
Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
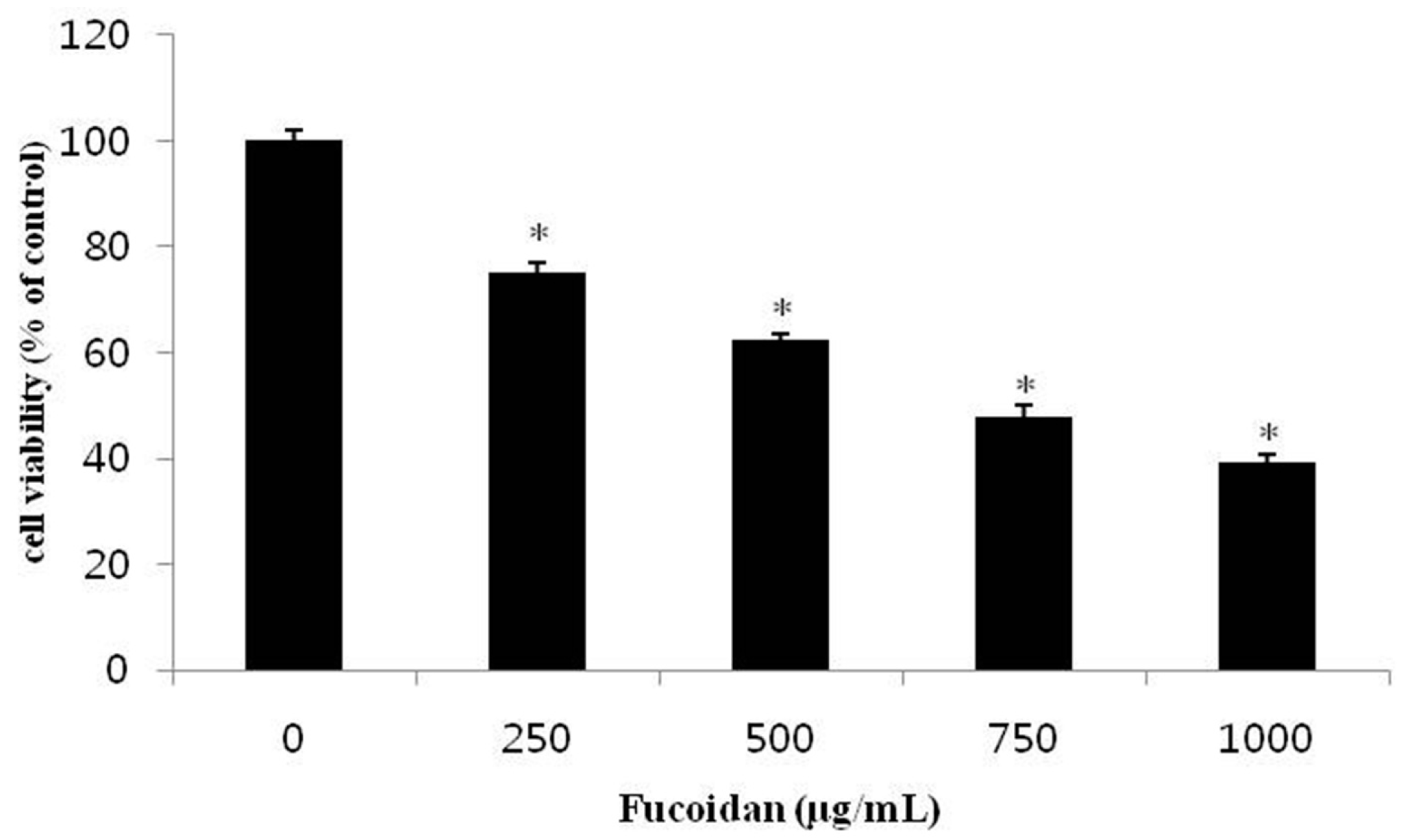
2.1. Effect of Fucoidan on the Viability of DU-145 Cancer Cells
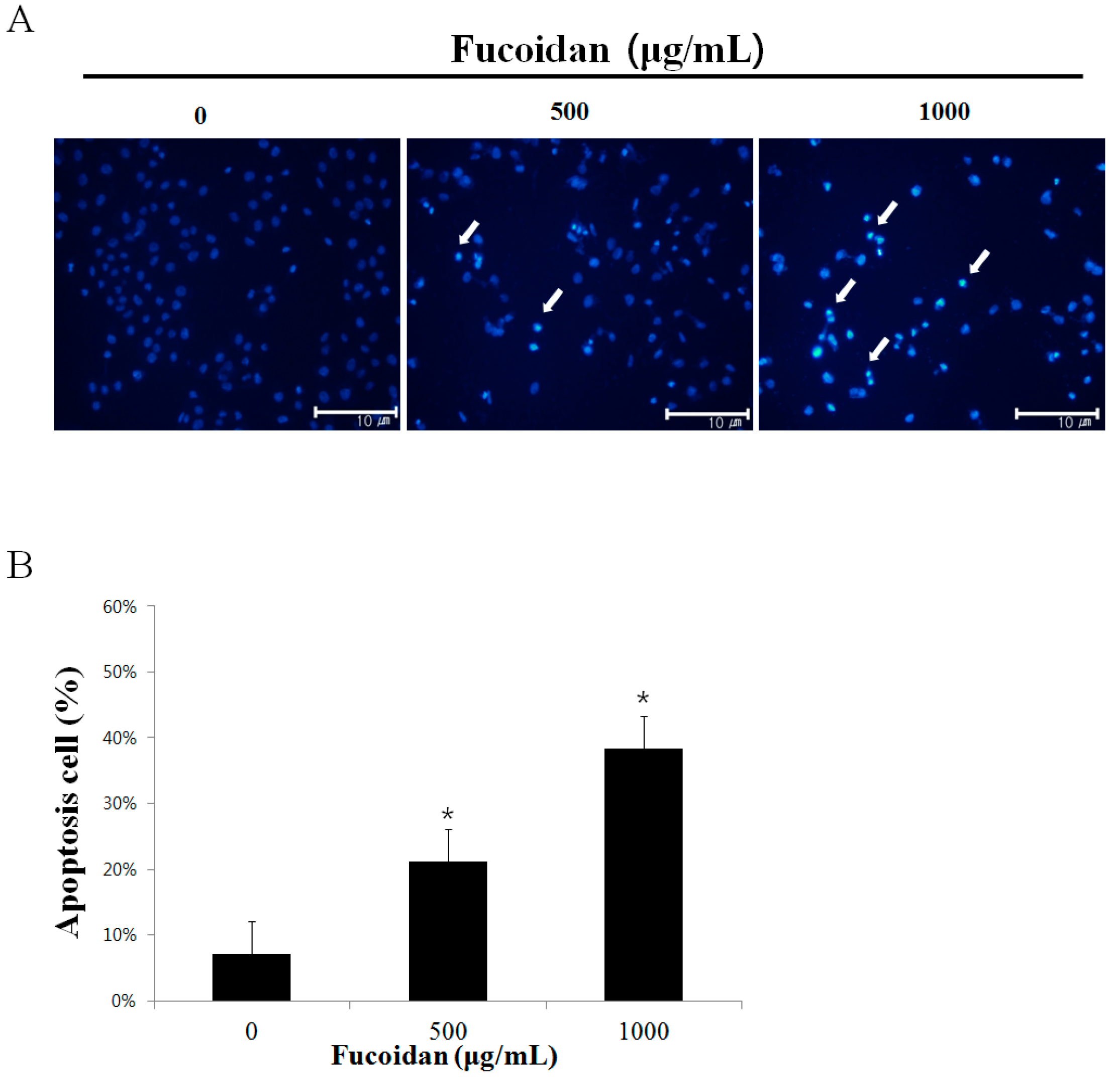
2.2. The Morphological Changes of DU-145 Cancer Cells by the Fucoidan
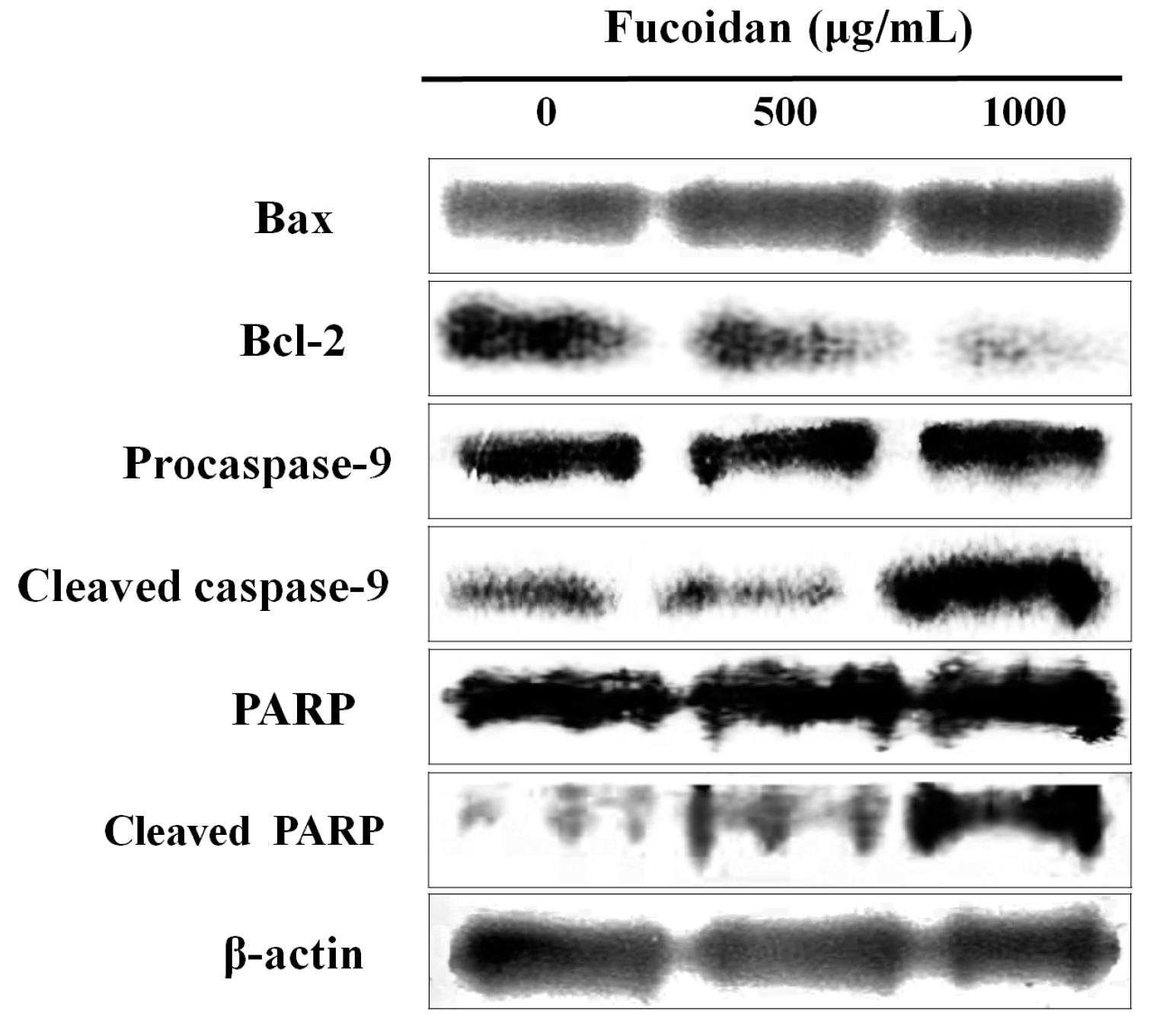
2.3. Effect of Fucoidan on the Apoptosis-Related Proteins of DU-145 Cancer Cells
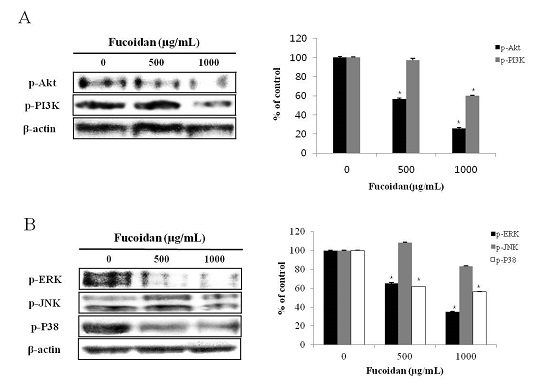
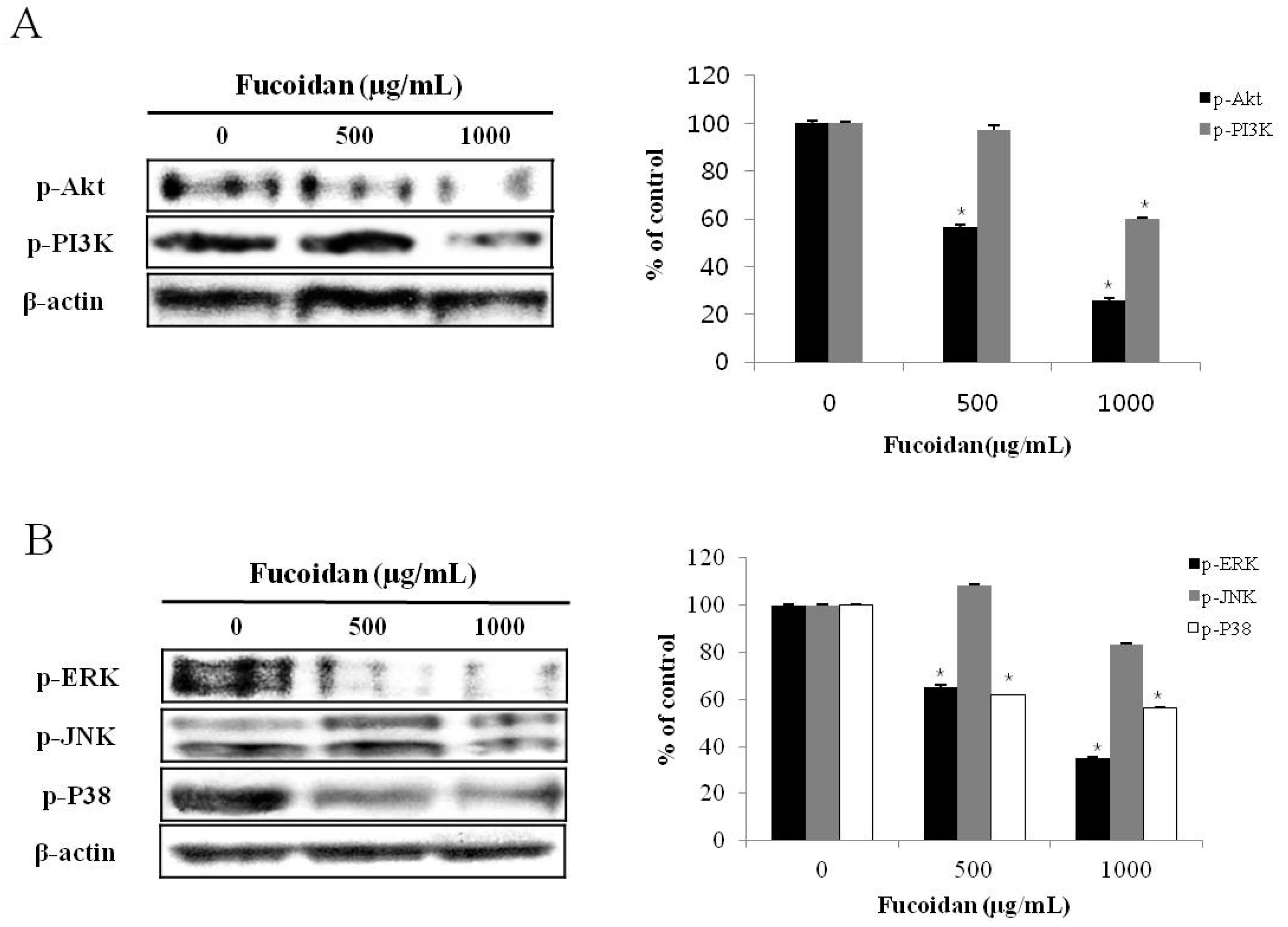
2.4. Effect of Fucoidan on PI3K/Akt Pathways in DU-145 Cancer Cells
2.5. Effect of Fucoidan on MAPK Pathways in DU-145 Cancer Cells
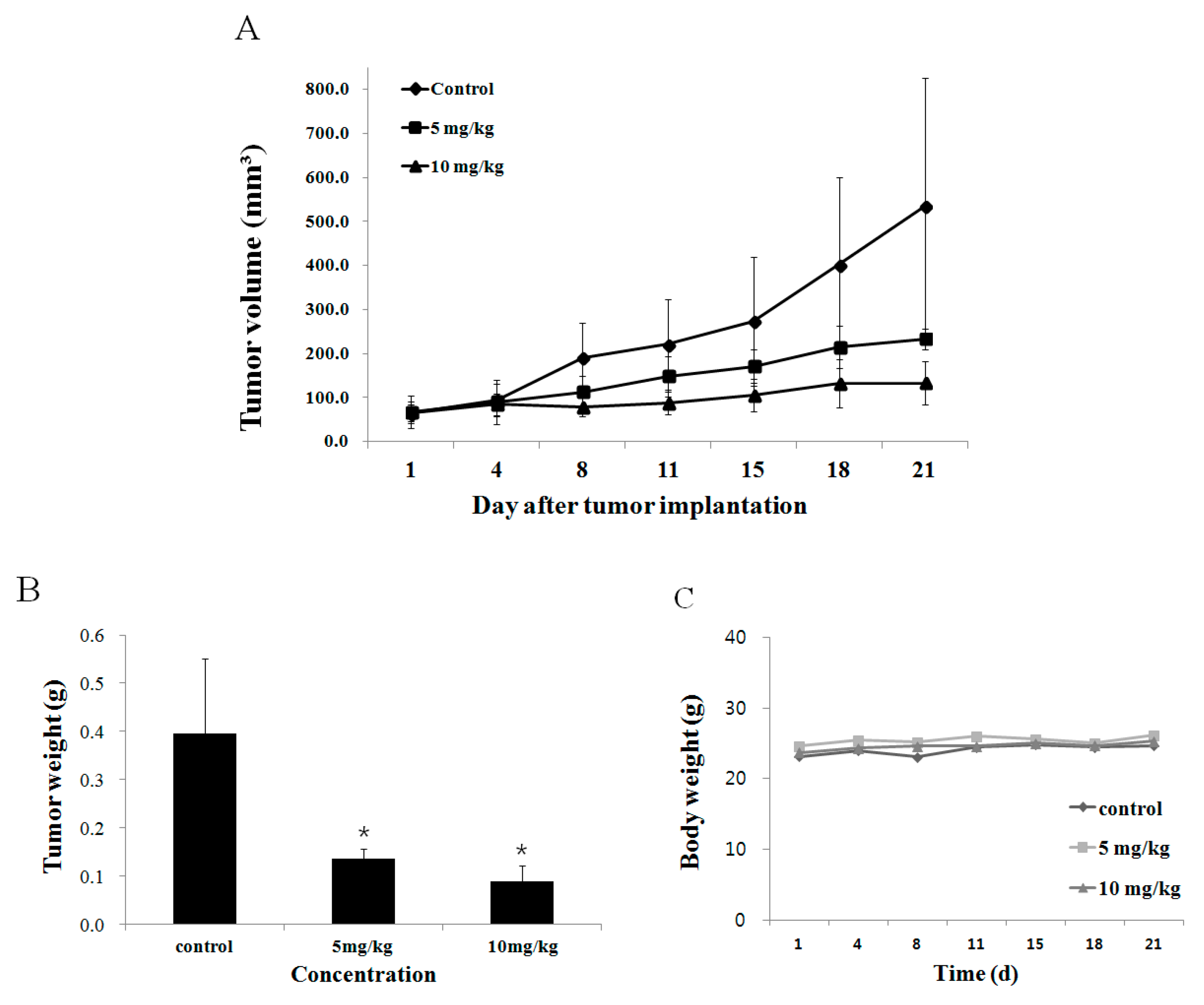
2.6. Effect of Fucoidan on Tumor Growth In Vivo Animal Model
2.7. Effect of Fucoidan on the Apoptosis Induction of DU-145 Tumor Tissue
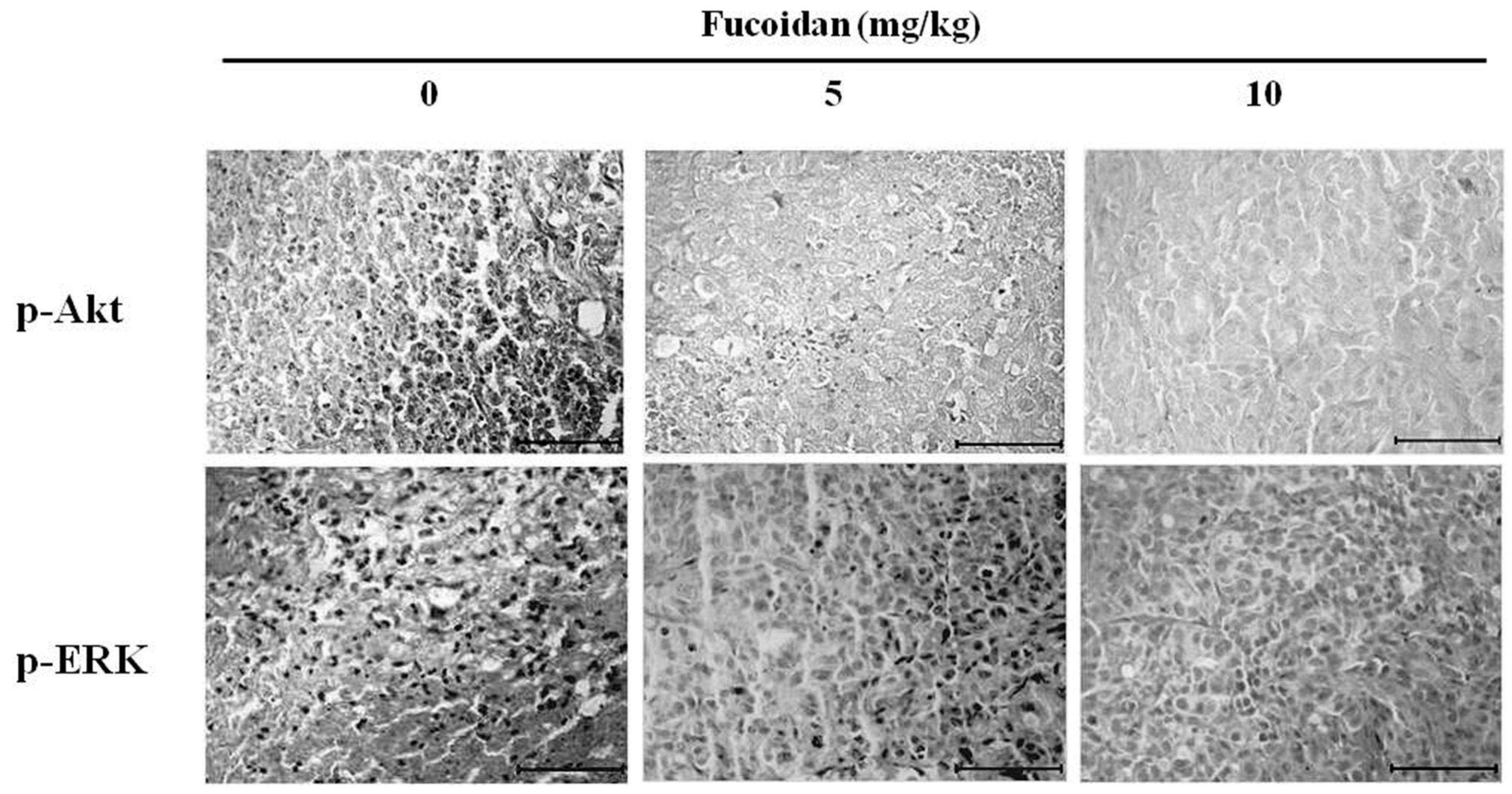
2.8. Effect of Fucoidan on Akt and ERK Expression in DU-145 Tumor Tissue
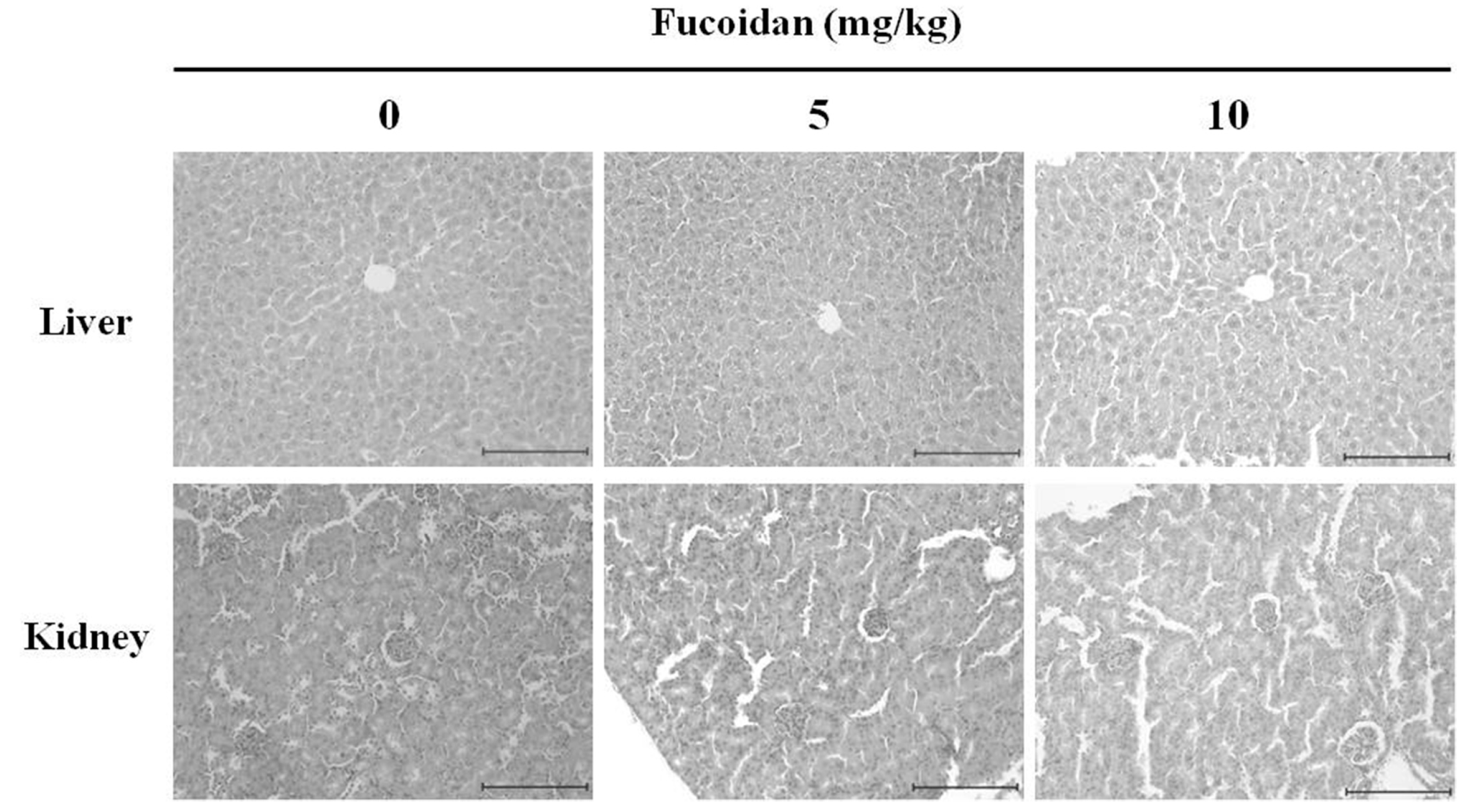
2.9. The Histopathological Changes in DU-145 Tumor Tissues by the Fucoidan
