
Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
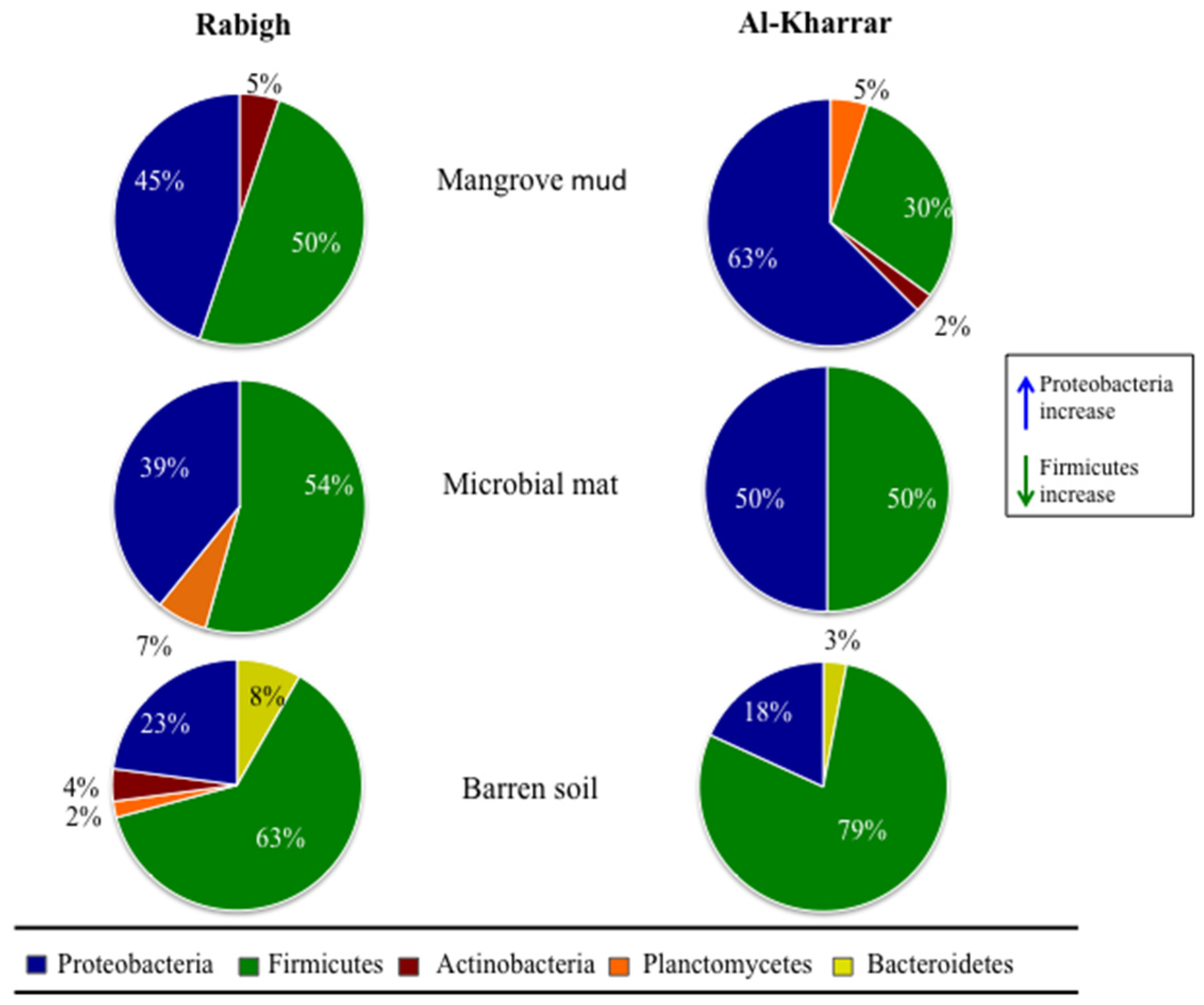
2.1. Bacterial Isolation
2.2. 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.3. Antimicrobial Screening
2.4. PCR-Screening for PKS and NRPS Domains
3. Material and Methods
3.1. Sediment Sampling
3.2. Isolation and Purification of Bacterial Strains
3.3. Molecular Identification and Phylogenetic Analysis
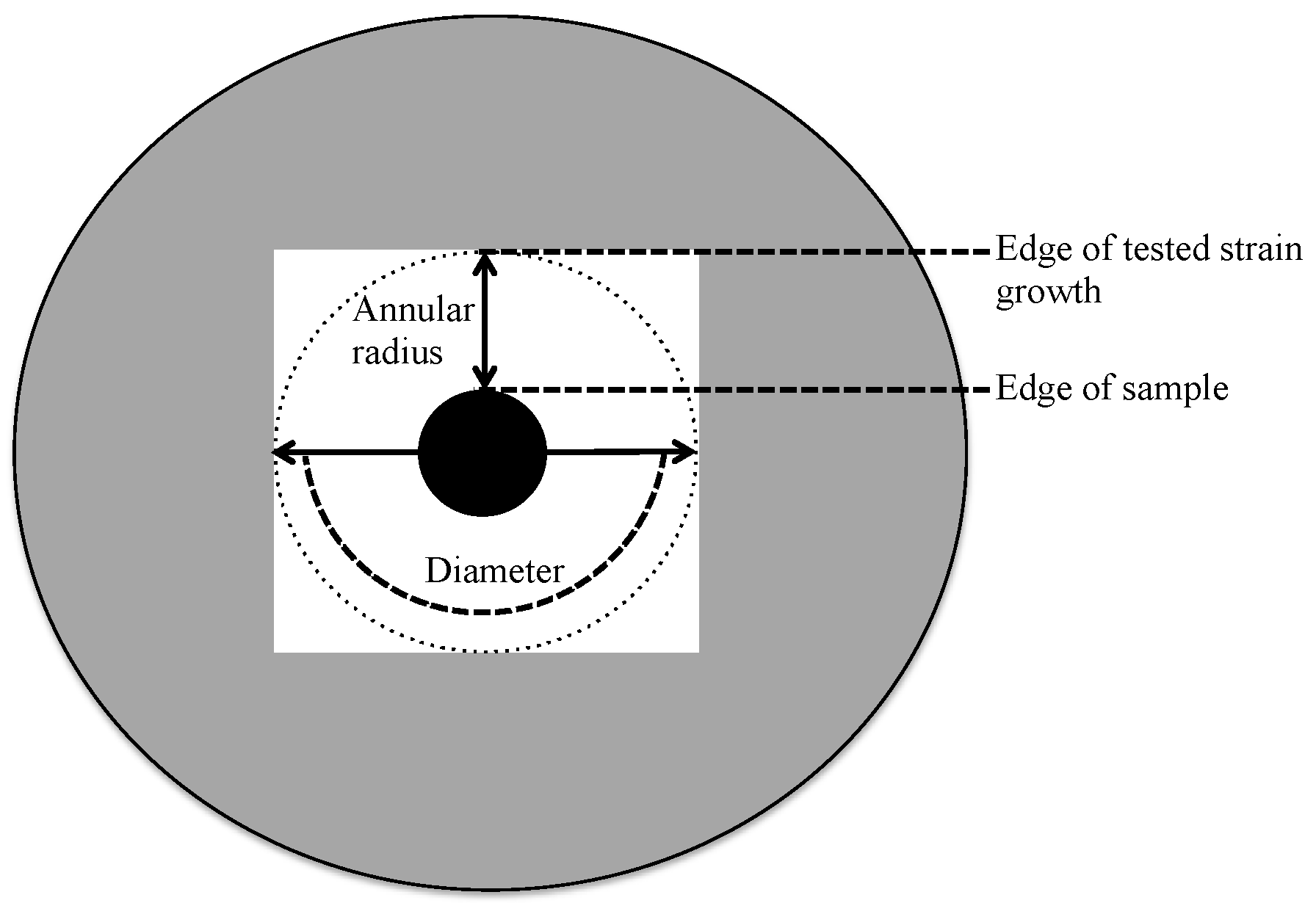
3.4. Screening for Antimicrobial Activity
3.5. PCR Screening for PKS and NRPS Gene Fragments
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carlet, J.; Collignon, P.; Goldmann, D.; Goossens, H.; Gyssens, I.C.; Harbarth, S.; Jarlier, V.; Levy, S.B.; N’Doye, B.; Pittet, D.; et al. Society’s failure to protect a precious resource: Antibiotics. Lancet 2011, 378, 369–371. [Google Scholar] [CrossRef]
- Tally, F.P.; DeBruin, M.F. Development of daptomycin for Gram-positive infections. J. Antimicrob. Chemother. 2000, 46, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Leach, K.L.; Brickner, S.J.; Noe, M.C.; Miller, P.F. Linezolid, the first oxazolidinone antibacterial agent. Ann. N. Y. Acad. Sci. 2011, 1222, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Donadio, S.; Maffioli, S.; Monciardini, P.; Sosio, M.; Jabes, D. Antibiotic discovery in the twenty-first century: Current trends and future perspectives. J. Antibiot. 2010, 63, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Amoutzias, G.D.; Chaliotis, A.; Mossialos, D. Discovery strategies of bioactive compounds synthesized by nonribosomal peptide synthetases and type-I polyketide synthases derived from marine microbiomes. Mar. Drugs 2016, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechnol. 2010, 3, 544–563. [Google Scholar] [CrossRef] [PubMed]
- Bourbouli, M.; Katsifas, E.A.; Papathanassiou, E.; Karagouni, A.D. The Kolumbo submarine volcano of Santorini island is a large pool of bacterial strains with antimicrobial activity. Arch. Microbiol. 2015, 197, 539–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yekkour, A.; Meklat, A.; Bijani, C.; Toumatia, O.; Errakhi, R.; Lebrihi, A.; Mathieu, F.; Zitouni, A.; Sabaou, N. A novel hydroxamic acid-containing antibiotic produced by a Saharan soil-living Streptomyces strain. Lett. Appl. Microbiol. 2015, 60, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, B.L.; Li, X.X.; Zhang, Z.B.; Yan, R.M.; Yang, H.L.; Zhu, D. Phylogenetic diversity of culturable endophytic fungi in Dongxiang wild rice (Oryza rufipogon Griff), detection of polyketide synthase gene and their antagonistic activity analysis. Fungal Biol. 2015, 119, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Hakvag, S.; Fjaervik, E.; Josefsen, K.D.; Ian, E.; Ellingsen, T.E.; Zotchev, S.B. Characterization of Streptomyces spp. isolated from the sea surface microlayer in the Trondheim Fjord, Norway. Mar. Drugs 2008, 6, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Abdelmohsen, U.R.; Yang, C.; Horn, H.; Hajjar, D.; Ravasi, T.; Hentschel, U. Actinomycetes from Red Sea sponges: Sources for chemical and phylogenetic diversity. Mar. Drugs 2014, 12, 2771–2789. [Google Scholar] [CrossRef] [PubMed]
