AS1041, a Novel Synthesized Derivative of Marine Natural Compound Aspergiolide A, Arrests Cell Cycle, Induces Apoptosis, and Inhibits ERK Activation in K562 Cells


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
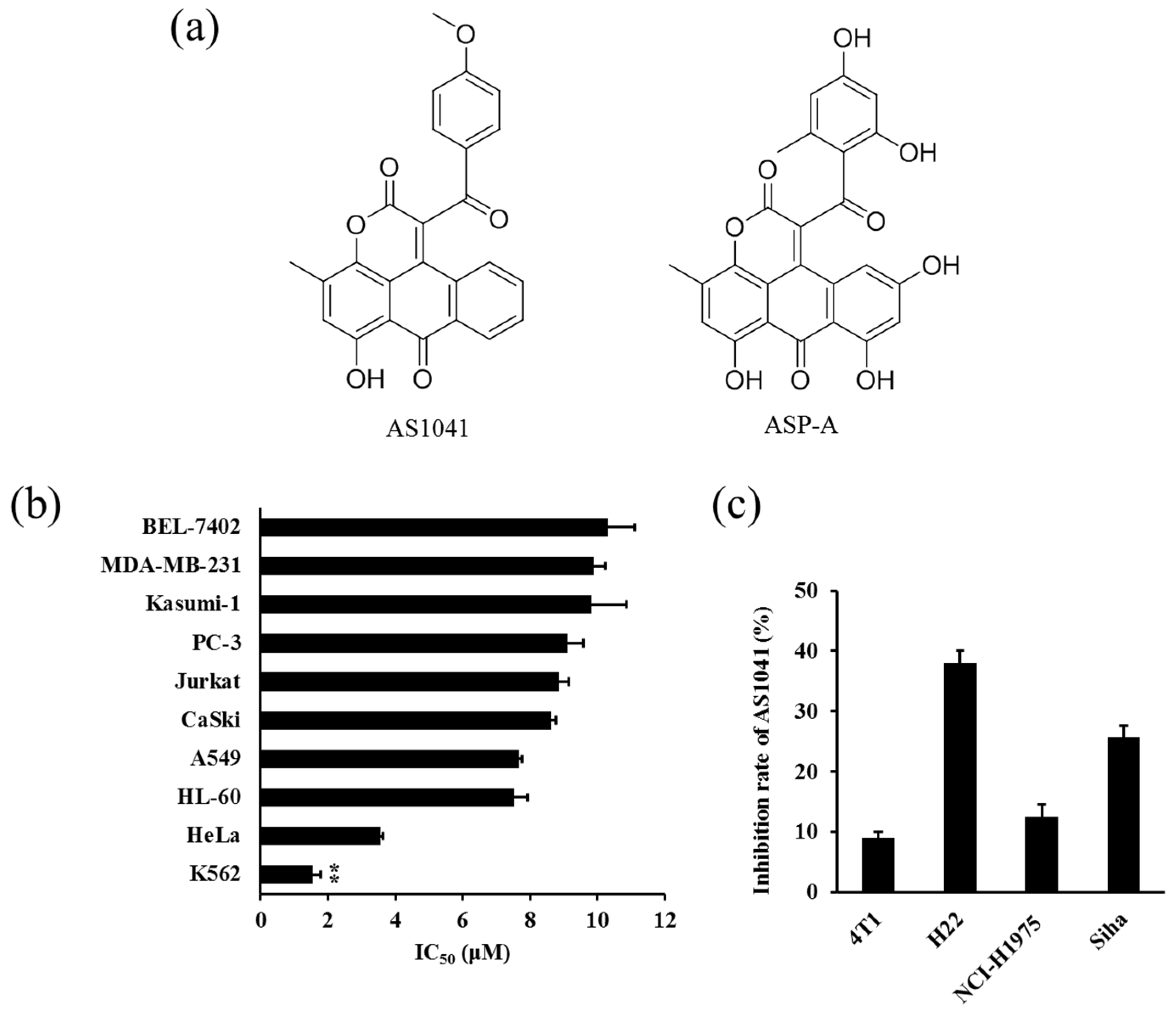
2.1. Anticancer Spectrum of AS1041
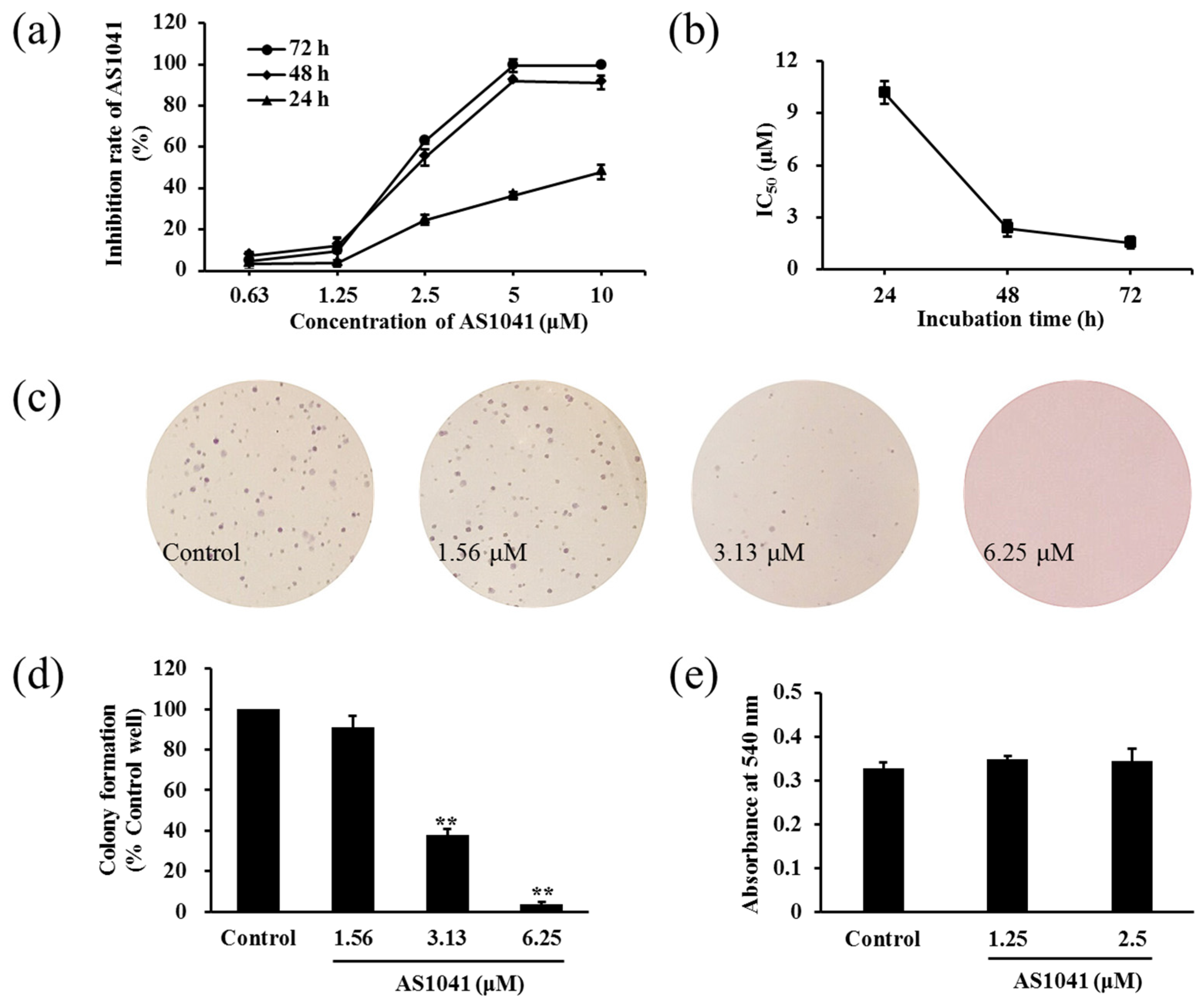
2.2. AS1041 Inhibits the Proliferation of K562 Cells
2.3. AS1041 Induces Cell Cycle Arrest at G2/M Phase in K562 Cells
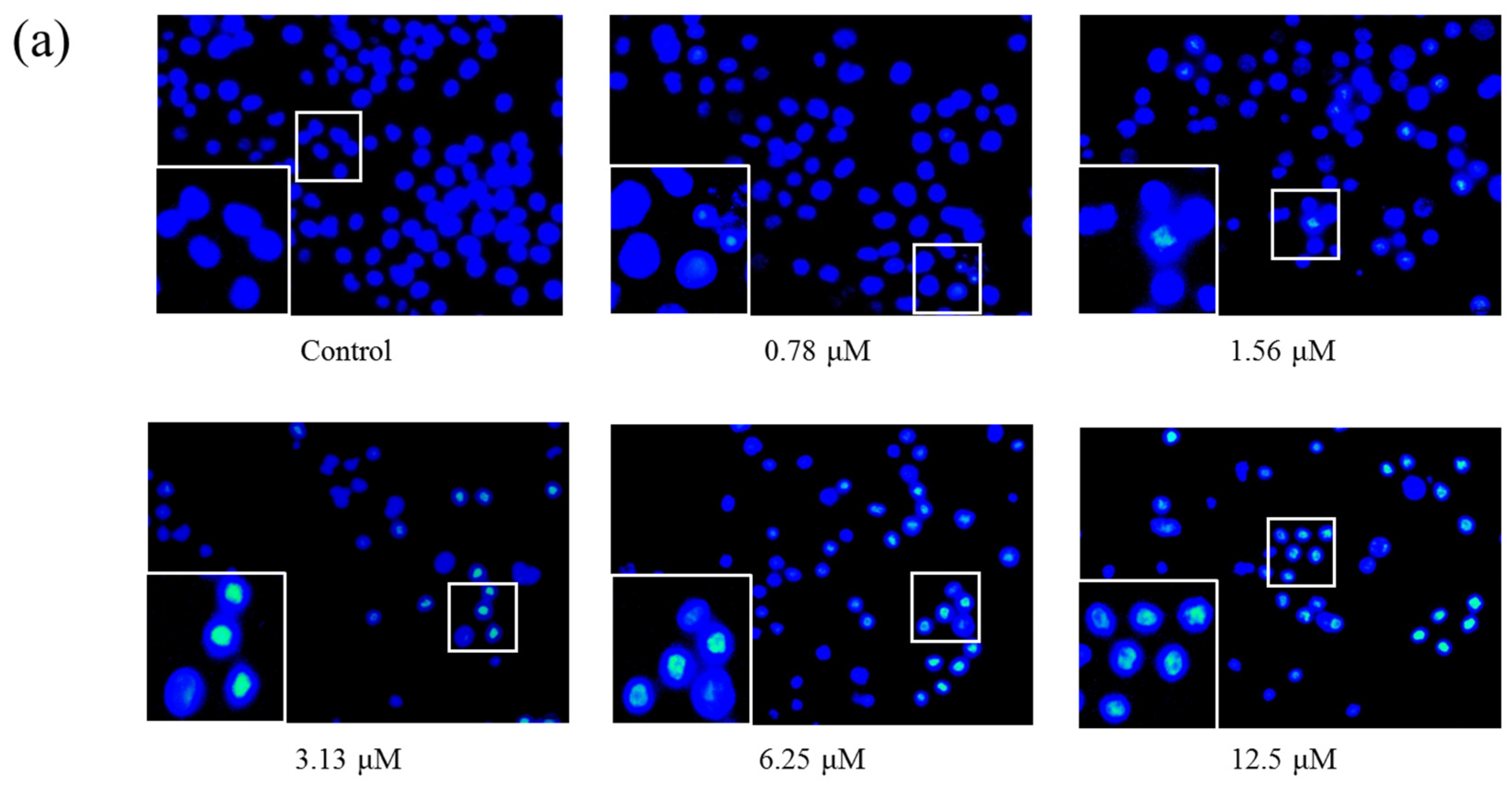
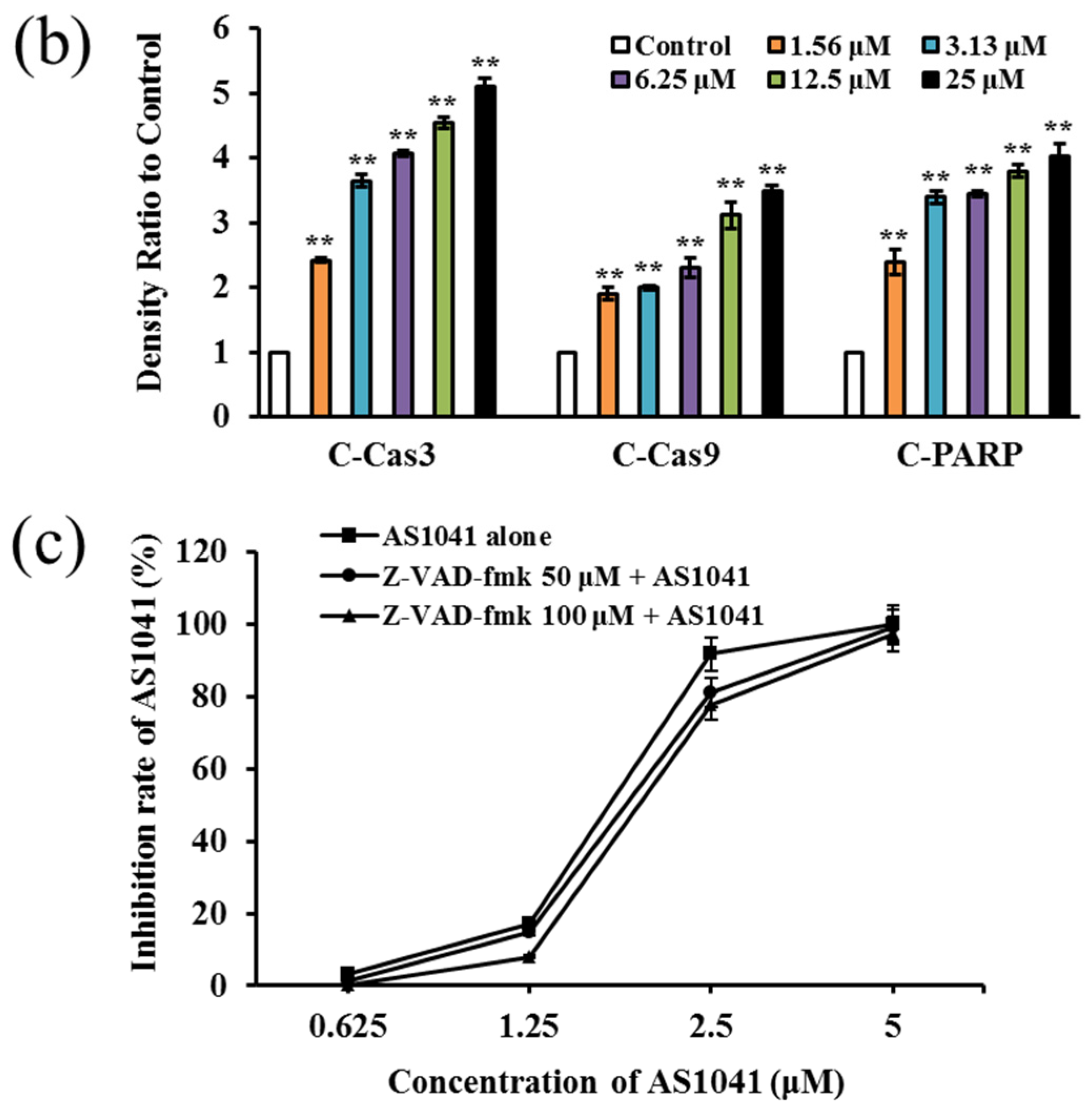
2.4. AS1041 Induces Apoptosis in K562 Cells
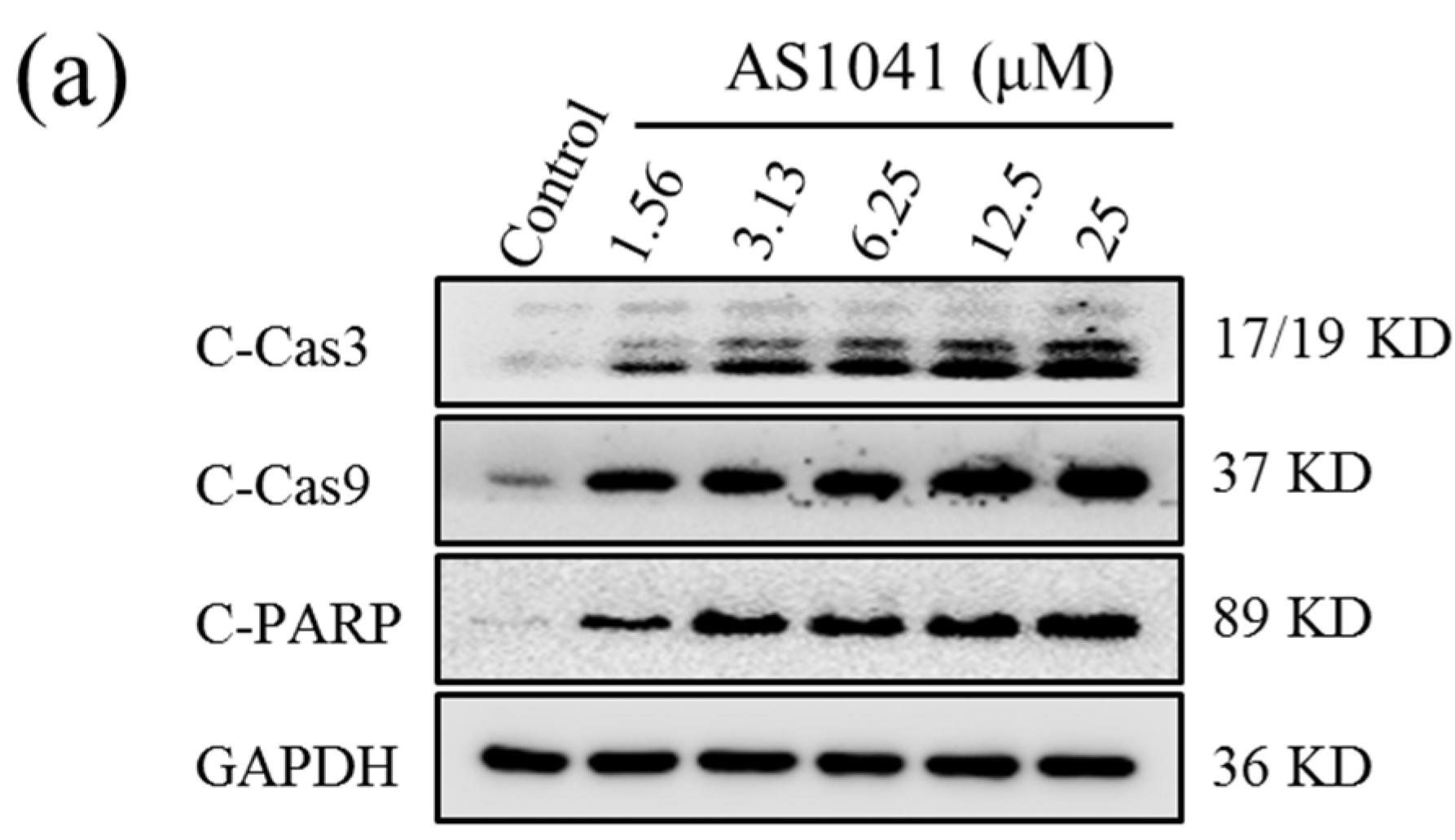
2.5. AS1041 Triggers Non-Caspase-Dependent Apoptosis in K562 Cells
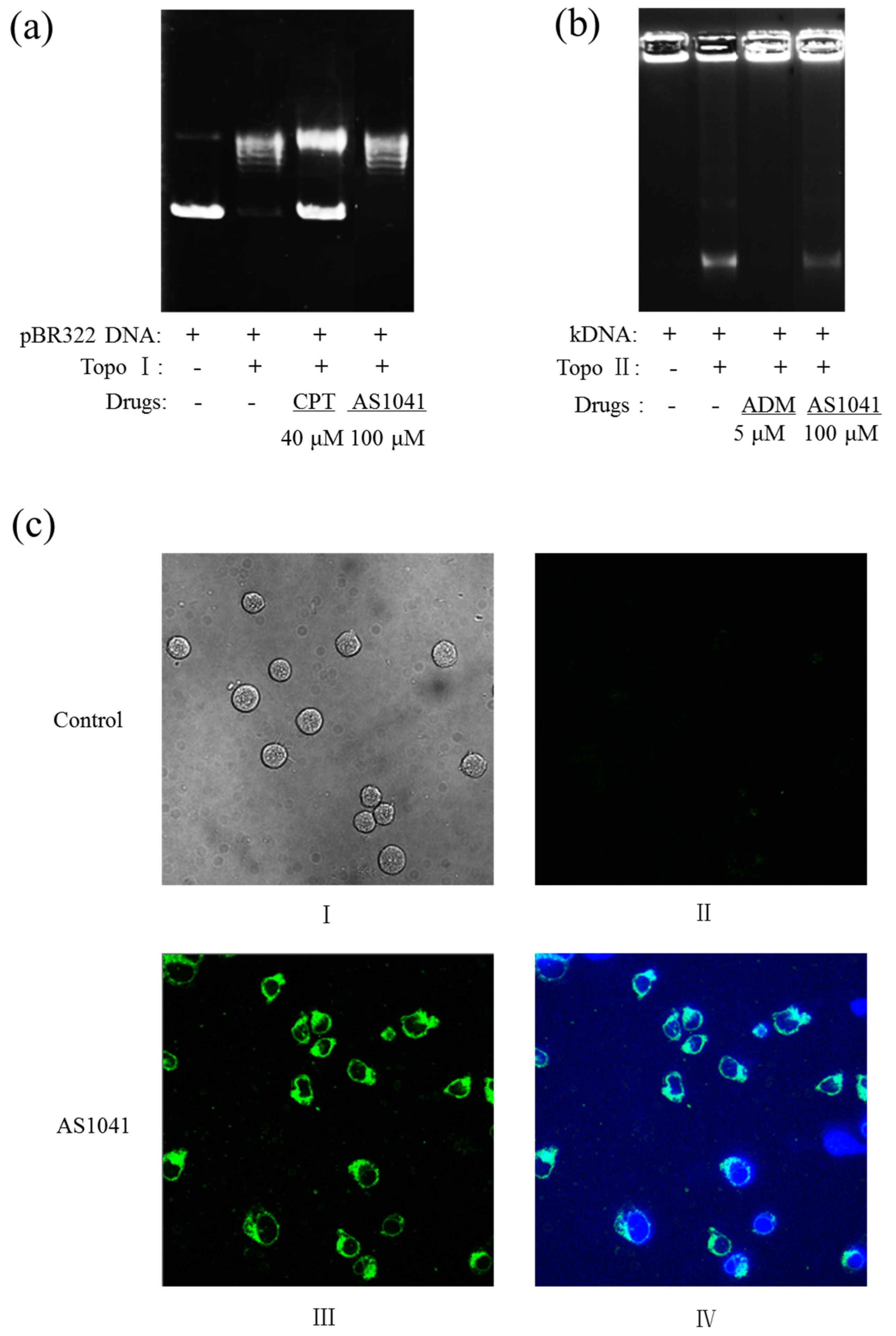
2.6. AS1041 Localizes in the Cytoplasm and Does Not Inhibit the Topoisomerases
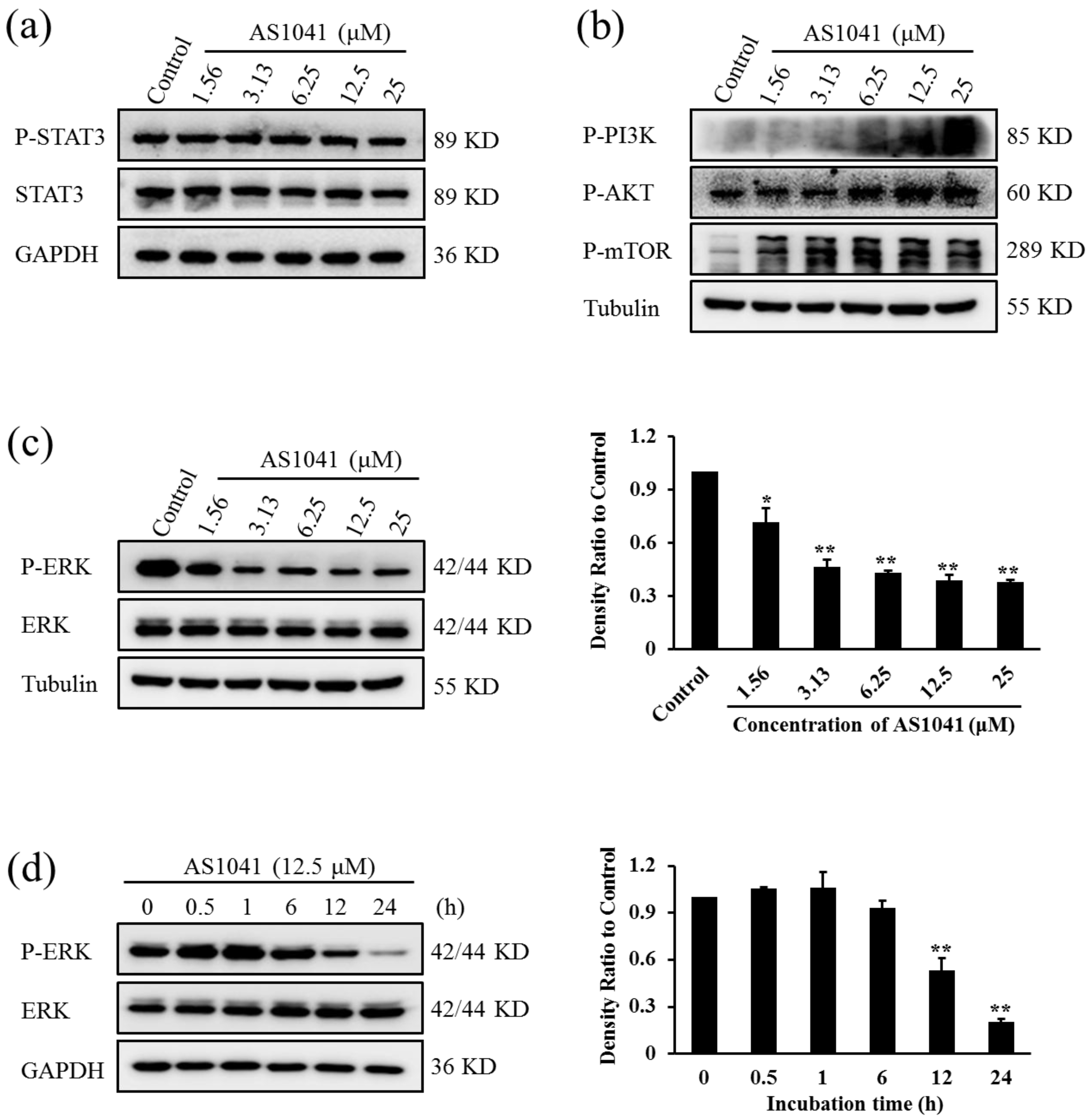
2.7. AS1041 Affects the Survial Signaling
3. Conclusions
4. Materials and Methods
4.1. Reagents and Drug
4.2. Cell Lines and Cell Culture
4.3. Cell Viability Assessment
4.4. Cell Cycle Analysis
4.5. Annexin V-FITC/PI Analysis Assay
4.6. Hoechst 33342 Staining Assay
4.7. Western Blotting Assay
4.8. Soft Agar Colony Formation Assay
4.9. NBT Reduction Assay
4.10. Topo I-Mediated Relaxation of pBR322 DNA Assay
4.11. Topo II-Mediated kDNA Decatenation Assay
4.12. Intracellular Localization of AS1041 in K562 Cells
4.13. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Srinivas, R.; Satterlee, A.; Wang, Y.; Zhang, Y.; Wang, Y.; Huang, L. Theranostic etoposide phosphate/indium nanoparticles for cancer therapy and imaging. Nanoscale 2015, 7, 18542–18551. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.; Cianfrocca, M. Overview of neuropathy associated with taxanes for the treatment of metastatic breast cancer. Cancer Chemother. Pharmacol. 