3. Materials and Methods
3.1. Chemicals, Drugs, and Antibodies
3.2. Cell Lines and Culture
3.3. Cell Viability Assay
3.4. 4′,6-diamidino-2-phenylindole (DAPI) Staining
3.5. Western Blot Analysis
3.6. Antitumor Activity In Vivo
3.7. Histological Examination
3.8. TUNEL Assay
3.9. Immunohistochemistry
3.10. Statistical Analysis
4. Conclusions
Acknowledgement
Author Contributions
Conflicts of Interest
References
- Doll, S.R. The lessons of life: keynote address to the nutrition and cancer conference. J. Cancer Res. 1992, 52, 2024–2029. [Google Scholar]
- Oh, C.M.; Won, Y.J.; Jung, K.W.; Kong, H.J.; Cho, H.; Lee, J.K.; Lee, D.H.; Lee, K.H. Cancer statistics in Korea: Incidence, mortality, survival, and prevalence in 2013. J. Cancer Res. Ther. 2014, 46, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Bowen, P.E. Effects of tomatoes and lycopene on prostate cancer prevention and treatment. J. Korean Soc. Food Sci. Nutr. 2004, 33, 455–462. [Google Scholar]
- Walsh, P.C.; Partin, A.W.; Epstein, J.I. Cancer control and quality of life following anatomical radical retropubic prostatectomy: Results at 10 years. J. Urol. 1944, 152, 1831–1836. [Google Scholar]
- Isaacs, J.T.; Lundmo, P.I.; Berges, R.; Martikainen, P.; Kyprianou, N. Androgen regulation of programmed death of normal and malignant prostatic cells. J. Androl. 1992, 13, 457–464. [Google Scholar] [PubMed]
- Kwon, M.J.; Nam, T.J. A polysaccharide of the marine alga Capsosiphon fulvescens induces apoptosis in AGS gastric cancer cells via an IGF-IR-mediated PI3K/Akt pathway. Cell Biol. Int. 2007, 31, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Gideon, T.P.; Rengasamy, R. Toxicological evaluation of fucoidan from Cladosiphon okamuranus. J. Med. Food 2008, 11, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2005, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki-Miyamoto, Y.; Yamasaki, M.; Tachibana, H.; Yamada, K. Fucoidan induces apoptosis through activation of caspase-8 on human breast cancer MCF-7 cells. J. Agric. Food Chem. 2009, 57, 8677–8682. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, P.; Wang, H.; Li, Q.; Teng, H.; Liu, Z.; Yang, W.; Hou, L.; Zou, X. Fucoidan derived from Undaria pinnatifida induces apoptosis in human hepatocellular carcinoma SMMC-7721 cells via the ROS-mediated mitochondrial pathway. Mar. Drugs 2013, 11, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.Y.; Kim, G.Y.; Chol, L.W.; Kim, N.D.; Nam, T.J.; Kwon, T.K.; Chol, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Sharmistha, S.; Akram, A.; Tuhin, G.; Paul, S.; Bimalendu, R. Polysaccharides from Sargassum tenerrimum: Structural features, chemical modification and anti-viral activity. Phytochemistry 2010, 71, 235–242. [Google Scholar]
- Han, Y.S.; Lee, J.H.; Lee, S.H. Antitumor effects of fucoidan on human colon cancer cells via activation of AKT signaling. Biomol. Ther. 2015, 23, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Carnero, A. The PKB/AKT pathway in cancer. Curr. Pharm. Des. 2010, 16, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Oshimura, M.; Ito, H. PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis 2004, 9, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Boo, H.J.; Hong, J.Y.; Kim, S.C.; Kang, J.I.; Kim, M.K.; Kim, E.J.; Hyun, J.W.; Koh, Y.S.; Yoo, E.S.; Kwon, J.M.; et al. The Anticancer Effect of Ficoidan in PC-3 Prostate Cancer cells. Mar. Drugs. 2013, 11, 2982–2999. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.; Gonzalez, F.A.; Gupta, S.; Raden, D.L.; Davis, R.J. Signal transduction within the nucleus by mitogen-activated protein kinase. J. Biol. Chem. 1992, 267, 24796–24804. [Google Scholar] [PubMed]
- Zhuang, Z.Y.; Gerner, P.; Woolf, C.J.; Ji, R.R. ERK is sequentially activated in neurons, microglia, and astrocytes by spinal nerve ligation and contributes to mechanical allodynia in this neuropathic pain model. Pain 2005, 114, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Park, S.Y.; Kim, Y.M.; Kim, D.C.; Lee, W.S.; Surh, Y.J.; Park, O.J. Suppression of mTOR via Akt-dependent and -independent mechanisms in