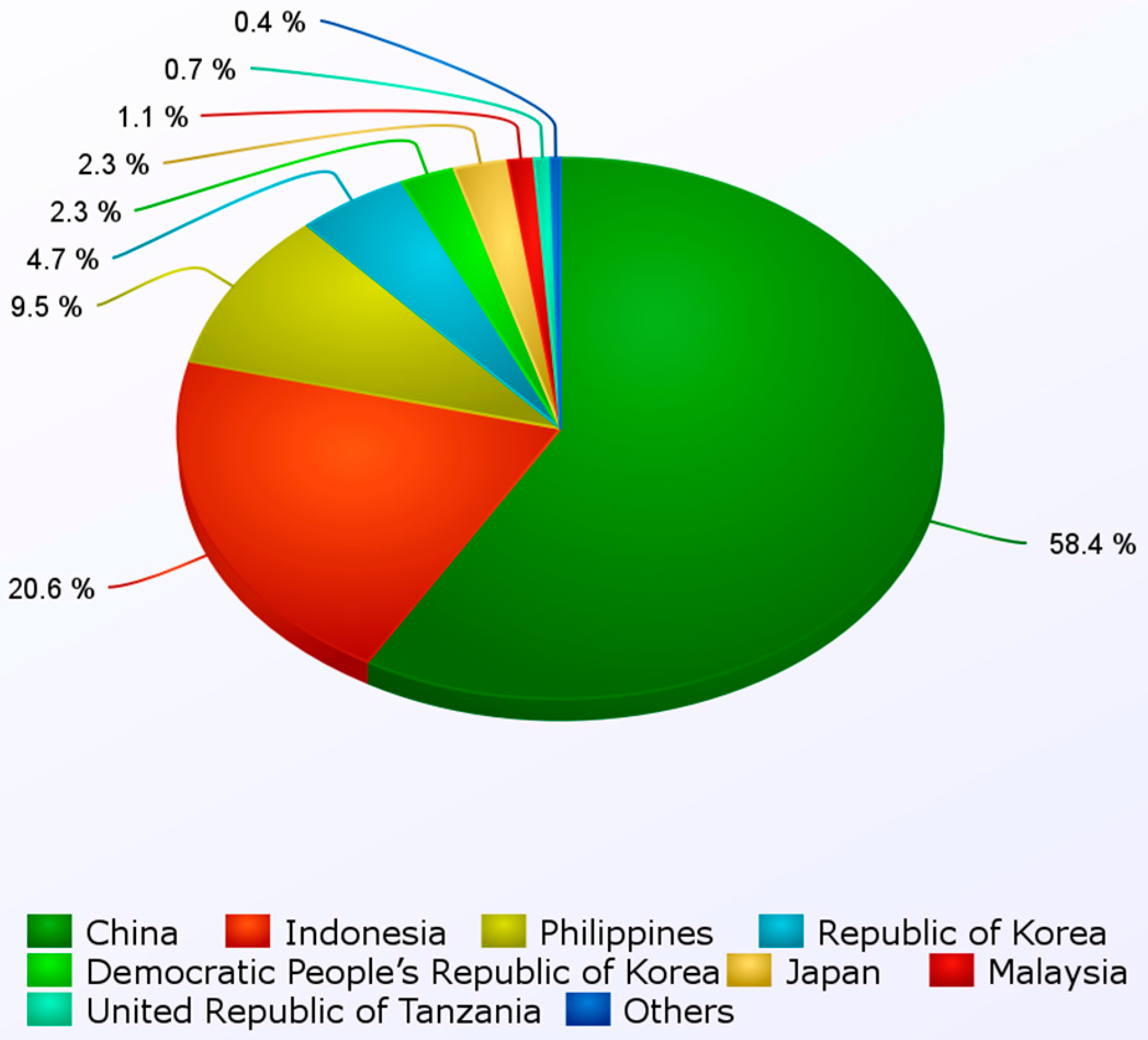
| Continent | 2008 | 2009 | 2010 | 2011 |
|---|---|---|---|---|
| Africa | 0.12 | 0.11 | 0.14 | 0.14 |
| Americas | 0.03 | 0.09 | 0.01 | 0.02 |
| Asia | 15.73 | 17.14 | 18.84 | 20.80 |
| Europe | 0.00 | 0.00 | 0.00 | 0.00 |
| Oceania | 0.00 | 0.01 | 0.01 | 0.01 |
| Total | 15.9 | 17.4 | 19.0 | 21.0 |
| Year-on-year growth rate | 5.9% | 9.3% | 9.5% | 10.4% |
| Type of Waste | Source | Organism | Quantity | Composition | CH4 Potential | Reference |
|---|---|---|---|---|---|---|
| “Macroalgae meal” | Residue from agar-agar extraction | Gelidium sesquipedale | 2000–2400 kg/day (dry powder) | High: carbon, nitrogen, hydrogen Low: ash | n.s. | [181] |
| “Macrocystis pyrifera residue” | Qingdao Mingye Seaweed Industrial Co. Ltd. (China) | Macrocystis pyrifera | n.s. | Moisture: 9.8% Ash: 59.2% VS: 18.3% Fixed carbon: 12.71% cellulose, hemicellulose, lignin | n.s. | [182] |
| Gracilaria gracilis residues | Residues from phycobiliprotein extraction | Gracilaria gracilis | n.s. | 75% VS 21% ash 4% fixed carbon | n.s. | [183] |
| Laminaria japonica residues | Remains from industrial biomass processing | Laminaria japonica | 50,000 t/year (wet mass); remains of 10%–30% biomass from downstream processing | 50.9% VS 39.2% carbohydrate 11.4% protein 0.3% lipid 49.1% ash | 172–214 mL·g−1·VS | [63] |
| Fermentation residue, saccharification residue | Residues from algal bioethanol production | Gelidium Amani | n.s. | Galactose: 52.4% Protein: 15.6% Cellulose: 14.9% Ash: 5.7% Others: 11.4% | 239–283 mL·g−1·VS | [184] |
| Alginate extraction residues | Kelco/AIL factory at Barcaldine (Scotland) | Ascophyllum spp. | n.s. | n.s. | 198–237 mL·g−1·VS | [178] |
| Alginate extraction sludge | Protan A/S, Haugesund, Norway | Laminaria hyperborea, Ascophyllum nodosum | n.s. | 78.8%–85.4% VS | 70–280 mL·g−1·VS | [179] |
