Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo

 ,
, 
Abstract
:
1. Introduction
2. Results
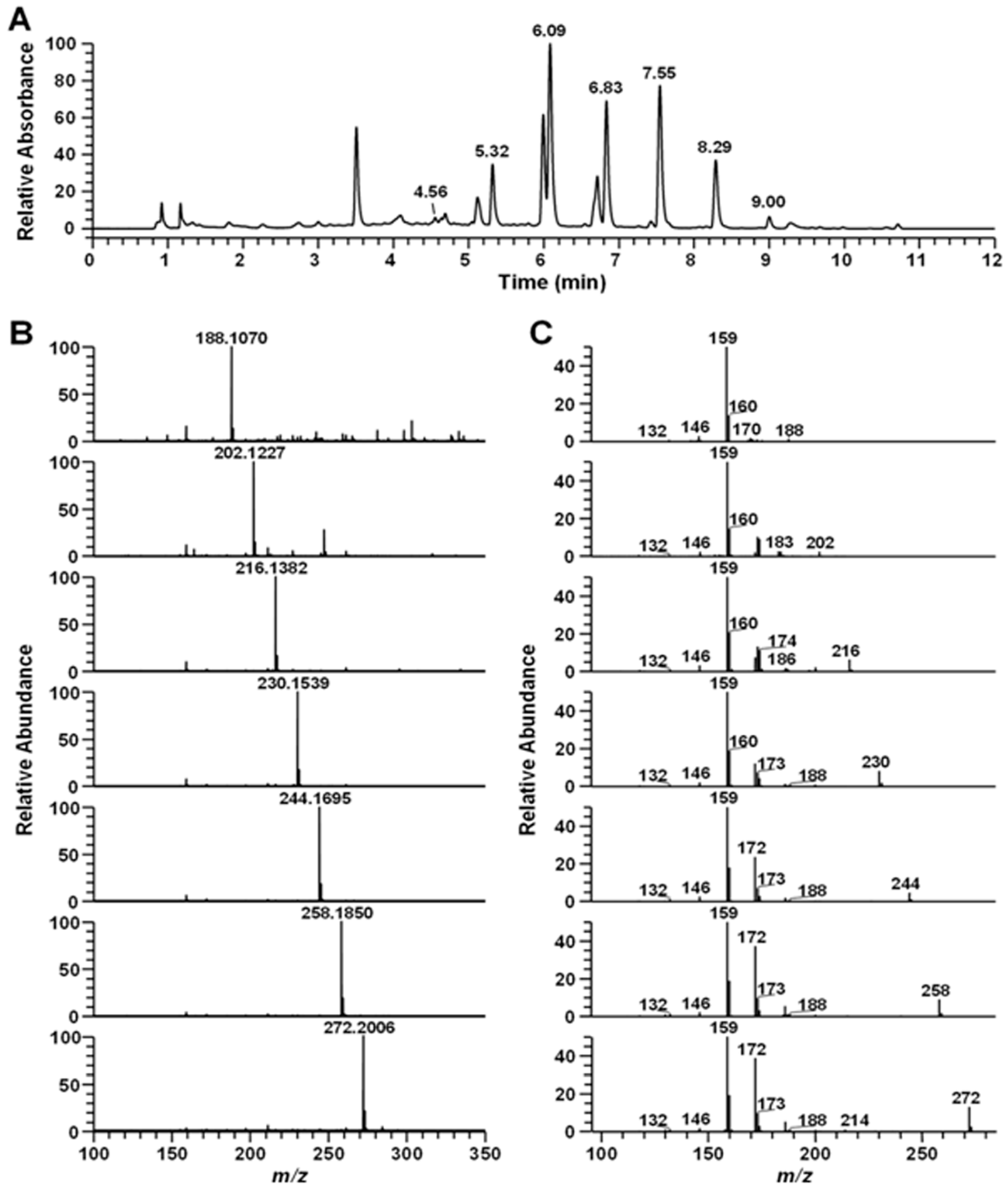
2.1. LC-MS Analysis of the Secondary Metabolite
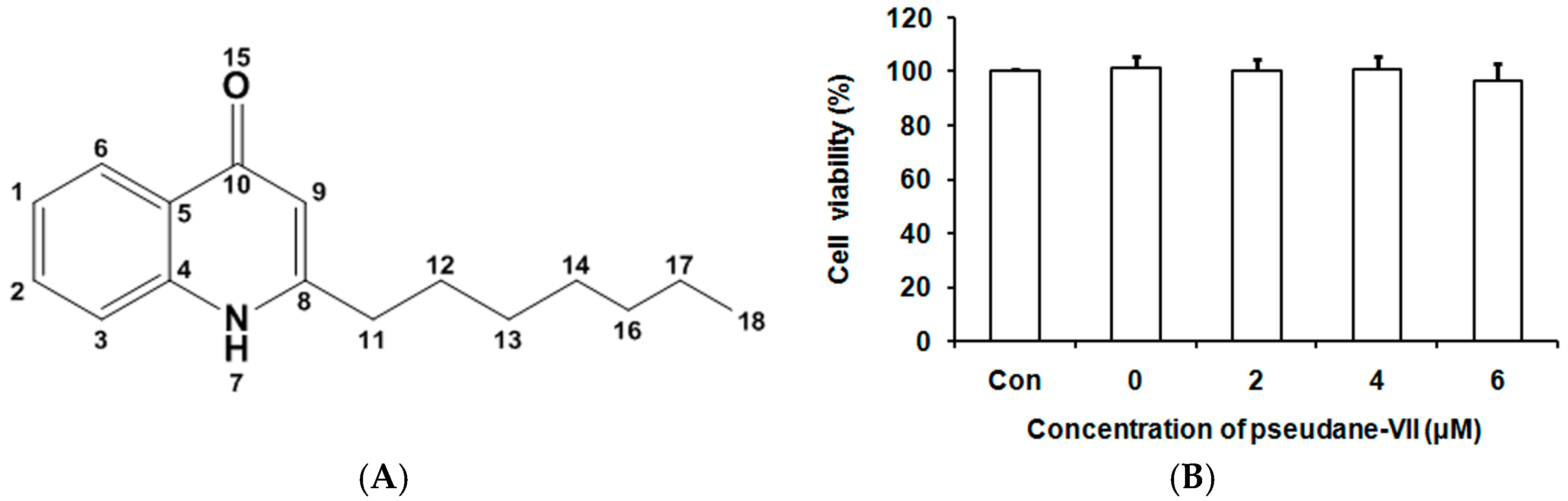
2.2. Purification and Analysis of Pseudane VII
2.3. Pseudane-VII Inhibited NO Production in LPS-Stimulated Macrophages
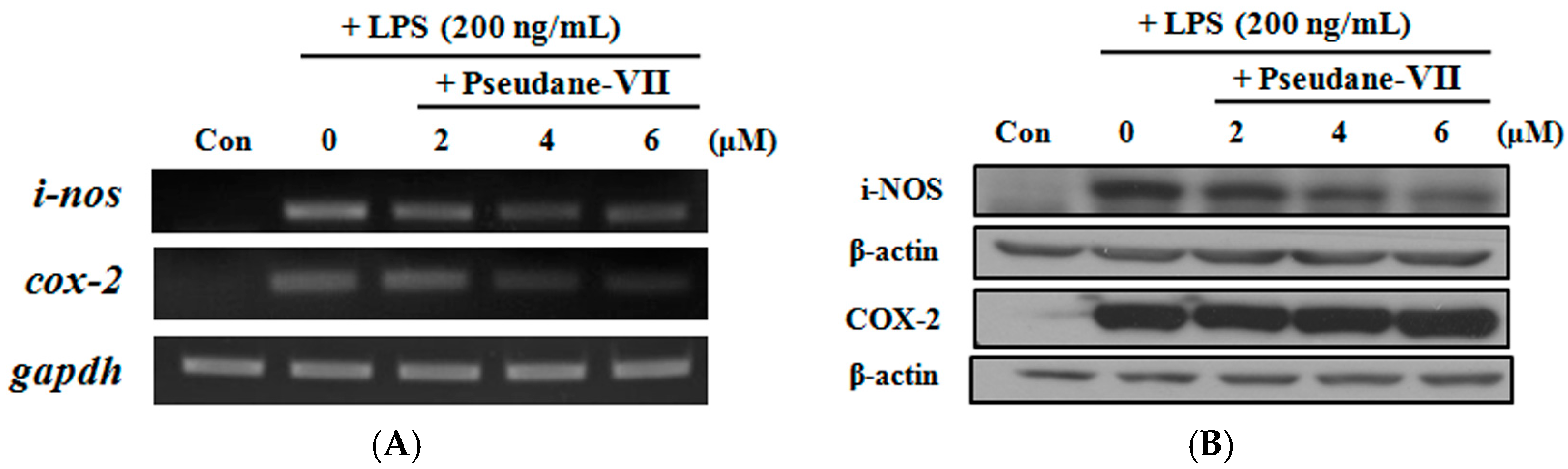
2.4. Pseudane-VII Suppressed Pro-Inflammatory Enzyme Expression
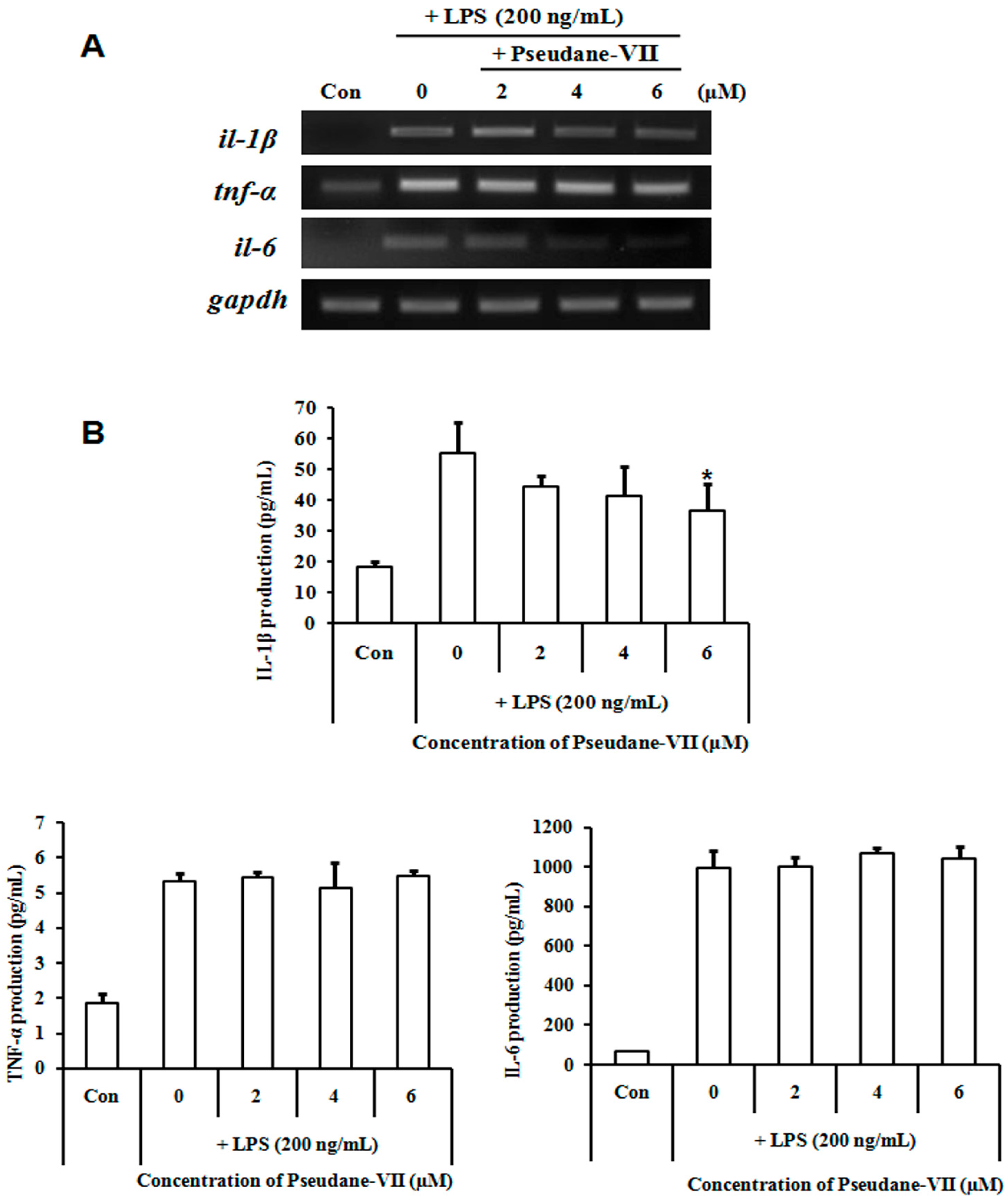
2.5. Pseudane-VII Diminished Pro-Inflammatory Cytokine Production
2.6. Pseudane-VII Inhibited LPS-Induced MAPK Activation and NF-κB Signaling
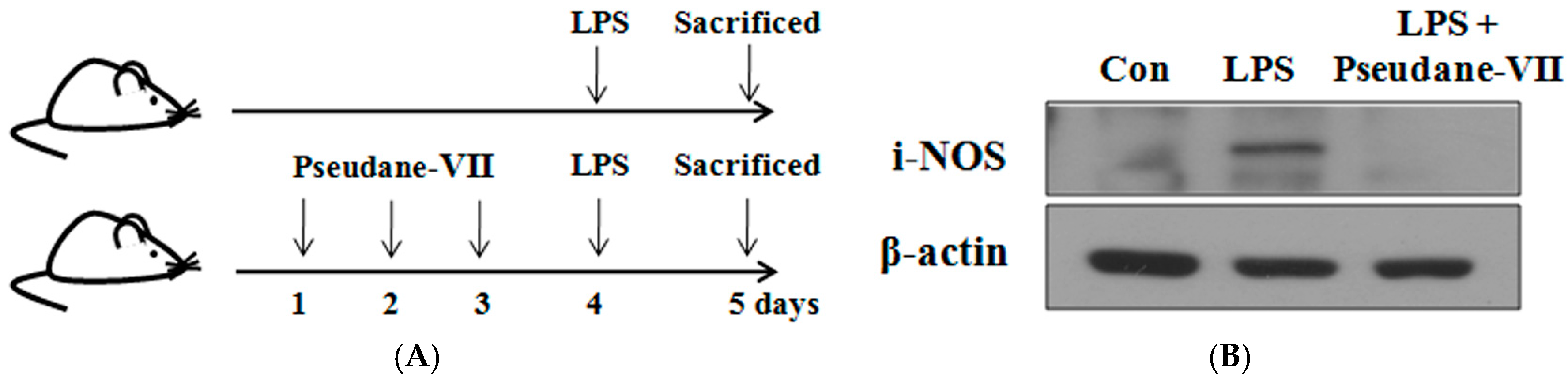
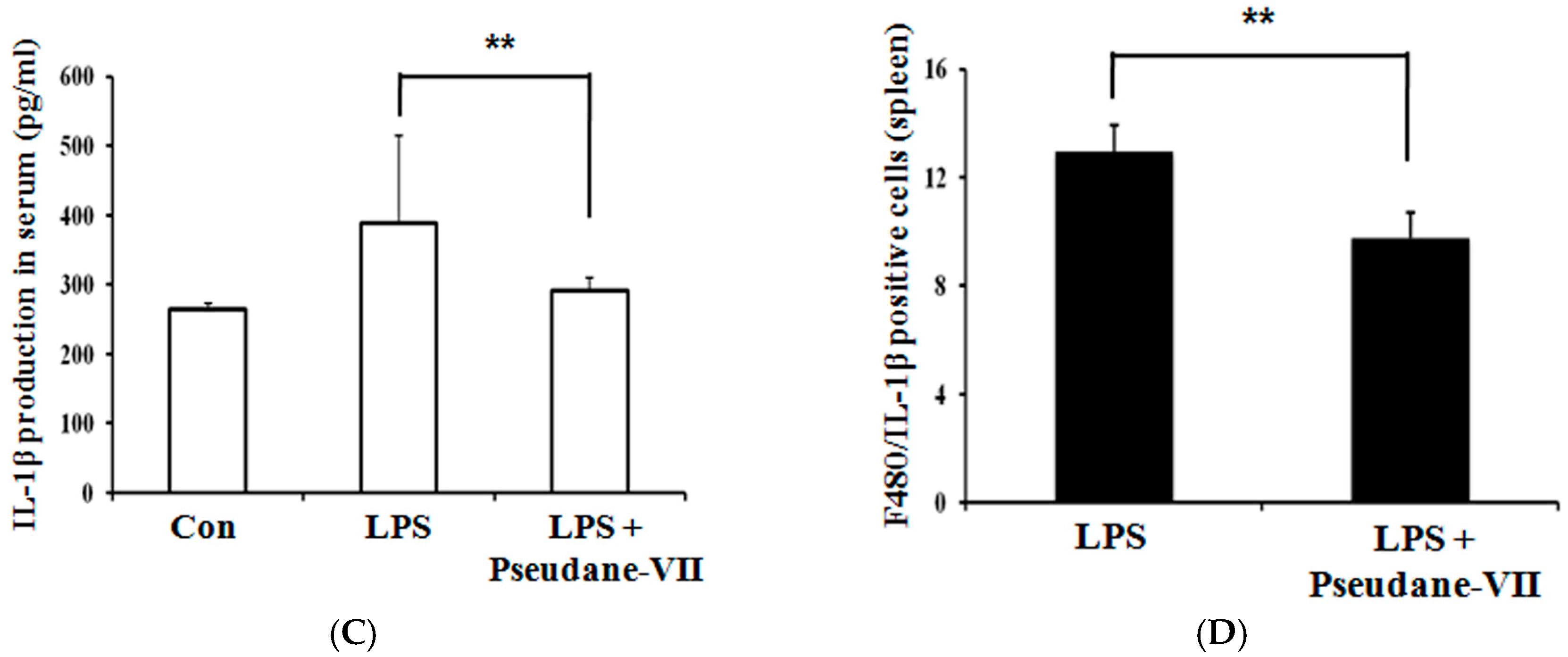
2.7. Pseudane-VII Decreased iNOS Expression and IL-1β Production in LPS-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Extraction and Purification of Secondary Metabolites
4.3. LC-MS Analysis of A5 (Pseudane VII)
4.4. Nuclear Magnetic Resonance (NMR) Analysis
4.5. Cell Culture and Pseudane-VII Treatment
4.6. Cytotoxicity Assay
4.7. NO Assay
4.8. Reverse Transcription (RT)-PCR
4.9. Western Blot Analysis
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Isolation of IL-1β Producing-Macrophages from Spleen