2015, 75, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Kurtulus, G. Strategies targeting DNA Topoisomerase I in cancer chemotherapy: Camptothecins, nanocarriers for camptothecins, crganic non-camptothecin compounds and metal complexes. Curr. Drug Targets 2016, 17, 1928–1939. [Google Scholar]
- Jin, L.; Quan, C.; Hou, X.; Fan, S. Potential pharmacological resources: Natural bioactive compounds from marine-derived fungi. Mar. Drugs 2016, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Liu, M.; Hu, C.; Liao, D.J. Targeting cellular proapoptotic molecules for developing anticancer agents from marine sources. Curr. Drug Targets 2010, 11, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine-sourced anti-cancer and cancer pain control agents in clinical and late preclinical development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Qiao, L.; Liu, Y.; Li, J.; Gu, Q.; Zhu, T.; Che, Q. Preparation of 9-Anthrone Lactone Skeleton Compounds and Their Analogues in Anti-Tumor Applications. China Patent 201710466457.5, 18 August 2017. [Google Scholar]
- Du, L.; Zhu, T.; Fang, Y.; Liu, H.; Gu, Q.; Zhu, W. Aspergiolide A, a novel anthraquinone derivative with naphtho[1,2,3-de]chromene-2,7-dione skeleton isolated from a marine-derived fungus Aspergillus glaucus. Tetrahedron 2007, 63, 1085–1088. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, X.; Li, D.; Zhu, T.; Mo, X.; Li, J. Anticancer efficacy and absorption, distribution, metabolism, and toxicity studies of Aspergiolide A in early drug development. Drug Des. Dev. Ther. 2014, 8, 1965–1977. [Google Scholar]