- Bister, B.; Bischoff, D.; Ströbele, M.; Riedlinger, J.; Reicke, A.; Wolter, F.; Bull, A.T.; Zähner, H.; Fiedler, H.P.; Süssmuth, R.D. Abyssomicin C-A polycyclic antibiotic from a marine Verrucosispora strain as an inhibitor of the p-aminobenzoic acid/tetrahydrofolate biosynthesis pathway. Angew. Chem. Int. Ed. Engl. 2004, 43, 2574–2576. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, H.; Urai, M.; Ishii, K.; Yasukawa, J.; Paudel, A.; Murai, M.; Kaji, T.; Kuranaga, T.; Hamase, K.; Katsu, T.; et al. Lysocin E is a new antibiotic that targets menaquinone in the bacterial membrane. Nat. Chem. Biol. 2015, 11, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Abd, Y.E.; Awad, M. Evaporitic sediment distributions in Al-Kharrar sabkha, west Red Sea coast of Saudi Arabia, as revealed from electrical soundings. Mar. Geol. 1991, 97, 137–143. [Google Scholar] [CrossRef]
- Al-Washmi, H. Sedimentological Aspects and Environmental Conditions Recognized from the Bottom Sediments of Al-Kharrar Lagoon, Eastern Red Sea Coastal Plain, Saudi Arabia. J. KAU Mar. Sci. 1999, 10, 71–87. [Google Scholar] [CrossRef]
- Basaham, A.S. Mineralogical and chemical composition of the mud fraction from the surface sediments of Sharm Al-Kharrar, a Red Sea coastal lagoon. Oceanologia 2008, 50, 577–585. [Google Scholar]
- Gheith, A.M.; Abou Ouf, M.A. Textural characteristics, mineralogy and fauna in the shore zone sediments at Rabigh and Sharm al-Kharrar, eastern Red Sea, Saudi Arabia. J. KAU Mar. Sci. 1996, 7, 107–131. [Google Scholar] [CrossRef]
- Alzubaidy, H.; Essack, M.; Malas, T.B.; Bokhari, A.; Motwalli, O.; Kamanu, F.K.; Jamhor, S.A.; Mokhtar, N.A.; Antunes, A.; Simões, M.F.; et al. Rhizosphere microbiome metagenomics of gray mangroves (Avicennia marina) in the Red Sea. Gene 2016, 576, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Simoes, M.F.; Antunes, A.; Ottoni, C.A.; Amini, M.S.; Alam, I.; Alzubaidy, H.; Mokhtar, N.; Archer, J.A.C.; Bajic, V.B. Soil and Rhizosphere Associated Fungi in Gray Mangroves (Avicennia marina) from the Red Sea—A Metagenomic Approach. Genom. Proteom. Bioinform. 2015, 13, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Balk, M.; Keuskamp, J.A.; Laanbroek, H.J. Potential Activity, Size, and Structure of Sulfate-Reducing Microbial Communities in an Exposed, Grazed and a Sheltered, Non-Grazed Mangrove Stand at the Red Sea Coast. Front. Microbiol. 2015, 6, 1478. [Google Scholar] [CrossRef] [PubMed]