4.12. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mitchell, G.; Isberg, R.R. Innate Immunity to Intracellular Pathogens: Balancing Microbial Elimination and Inflammation. Cell Host Microbe 2017, 22, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Manthiram, K.; Zhou, Q.; Aksentijevich, I.; Kastner, D.L. The monogenic autoinflammatory diseases define new pathways in human innate immunity and inflammation. Nat. Immunol. 2017, 18, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Choi, J.Y.; Lee, J.M.; Lee, S.; Cho, E.J. Tartary buckwheat on nitric oxide-induced inflammation in RAW264.7 macrophage cells. Food Funct. 2015, 6, 2664–2670. [Google Scholar] [CrossRef] [PubMed]
- Bombicino, S.S.; Iglesias, D.E.; Rukavina-Mikusic, I.A.; Buchholz, B.; Gelpi, R.J.; Boveris, A.; Valdez, L.B. Hydrogen peroxide, nitric oxide and ATP are molecules involved in cardiac mitochondrial biogenesis in Diabetes. Free Radic. Biol. Med. 2017, 112, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Hong, C.; Wang, Y.; Liu, J.; Zhang, N.; Shen, C.; Wei, W.; Zheng, F. Calreticulin promotes angiogenesis via activating nitric oxide signalling pathway in rheumatoid arthritis. Clin. Exp. Immunol. 2014, 178, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Sun, K.H.; Chiang, C.P.; Huang, C.F.; Sun, G.H.; Tsou, Y.C.; Liu, H.Y.; Tang, S.J. Nitric oxide suppresses LPS-induced inflammation in a mouse asthma model by attenuating the interaction of IKK and Hsp90. Exp. Biol. Med. 2015, 240, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Lee, C.H.; Chang, Y.W.; Wang, H.M.; Chen, C.Y.; Chen, Y.H. Pheophytin a inhibits inflammation via suppression of LPS-induced nitric oxide synthase-2, prostaglandin E2, and interleukin-1β of macrophages. Int. J. Mol. Sci. 2014, 15, 22819–22834. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Z.; Jiang, Z.K.; He, B.X.; Liu, X.B. Arctigenin Protects against Lipopolysaccharide-Induced Pulmonary Oxidative Stress and Inflammation in a Mouse Model via Suppression of MAPK, HO-1, and iNOS Signaling. Inflammation 2015, 38, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Merighi, S.; Bencivenni, S.; Vincenzi, F.; Varani, K.; Borea, P.A.; Gessi, S. A2B adenosine receptors stimulate IL-6 production in primary murine microglia through p38 MAPK kinase pathway. Pharmacol. Res. 2017, 117, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Lu, Y.; Zhao, C.; Gao, W.; He, F.; Zhang, J.; Zhao, D.; Qiu, W.; Wang, Y. C5a Induces the Synthesis of IL-6 and TNF-alpha in Rat Glomerular Mesangial Cells through MAPK Signaling Pathways. PLoS ONE 2016, 11, e0161867. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.; Gauthier, M.; Christen, R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. Int. J. Syst. Bacteriol. 1995, 45, 755–761. [Google Scholar] [PubMed]