- Ichimura, A.; Ruike, Y.; Terasawa, K.; Shimizu, K.; Tsujimoto, G. MicroRNA-34a inhibits cell proliferation by repressing mitogen-activated protein kinase kinase 1 during megakaryocytic differentiation of K562 Cells. Mol. Pharmacol. 2010, 77, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Nabissi, M.; Morelli, M.B.; Amantini, C.; Liberati, S.; Santoni, M.; Ricci-Vitiani, L.; Pallini, R.; Santoni, G. Cannabidiol stimulates AML-1a-dependent glial differentiation and inhibits glioma stem-like cells proliferation by inducing autophagy in a TRPV2-dependent manner. Int. J. Cancer 2015, 137, 1855–1869. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, R.; Song, S.; Hua, Z.; Wang, J.; Wang, Y. Angelica sinensis polysaccharide induces erythroid differentiation of human chronic myelogenous leukemia K562 cells. Asian Pac. J. Cancer Prev. 2015, 16, 3715–3721. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Yamaguchi, N. Chapter seven-role of Cyclin B1 levels in DNA damage and DNA damage-induced senescence. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Amsterdam, The Netherlands, 2013; Volume 305, pp. 303–337. [Google Scholar]
- Gavet, O.; Pines, J. Progressive activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev. Cell 2010, 18, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Khammanivong, A.; Wang, C.; Sorenson, B.S.; Ross, K.F.; Herzberg, M.C. S100A8/A9 (Calprotectin) negatively regulates G2/M cell cycle progression and growth of squamous cell carcinoma. PLoS ONE 2013, 8, e69395. [Google Scholar] [CrossRef] [PubMed]
- Pietenpol, J.A.; Stewart, Z.A. Cell cycle checkpoint signaling: Cell cycle arrest versus apoptosis. Toxicology 2002, 181, 475–481. [Google Scholar] [CrossRef]
- Higginbottom, K.; Cummings, M.; Newland, A.C.; Allen, P.D. Etoposide-mediated deregulation of the G2M checkpoint in myeloid leukaemic cell lines results in loss of cell survival. Br. J. Haematol. 2002, 119, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Cytometry of apoptosis. Historical perspective and new advances. Exp. Oncol. 2012, 34, 255–262. [Google Scholar] [PubMed]