| Green tide 2008 | Qingdao algae bloom (China) | Enteromorpha prolifera | 150,000–1 million t (wet mass) | Moisture: 4.85% Ash: 17,63% VS: 70.29% Fixed carbon: 7.4% | n.s. | [154,165] |
| Macroalgae bloom | Venice lagoon (Italy), bloom | Ulva rigida, Gracilaria confervoides | 40,000 t/year (wet mass) | 25.4% total solids (TS) 32.0% VS | 129–212 mL·g−1·VS | [76,96,185] |
| Green tide | Patagonia beaches (Chile), bloom | Green seaweed | 8000 t/year (wet mass) | n.s. | n.s. | [185] |
| Green tide | Brittany beaches (France), bloom | Ulva sp. | 100,000 t/year (wet mass) | n.s. | 91–200 mL·g−1·VS | [73,100] |
| Golden tide | Gulf of Mexico, bloom | Sargassum natans, Sargassum fluitans | 1 million/year | n.s. | n.s. | [154] |
| Beached macroalgae | Orbetello lagoon (Italy), bloom | Gracilariopsis longissima, Chaetomorpha linum | 5000 t/year (wet mass) | 30% TS 47% VS | 380 mL·g−1·VS | [74] |
| Product | Content (% of Dry Weight) | Value (€/t Dry Weight) |
|---|---|---|
| Alginate | 23 | 1265 |
| Mannitol | 12–21 | 645 |
| Iodine | 0.45 | 58.50 |
| Potash | 9.5 | 5.10 |
| Phosphorous | 0.3 | 2.70 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbot, Y.N.; Al-Ghaili, H.; Benz, R. A Review on the Valorization of Macroalgal Wastes for Biomethane Production. Mar. Drugs 2016, 14, 120. https://doi.org/10.3390/md14060120
Barbot YN, Al-Ghaili H, Benz R. A Review on the Valorization of Macroalgal Wastes for Biomethane Production. Marine Drugs. 2016; 14(6):120. https://doi.org/10.3390/md14060120
Chicago/Turabian StyleBarbot, Yann Nicolas, Hashem Al-Ghaili, and Roland Benz. 2016. "A Review on the Valorization of Macroalgal Wastes for Biomethane Production" Marine Drugs 14, no. 6: 120. https://doi.org/10.3390/md14060120
APA StyleBarbot, Y. N., Al-Ghaili, H., & Benz, R. (2016). A Review on the Valorization of Macroalgal Wastes for Biomethane Production. Marine Drugs, 14(6), 120. https://doi.org/10.3390/md14060120