- Kim, W.J.; Kim, Y.O.; Kim, J.H.; Nam, B.H.; Kim, D.G.; An, C.M.; Lee, J.S.; Kim, P.S.; Lee, H.M.; Oh, J.S. Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2. Mar. Drugs 2016, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, T.; Honda, M.; Oba, T.; Ozawa, K. ER stress upregulated PGE(2)/IFNgamma-induced IL-6 expression and down-regulated iNOS expression in glial cells. Sci. Rep. 2013, 3, 3388. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.H.; Kim, I.K.; Lee, H.I.; Joo, H.; Lim, J.U.; Lee, J.; Lee, S.H.; Moon, H.S. Chronic intermittent hypoxia induces liver fibrosis in mice with diet-induced obesity via TLR4/MyD88/MAPK/NF-kB signaling pathways. Biochem. Biophys. Res. Commun. 2017, 490, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Li, W.; Zhang, Y.; Wu, X.; Song, Y.; Kang, L.; Liu, W.; Wang, K.; Li, S.; Hua, W.; et al. Simvastatin Inhibits IL-1β-Induced Apoptosis and Extracellular Matrix Degradation by Suppressing the NF-kB and MAPK Pathways in Nucleus Pulposus Cells. Inflammation 2017, 40, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, Y.; Yao, X.; Hao, F.; Yu, C.; Bao, Y.; Song, Z.; Sun, Y.; Zheng, L.; Wang, G.; et al. Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 794. [Google Scholar] [CrossRef] [PubMed]