- Chen, S.; Cheng, A.; Wang, M.; Peng, X. Detection of apoptosis induced by new type gosling viral enteritis virus in vitro through fluorescein annexin V-FITC/PI double labeling. World J. Gastroenterol. 2008, 14, 2174–2178. [Google Scholar] [CrossRef] [PubMed]
- Blankenberg, F.G. In vivo detection of apoptosis. J. Nucl. Med. NLM 2008, 49 (Suppl. 2), 81S–95S. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Browne, G.; Melino, G.; Cohen, G.M. Ordering of caspases in cells undergoing apoptosis by the intrinsic pathway. Cell Death Differ. 2009, 16, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulos, P.T.; Sofotasiou, M.; Papadopoulou, V.; Polonyfi, K.; Iliakis, T.; Viniou, N. PARP1-driven apoptosis in chronic lymphocytic leukemia. BioMed Res. Int. 2014, 2014, 106713. [Google Scholar] [CrossRef] [PubMed]
- Bröker, L.E.; Kruyt, F.A.E.; Giaccone, G. Cell death independent of caspases: A review. Clin. Cancer Res. 2005, 11, 3155–3162. [Google Scholar] [CrossRef] [PubMed]
- Hegde, R.; Srinivasula, S.; Zhang, Z.; Wassell, R.; Mukattash, R.; Cilenti, L.; DuBois, G.; Lazebnik, Y.; Zervos, A.; Fernandes-Alnemri, T.; et al. Identification of Omi/HtrA2 as a mitochondrial apoptotic serine protease that disrupts inhibitor of apoptosis protein-caspase interaction. J. Biol. Chem. 2002, 277, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Z.; Chen, T. Curcumol induces apoptosis via caspases-independent mitochondrial pathway in human lung adenocarcinoma ASTC-a-1 cells. Med. Oncol. 