selenium-treated colon cancer cells: Involvement of AMPKα1. Carcinogenesis 2010, 31, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, G.Y.; Nam, T.J.; Kim, D.N.; Choi, Y.H. Antiproliferative activity of fucoidan was associated with the induction of apoptosis and autophagy in AGS human gastric cancer cells. J. Food Sci. 2011, 76, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.; Cotter, T.G. Control of mitochondrial integrity by Bcl-2 family members and caspase-independent cell death. Biochim. Biophys. Acta 2004, 1644, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Xiao, Q.; Kuang, X.; Zhang, T.; Yang, Z.; Wang, L. Fucoidan inhibits proliferation of the SKM-1 acute myeloid leukaemia cell line via the activation of apoptotic pathways and production of reactive oxygen species. Mol. Med. Rep. 2015, 12, 6649–6655. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yun, S.G.; Oh, Y.N.; Lee, J.Y.; Park, H.J.; Jin, K.S.; Kwon, H.J.; Kim, B.W. Induction of G2/M arrest and apoptosis by methanol extract of Typha orientalis in human colon adenocarcinoma HT29 cells. Microbiol. Biotechnol. Lett. 2013, 41, 425–432. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, G.Y.; Moon, S.K.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Fucoidan inhibits the proliferation of human urinary bladder cancer T24 cells by blocking cell cycle progression and inducing apoptosis. Molecules 2014, 19, 5981–5998. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, J.S.; Kim, E. Fucoidan from seaweed Fucus vesiculosus inhibits migration and invasion of human lung cancer cell via PI3K-Akt-mTOR pathways. PLoS ONE 2012, 7, e50624. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, X.; Chen, Y.; Wang, X.; Chen, R. P38 and JNK MAPK, but not ERK1/2 MAPK, play important role in colchicine-induced cortical neurons apoptosis. Eur. J. Pharmacol. 2007, 576, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer properties and mechanisms of fucoidan on mouse breast cancer in vitro and in vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Zhang, M.; Jin, M.; Bai, C.; Wang, X. Potential mechanism of interleukin-8 production from lung cancer cells: An involvement of EGF-EGFR-PI3K-Akt-Erk pathway. J. Cell. Physiol. 2012, 227, 35–43. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choo, G.-S.; Lee, H.-N.; Shin, S.-A.; Kim, H.-J.; Jung, J.-Y. Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Mar. Drugs 2016, 14, 126. https://doi.org/10.3390/md14070126
Choo G-S, Lee H-N, Shin S-A, Kim H-J, Jung J-Y. Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Marine Drugs. 2016; 14(7):126. https://doi.org/10.3390/md14070126
Chicago/Turabian StyleChoo, Gang-Sik, Hae-Nim Lee, Seong-Ah Shin, Hyeong-Jin Kim, and Ji-Youn Jung. 2016. "Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression" Marine Drugs 14, no. 7: 126. https://doi.org/10.3390/md14070126
APA StyleChoo, G. -S., Lee, H. -N., Shin, S. -A., Kim, H. -J., & Jung, J. -Y. (2016). Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Marine Drugs, 14(7), 126. https://doi.org/10.3390/md14070126