- El-Said, G.F.; Youssef, D.H. Ecotoxicological impact assessment of some heavy metals and their distribution in some fractions of mangrove sediments from Red Sea, Egypt. Environ. Monit. Assess. 2013, 185, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Kersten, R.D.; Nam, S.J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.; Dorrestein, P.C.; Moore, B.S.; Qian, P.Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.; Esau, L.; Hikmawan, T.; Antunes, A.; Holtermann, K.; Stingl, U.; Bajic, V.B.; Kaur, M. Cytotoxic and apoptotic evaluations of marine bacteria isolated from brine-seawater interface of the Red Sea. BMC Complement. Altern. Med. 2013, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Nawwar, M.; Hussein, S.; Ayoub, N.A.; Hashim, A.; Mernitz, G.; Cuypers, B.; Linscheid, M.; Lindequist, U. Deuteromycols A and B, two benzofuranoids from a Red Sea marine-derived Deuteromycete sp. Arch. Pharm. Res. 2010, 33, 1729–1733. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Rakhisi, Z.; Ahmady, A.Z. Isolation and Identification of Bacillus Species from Soil and Evaluation of Their Antibacterial Properties. Avicenna J. Clin. Microbiol. Infect. 2015, 2. [Google Scholar] [CrossRef]
- Shaligram, N.S.; Singhal, R.S. Surfactin—A review on biosynthesis, fermentation, purification and applications. Food Technol. Biotechnol. 2010, 48, 119–134. [Google Scholar]
- Schneider, K.; Chen, X.H.; Vater, J.; Franke, P.; Nicholson, G.; Borriss, R.; Süssmuth, R.D. Macrolactin is the polyketide biosynthesis product of the pks2 cluster of Bacillus amyloliquefaciens FZB42. J. Nat. Prod. 2007, 70, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New drugs from marine organisms in Alzheimer’s Disease. Mar. Drugs 2015, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Lombó, F.; Velasco, A.; Castro, A.; de la Calle, F.; Braña, A.F.; Sánchez-Puelles, J.M.; Méndez, C.; Salas, J.A. Deciphering the biosynthesis pathway of the antitumor thiocoraline from a marine actinomycete and its expression in two Streptomyces species. ChemBioChem 2006, 7, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular networking as a dereplication strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Della Sala, G.; Hochmuth, T.; Teta, R.; Costantino, V.; Mangoni, A. Polyketide synthases in the microbiome of the marine sponge Plakortis halichondrioides: A metagenomic update. Mar. Drugs 2014, 12, 5425–5440. [Google Scholar] [CrossRef] [PubMed]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Rappe, M.S.; Giovannoni, S.J. The uncultured microbial majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Zengler, K. Tapping into microbial diversity. Nat. Rev. Microbiol. 2004, 2, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Borneman, J.; Triplett, E.W. Molecular microbial diversity in soils from eastern Amazonia: Evidence for unusual microorganisms and microbial population shifts associated with deforestation. Appl. Environ. Microbiol. 1997, 63, 2647–2653. [Google Scholar] [PubMed]