2011, 28, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Switalska, M.; Grynkiewicz, G.; Strzadala, L.; Wietrzyk, J. Novel genistein derivatives induce cell death and cell cycle arrest through different mechanisms. Nutr. Cancer 2013, 65, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, G.; De Benedictis, I.; Malfitano, A.; Morelli, G.; Novellino, E.; Marasco, D. Interactions of cisplatin analogues with lysozyme: A comparative analysis. BioMetals 2017, 30, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Sagineedu, S.R.; Sumon, S.H.; Sidik, S.M.; Phillips, R.; Lajis, N.H.; Stanslas, J. NCI in vitro and in silico anticancer screen, cell cycle pertubation and apoptosis-inducing potential of new acylated, benzylidene and isopropylidene derivatives of andrographolide. Environ. Toxicol. Pharmacol. 2014, 38, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Wake, M.S.; Watson, C.J. STAT3 the oncogene-still eluding therapy? FEBS J. 2015, 282, 2600–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zhang, Y.; Li, R.; Ye, J.; Li, H.; Zhang, Y.; Ma, Z.; Li, J.; Zhong, X.; Yang, X. Tetrandrine suppresses human glioma growth by inhibiting cell survival, proliferation and tumour angiogenesis through attenuating STAT3 phosphorylation. Eur. J. Pharmacol. 2015, 764, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Xu, X.; Yang, B.; Chen, Z.; Cheng, R. Saponins from rubus parvifolius L. induce apoptosis in human chronic myeloid leukemia cells through AMPK activation and STAT3 inhibition. Asian Pac. J. Cancer Prev. 2014, 15, 5455–5461. [Google Scholar] [CrossRef] [PubMed]
- Bertacchini, J.; Heidari, N.; Mediani, L.; Capitani, S.; Shahjahani, M.; Ahmadzadeh, A.; Saki, N. Targeting PI3K/AKT/mTOR network for treatment of leukemia. Cell Mol. Life Sci. 2015, 72, 2337–2347. [Google Scholar] [CrossRef] [PubMed]