- Boniakowski, A.E.; Kimball, A.S.; Jacobs, B.N.; Kunkel, S.L.; Gallagher, K.A. Macrophage-Mediated Inflammation in Normal and Diabetic Wound Healing. J. Immunol. 2017, 199, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Q.; Dai, Y.; Yang, Y.; Huang, C.; Meng, X.M.; Wu, B.M.; Li, J. Emerging role of microRNAs in regulating macrophage activation and polarization in immune response and inflammation. Immunology 2016, 148, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Dunster, J.L. The macrophage and its role in inflammation and tissue repair: Mathematical and systems biology approaches. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Morrison, M.C.; Kleemann, R. Role of Macrophage Migration Inhibitory Factor in Obesity, Insulin Resistance, Type 2 Diabetes, and Associated Hepatic Co-Morbidities: A Comprehensive Review of Human and Rodent Studies. Front. Immunol. 2015, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Bornfeldt, K.E. Macrophage Phenotype and Function in Different Stages of Atherosclerosis. Circ. Res. 2016, 118, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Dehne, N.; Mora, J.; Namgaladze, D.; Weigert, A.; Brune, B. Cancer cell and macrophage cross-talk in the tumor microenvironment. Curr. Opin. Pharmacol. 2017, 35, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Binnebosel, M.; Rosch, R.; Junge, K.; Lynen-Jansen, P.; Schumpelick, V.; Klinge, U. Macrophage and T-lymphocyte infiltrates in human peritoneal adhesions indicate a chronic inflammatory disease. World