- Ohtonen, R.; Fritze, H.; Pennanen, T.; Jumpponen, A.; Trappe, J. Ecosystem properties and microbial community changes in primary succession on a glacier forefront. Oecologia 1999, 119, 239–246. [Google Scholar] [CrossRef]
- Davis, K.E.; Joseph, S.J.; Janssen, P.H. Effects of growth medium, inoculum size, and incubation time on culturability and isolation of soil bacteria. Appl. Environ. Microbiol. 2005, 71, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.-I. Establishment and use of gellan gum media for selective isolation and distribution survey of specific rare actinomycetes. Actinomycetologica 2001, 15, 55–60. [Google Scholar] [CrossRef]
- Stingl, U.; Cho, J.C.; Foo, W.; Vergin, K.L.; Lanoil, B.; Giovannoni, S.J. Dilution-to-extinction culturing of psychrotolerant planktonic bacteria from permanently ice-covered lakes in the McMurdo Dry Valleys, Antarctica. Microb. Ecol. 2008, 55, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Letelier, A.; Olmedo, G.; Eguiarte, L.E.; Martinez-Castilla, L.; Souza, V. Parallel evolution and horizontal gene transfer of the pst operon in Firmicutes from oligotrophic environments. Int. J. Evol. Biol. 2011, 2011, 781642. [Google Scholar] [CrossRef] [PubMed]
- Stolz, J.; Basu, P.; Oremland, R. Microbial transformation of elements: The case of arsenic and selenium. Int. Microbiol. 2002, 5, 201–207. [Google Scholar] [PubMed]
- Aislabie, J.; Deslippe, J.; Dymond, J. Soil Microbes and Their Contribution to Soil Services. In Ecosystem Services in New Zealand: Conditions and Trends; Manaaki Whenua Press: Lincoln, New Zealand, 2013; pp. 143–161. [Google Scholar]
- Chen, X.-H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W.; et al. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB 42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.S.; Priest, F.; Diderichsen, B. On the industrial use of Bacillus licheniformis: A review. Appl. Microbiol. Biotechnol. 1994, 40, 595–598. [Google Scholar] [CrossRef]
- Wayne, L.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef]
- Johnson, J. Bacterial classification III. Nucleic acids in bacterial classification. In Bergey's Manual of Systematic Bacteriology; Williams & Wilkins: Baltimore, MD, USA, 1984; pp. 8–11. [Google Scholar]
- Stackebrandt, E.; Goebel, B. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Boone, D.R.; Whitman, W.B.; Rouvière, P. Diversity and Taxonomy of Methanogens. In Methanogenesis; Springer: New York, NY, USA, 1993; pp. 35–80. [Google Scholar]
- Bhatnagar, I.; Kim, S.-K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef] [PubMed]