- Polivka, J.; Janku, F. Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol. Ther. 2014, 142, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, Y.; Li, H.; Bi, S.; Zhao, J. ERK/CANP rapid signaling mediates 17β-estradiol-induced proliferation of human breast cancer cell line MCF-7 cells. Int. J. Clin. Exp. Med. 2014, 7, 156–162. [Google Scholar] [PubMed]
- Dong, L.; Liu, L.; Ma, C.; Li, J.; Du, C.; Xu, S.; Han, L.; Li, L.; Wang, X. E-cadherin promotes proliferation of human ovarian cancer cells in vitro via activating MEK/ERK pathway. Acta Pharmacol. Sin. 2012, 33, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Lompardía, S.L.; Díaz, M.; Papademetrio, D.L.; Pibuel, M.; Álvarez, É.; Hajos, S.E. 4-methylumbelliferone and imatinib combination enhances senescence induction in chronic myeloid leukemia cell lines. Investig. New Drugs 2017, 35, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ding, Q.; Ding, Y.; Lu, L.; Wang, X.; Zhang, Y.; Zhang, X.; Guo, Q.; Zhao, L. Oroxylin a reverses the drug resistance of chronic myelogenous leukemia cells to imatinib through CXCL12/CXCR7 axis in bone marrow microenvironment. Mol. Carcinog. 2017, 56, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Hu, S.; Liu, T.; Wang, F.; Wang, X.; Wu, G.; Zhang, Y.; Sui, M.; Liu, H.; Jiang, L. Nitidine chloride inhibits proliferation and induces apoptosis in colorectal cancer cells by suppressing the ERK signaling pathway. Mol. Med. Rep. 2016, 13, 2536–2542. [Google Scholar] [CrossRef] [PubMed]
- Ponnurangam, S.; Standing, D.; Rangarajan, P.; Subramaniam, D. Tandutinib inhibits the Akt/mTOR signaling pathway to inhibit colon cancer growth. Mol. Cancer Ther. 2013, 12, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Okada, H.; Ruland, J.; Liu, L.; Stambolic, V.; Mak, T.W.; Ingram, A.J. Akt is activated in response to an apoptotic signal. J. Biol. Chem. 2001, 276, 30461–30466. [Google Scholar] [CrossRef] [PubMed]