J. Surg. 2008, 32, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Landriscina, A.; Rosen, J.M.; Wei, G.; Kao, S.; Olcott, W.; Agak, G.W.; Paz, K.B.; Bonventre, J.; Clendaniel, A.; et al. Nitric Oxide-Releasing Nanoparticles Prevent Propionibacterium Acnes-Induced Inflammation by Both Clearing the Organism and Inhibiting Microbial Stimulation of the Innate Immune Response. J. Investig. Dermatol. 2015, 135, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Madhu, B.P.; Singh, K.P.; Saminathan, M.; Singh, R.; Shivasharanappa, N.; Sharma, A.K.; Malik, Y.S.; Dhama, K.; Manjunatha, V. Role of nitric oxide in the regulation of immune responses during rabies virus infection in mice. Virusdisease 2016, 27, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ren, X.; Zhao, M.; Zhou, B.; Han, Y. Angiotensin-(1-7) abrogates angiotensin II-induced proliferation, migration and inflammation in VSMCs through inactivation of ROS-mediated PI3K/Akt and MAPK/ERK signaling pathways. Sci. Rep. 2016, 6, 34621. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. C/H | δC ppm | δH (ppm), Integration, Multiplicity, J (Hz) |
|---|---|---|
| 1(CH) | 123.6 | 7.33 (1H, t, J = 8.05, 7.61 Hz) |
| 2(CH) | 131.8 | 7.58 (1H, t, J = 7.96, 7.61 Hz) |
| 3(CH) | 118.3 | 7.71 (1H, d, J = 7.96 Hz) |
| 4(C) | 140.5 | - |
| 5(C) | 125.0 | - |
| 6(CH) | 125.4 | 8.36 (1H, d, J = 8.05 Hz) |