- Rusin, P.A.; Maxwell, S.L.; Brooks, J.P.; Gerba, C.P.; Pepper, I.L. Evidence for the absence of Staphylococcus aureus in land applied biosolids. Environ. Sci. Technol. 2003, 37, 4027–4030. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Maybank, R.A.; Enke, S.; Friss, M.B.; Diviak, L.F.; Karaolis, D.K.; Koren, S.; Ondov, B.; Phillippy, A.M.; Bergman, N.H.; et al. Complete genome sequence of the quality control strain Staphylococcus aureus subsp. aureus ATCC 25923. Genome Announc. 2014, 2, e01110-14. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.-F.; He, S.Y. Pseudomonas syringae pv. tomato DC3000: A model pathogen for probing disease susceptibility and hormone signaling in plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Kay, S.; Connor, T.; Barquist, L.; Sait, L.; Holt, K.E.; Sivaraman, K.; Wileman, T.; Goulding, D.; Clare, S.; et al. Genome and transcriptome adaptation accompanying emergence of the definitive type 2 host-restricted Salmonella enterica serovar Typhimurium pathovar. MBio 2013, 4, e00565-13. [Google Scholar] [CrossRef] [PubMed]
- Rautenbach, M.; Eyéghé-Bickong, H.A.; Vlok, N.M.; Stander, M.; de Beer, A. Direct surfactin-gramicidin S antagonism supports detoxification in mixed producer cultures of Bacillus subtilis and Aneurinibacillus migulanus. Microbiology 2012, 158, 3072–3082. [Google Scholar] [CrossRef] [PubMed]
- Gause, G.F.; Brazhnikova, M.G. Gramicidin S and its use in the Treatment of Infected Wounds. Nature 1944, 154, 703. [Google Scholar] [CrossRef]
- Veith, B.; Herzberg, C.; Steckel, S.; Feesche, J.; Maurer, K.H.; Ehrenreich, P.; Bäumer, S.; Henne, A.; Liesegang, H.; Merkl, R.; et al. The complete genome sequence of Bacillus licheniformis DSM13, an organism with great industrial potential. J. Mol. Microbiol. Biotechnol. 2004, 7, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Ordonez, A.; Begley, M.; Clifford, T.; Deasy, T.; Considine, K.; O’Connor, P.; Ross, R.P.; Hill, C. Investigation of the antimicrobial activity of Bacillus licheniformis strains isolated from retail powdered infant milk formulae. Probiot. Antimicrob. Proteins 2014, 6, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Todorova, S.; Kozhuharova, L. Characteristics and antimicrobial activity of Bacillus subtilis strains isolated from soil. World J. Microbiol. Biotechnol. 2010, 26, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Pandya, U.; Saraf, M. Isolation and identification of allelochemicals produced by B. sonorensis for suppression of charcoal rot of Arachis hypogaea L. J. Basic Microbiol. 2015, 55, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Chopra, L.; Singh, G.; Jena, K.K.; Verma, H.; Sahoo, D.K. Bioprocess. development for the production of sonorensin by Bacillus sonorensis MT93 and its application as a food preservative. Bioresour. Technol. 2014, 175, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, Q.; Wang, K.; Brian, K.; Liu, C.; Gu, Y. Study of the antifungal activity of Bacillus vallismortis ZZ185 in vitro and identification of its antifungal components. Bioresour. Technol. 2010, 101, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Sacido, A.; Genilloud, O. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: Detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 2005, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.-K. Pharmaceutically active secondary metabolites of marine actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Salomon, C.E.; Magarvey, N.A.; Sherman, D.H. Merging the potential of microbial genetics with biological and chemical diversity: An even brighter future for marine natural product drug discovery. Nat. Prod. Rep. 2004, 21, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.H.; Song, J.J.; Lee, S.G.; Kwon, S.J.; Asano, Y.; Sung, M.H. New thermostable d-methionine amidase from Brevibacillus borstelensis BCS-1 and its application for d-phenylalanine production. Enzym. Microb. Technol. 2003, 32, 131–139. [Google Scholar] [CrossRef]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [PubMed]
- Ausubel, F.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. (Eds.) Short Protocols in Molecular Biology; John Wiley & Sons: New York, NY, USA, 1995; pp. 2–3.
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhu, K.; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Fuerst, J.A. Diversity of polyketide synthase genes from bacteria associated with the marine sponge Pseudoceratina clavata: Culture-dependent and culture-independent approaches. Environ. Microbiol. 2006, 8, 1460–1470. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, A.; Clark, D.; González, I.; Salazar, O.; Anderson, A.; Genilloud, O. A novel actinomycete strain de-replication approach based on the diversity of polyketide synthase and nonribosomal peptide synthetase biosynthetic pathways. Appl. Microbiol. Biotechnol. 2005, 67, 795–806. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Enzyme | Source | Activity | Reference |
|---|---|---|---|---|
| Bacitracin | NRPS | Bacillus spp. | Antibacterial | [26] |
| Surfactin | NRPS | Bacillus subtilis | Antibacterial | [27] |
| Macrolactin | PKS | Bacillus amyloliquefaciens | Antibacterial | [28] |
| Mupirocin | PKS | Pseudomonads fluorescens | Antibacterial | [29] |
| Retimycin | NRPS | Sallinispora arenicola | Antitumor | [30] |
| Pederin | PKS | Paederus fuscipes | Antitumor | [31] |
| Salinosporamide K | NRPS | Salinispora pacifica | Antitumor | [32] |
| Salinilactam A | PKS | Salinispora tropica | Antitumor | [33] |
| Bryostatin | PKS | Candidatus Endobugula sertula | Antitumor | [34] |
| Strain ID | % Similarity of 16S rRNA Sequences to GenBank Strains: | Number of Nucleotides (bp) | Phylum | Location | Sediment Type | Media |
|---|---|---|---|---|---|---|
| Bac34 | 98% Vibrio alginolyticus | 1429 | Proteobacteria | RHL | MN | 10% MA |
| Bac56 | 98% Microbulbifer maritimus | 1390 | Proteobacteria | RHL | MN | 10% MA |
| Bac77 | 98% Pseudoalteromonas flavipulchra | 1406 | Proteobacteria | RHL | MM | 10% MA |
| Bac85 | 95% Microbulbifer gwangyangensis | 1406 | Proteobacteria | RHL | MM | 10% MA |