- Follo, M.Y.; Manzoli, L.; Poli, A.; McCubrey, J.A.; Cocco, L. PLC and PI3K/Akt/mTOR signalling in disease and cancer. Adv. Biol. Regul. 2015, 57, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Hsia, T.; Yeh, M.; Chen, T.; Chen, Y.; Chen, J.; Wei, Y.; Tu, C.; Huang, W. MEK inhibitors induce Akt activation and drug resistance by suppressing negative feedback ERK-mediated HER2 phosphorylation at Thr701. Mol. Oncol. 2017, 11, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Turke, A.B.; Song, Y.; Costa, C.; Cook, R.; Arteaga, C.L.; Asara, J.M.; Engelman, J.A. MEK inhibition leads to PI3K/AKT activation by relieving a negative feedback on ERBB receptors. Cancer Res. 2012, 72, 3228–3237. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Kim, H.; Han, S.; Hur, H.; Oh, D.; Im, S.; Bang, Y.; Kim, T. Combination of EGFR and MEK1/2 inhibitor shows synergistic effects by suppressing EGFR/HER3-dependent AKT activation in human gastric cancer cells. Mol. Cancer Ther. 2009, 8, 2526–2536. [Google Scholar] [CrossRef] [PubMed]
- Mirzoeva, O.K.; Das, D.; Heiser, L.M.; Bhattacharya, S.; Siwak, D.; Gendelman, R.; Bayani, N.; Wang, N.J.; Neve, R.M.; Guan, Y.; et al. Basal subtype and MAPK/ERK kinase (MEK)-phosphoinositide 3-Kinase feedback signaling determine susceptibility of breast cancer cells to MEK inhibition. Cancer Res. 2009, 69, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Hoeflich, K.P.; O’Brien, C.; Boyd, Z.; Cavet, G.; Guerrero, S.; Jung, K.; Januario, T.; Savage, H.; Punnoose, E.; Truong, T. In vivo antitumor activity of MEK and phosphatidylinositol 3-kinase inhibitors in basal-like breast cancer models. Clin. Cancer Res. 2009, 15, 4649–4664. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AS1041 (μM) | Imatinib (μM) | CI |