| 8(C) | 154.9 | - |
| 9(CH) | 108.3 | 6.24 (1H, s) |
| 10(C) | 178.9 | - |
| 11(CH2) | 34.4 | 2.68 (2H, t, J = 7.50 Hz) |
| 12(CH2) | 28.5 | 1.72 (2H, tt, J = 7.50, 6.60 Hz) |
| 13(CH2) | 29.5 | 1.28 (2H, tt, J = 7.54, 6.60 Hz) |
| 14(CH2) | 28.7 | 1.10 (2H, quin, J = 7.54, 7.40 Hz) |
| 16(CH2) | 30.8 | 1.19 (2H, quin, J = 7.40 Hz) |
| 17(CH2) | 21.9 | 1.28 (2H, tt, J = 7.54, 6.60 Hz) |
| 18(CH3) | 13.8 | 0.87 (3H, t, J = 6.77 Hz) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.E.; Jung, I.; Lee, J.S.; Na, J.Y.; Kim, W.J.; Kim, Y.-O.; Park, Y.-D.; Lee, J.S. Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Mar. Drugs 2017, 15, 336. https://doi.org/10.3390/md15110336
Kim ME, Jung I, Lee JS, Na JY, Kim WJ, Kim Y-O, Park Y-D, Lee JS. Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Marine Drugs. 2017; 15(11):336. https://doi.org/10.3390/md15110336
Chicago/Turabian StyleKim, Mi Eun, Inae Jung, Jong Suk Lee, Ju Yong Na, Woo Jung Kim, Young-Ok Kim, Yong-Duk Park, and Jun Sik Lee. 2017. "Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo" Marine Drugs 15, no. 11: 336. https://doi.org/10.3390/md15110336
APA StyleKim, M. E., Jung, I., Lee, J. S., Na, J. Y., Kim, W. J., Kim, Y. -O., Park, Y. -D., & Lee, J. S. (2017). Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Marine Drugs, 15(11), 336. https://doi.org/10.3390/md15110336