| Bac92 | 97% Planctomyces brasiliensis | 1372 | Planctomycete | RHL | MM | Anti-MA |
| Bac94 | 98% Bacillus simplex | 1423 | Firmicutes | RHL | MM | 10% MA |
| Bac120 | 98% Oceanicaulis sp. | 1336 | Proteobacteria | RHL | BS | 10% MA |
| Bac131 | 98% Planctomycete sp. | 1417 | Planctomycete | RHL | BS | Anti-MA |
| Bac140 | 97% Flavobacteriaceae bacterium | 1394 | Bacteroidetes | RHL | BS | 10% MA |
| Bac175 | 98% Blastopirellula cremea | 1385 | Planctomycete | AKL | MN | Anti-MA |
| Bac181 | 91% Brevibacterium avium | 1341 | Actinobacteria | AKL | MN | AIA |
| Bac216 | 97% Marinobacter xestospongiae | 1414 | Proteobacteria | AKL | MM | 10% MA |
| Bac319 | 98% Pseudoalteromonas espejiana | 1380 | Proteobacteria | RHL | MM | 10% MA |
| Bac320 | 98% Pseudoalteromonas atlantica | 1392 | Proteobacteria | RHL | MM | MB-GM |
| Bac387 | 96% Exiguobacterium profundum | 1327 | Firmicutes | RHL | MN | 10% MA |
| Closest Phylogenetic Relative by BLAST | Sequence Similarity (%) | Zone of Inhibition (mm) | |||||
|---|---|---|---|---|---|---|---|
| Staphylococcus aureus | Salmonella typhimurium | Pseudomonas syringae | |||||
| Annular radius | Diameter | Annular radius | Diameter | Annular radius | Diameter | ||
| Bacillus licheniformis | 99 | 4.5 | 14 | 3 | 11 | 3 | 11 |
| Bacillus sonorensis | 99 | 3.8 | 12 | 2.8 | 10 | 3 | 11 |
| Brevibacillus borstelensis | 99 | 5 | 15 | 0.5 | 6 | 4.3 | 13.5 |
| Bacillus vallismortis | 99 | 2 | 9 | 4 | 13 | 0.5 | 6 |
| Microbulbifer salipaludis | 99 | 3.3 | 11 | 6.5 | 18 | 1 | 7 |
| Bacillus subtilis | 99 | 5 | 15 | 3 | 11 | 0.5 | 6 |
| Aneurinibacillus migulanus | 99 | 4 | 13 | 2 | 9 | 3.3 | 11.5 |
| Aneurinibacillus migulanus | 99 | 5.5 | 16 | 3 | 11 | 4.5 | 14 |
| Paenibacillus dendritiformis | 99 | 0.5 | 6 | 3 | 11 | 1 | 7 |
| Paenibacillus dendritiformis | 99 | 2 | 9 | 2 | 6 | 5 | 15 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Amoudi, S.; Essack, M.; Simões, M.F.; Bougouffa, S.; Soloviev, I.; Archer, J.A.C.; Lafi, F.F.; Bajic, V.B. Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential. Mar. Drugs 2016, 14, 165. https://doi.org/10.3390/md14090165
Al-Amoudi S, Essack M, Simões MF, Bougouffa S, Soloviev I, Archer JAC, Lafi FF, Bajic VB. Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential. Marine Drugs. 2016; 14(9):165. https://doi.org/10.3390/md14090165
Chicago/Turabian StyleAl-Amoudi, Soha, Magbubah Essack, Marta F. Simões, Salim Bougouffa, Irina Soloviev, John A. C. Archer, Feras F. Lafi, and Vladimir B. Bajic. 2016. "Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential" Marine Drugs 14, no. 9: 165. https://doi.org/10.3390/md14090165
APA StyleAl-Amoudi, S., Essack, M., Simões, M. F., Bougouffa, S., Soloviev, I., Archer, J. A. C., Lafi, F. F., & Bajic, V. B. (2016). Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential. Marine Drugs, 14(9), 165. https://doi.org/10.3390/md14090165