|---|---|---|
| 6.25 | 1.00 | 0.59 |
| 6.25 | 0.50 | 0.32 |
| 6.25 | 0.25 | 0.18 |
| 6.25 | 0.13 | 0.12 |
| 6.25 | 0.06 | 0.08 |
| 3.13 | 1.00 | 0.57 |
| 3.13 | 0.50 | 0.30 |
| 3.13 | 0.25 | 0.16 |
| 3.13 | 0.13 | 0.09 |
| 3.13 | 0.06 | 0.06 |
| 1.56 | 1.00 | 0.56 |
| 1.56 | 0.50 | 0.28 |
| 1.56 | 0.25 | 0.15 |
| 1.56 | 0.13 | 0.59 |
| 1.56 | 0.06 | 0.72 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, F.; Qiao, L.; Chen, Y.; Qi, X.; Liu, Y.; Li, D.; Gu, Q.; Li, J.; Liu, M. AS1041, a Novel Synthesized Derivative of Marine Natural Compound Aspergiolide A, Arrests Cell Cycle, Induces Apoptosis, and Inhibits ERK Activation in K562 Cells. Mar. Drugs 2017, 15, 346. https://doi.org/10.3390/md15110346
Yuan F, Qiao L, Chen Y, Qi X, Liu Y, Li D, Gu Q, Li J, Liu M. AS1041, a Novel Synthesized Derivative of Marine Natural Compound Aspergiolide A, Arrests Cell Cycle, Induces Apoptosis, and Inhibits ERK Activation in K562 Cells. Marine Drugs. 2017; 15(11):346. https://doi.org/10.3390/md15110346
Chicago/Turabian StyleYuan, Fengli, Liang Qiao, Yinghan Chen, Xin Qi, Yankai Liu, Dehai Li, Qianqun Gu, Jing Li, and Ming Liu. 2017. "AS1041, a Novel Synthesized Derivative of Marine Natural Compound Aspergiolide A, Arrests Cell Cycle, Induces Apoptosis, and Inhibits ERK Activation in K562 Cells" Marine Drugs 15, no. 11: 346. https://doi.org/10.3390/md15110346
APA StyleYuan, F., Qiao, L., Chen, Y., Qi, X., Liu, Y., Li, D., Gu, Q., Li, J., & Liu, M. (2017). AS1041, a Novel Synthesized Derivative of Marine Natural Compound Aspergiolide A, Arrests Cell Cycle, Induces Apoptosis, and Inhibits ERK Activation in K562 Cells. Marine Drugs, 15(11), 346. https://doi.org/10.3390/md15110346