
Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of EXCB on the Viability of LA Myocytes
2.2. Effects of EXCB on the Action Potential Morphology of LA Myocytes
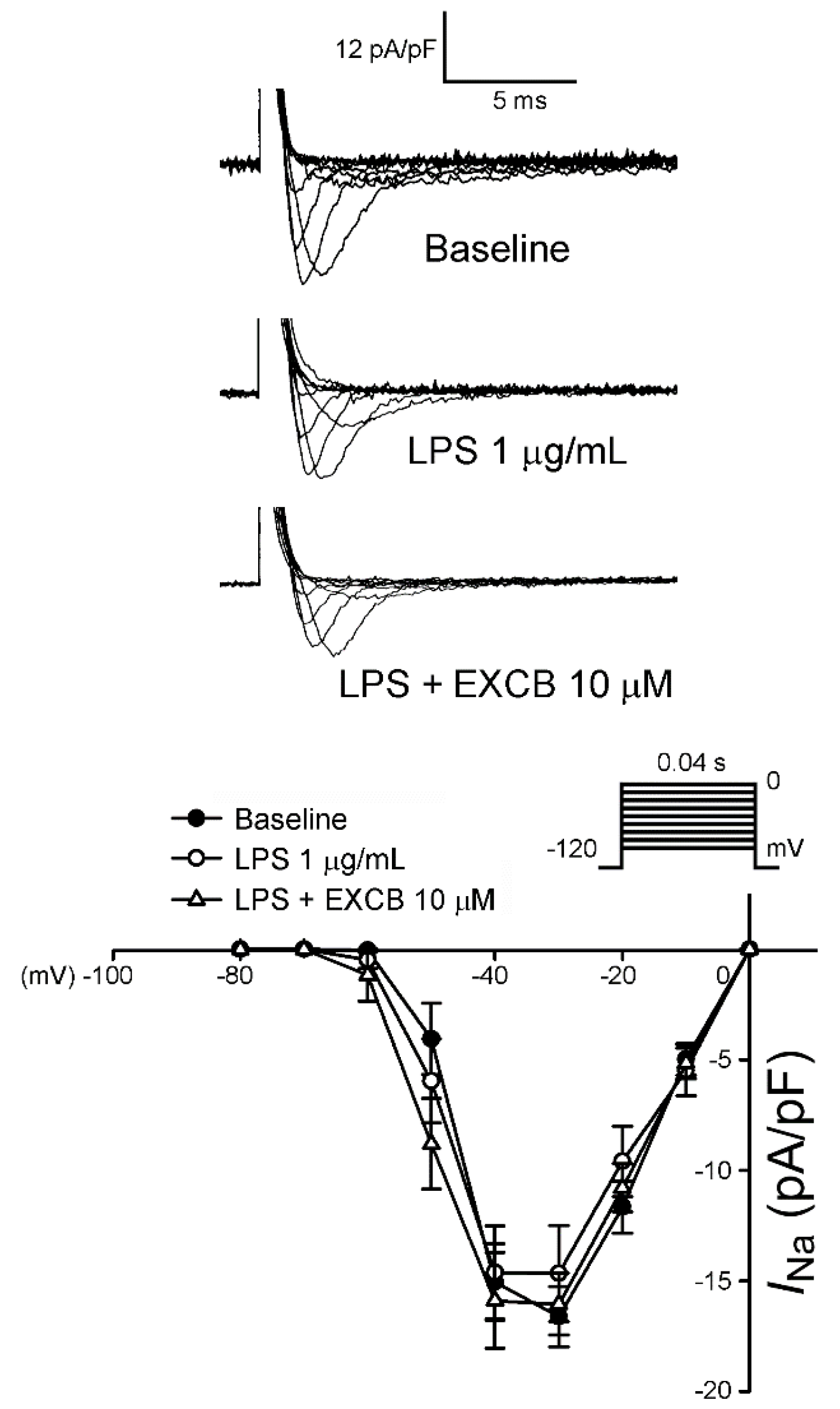
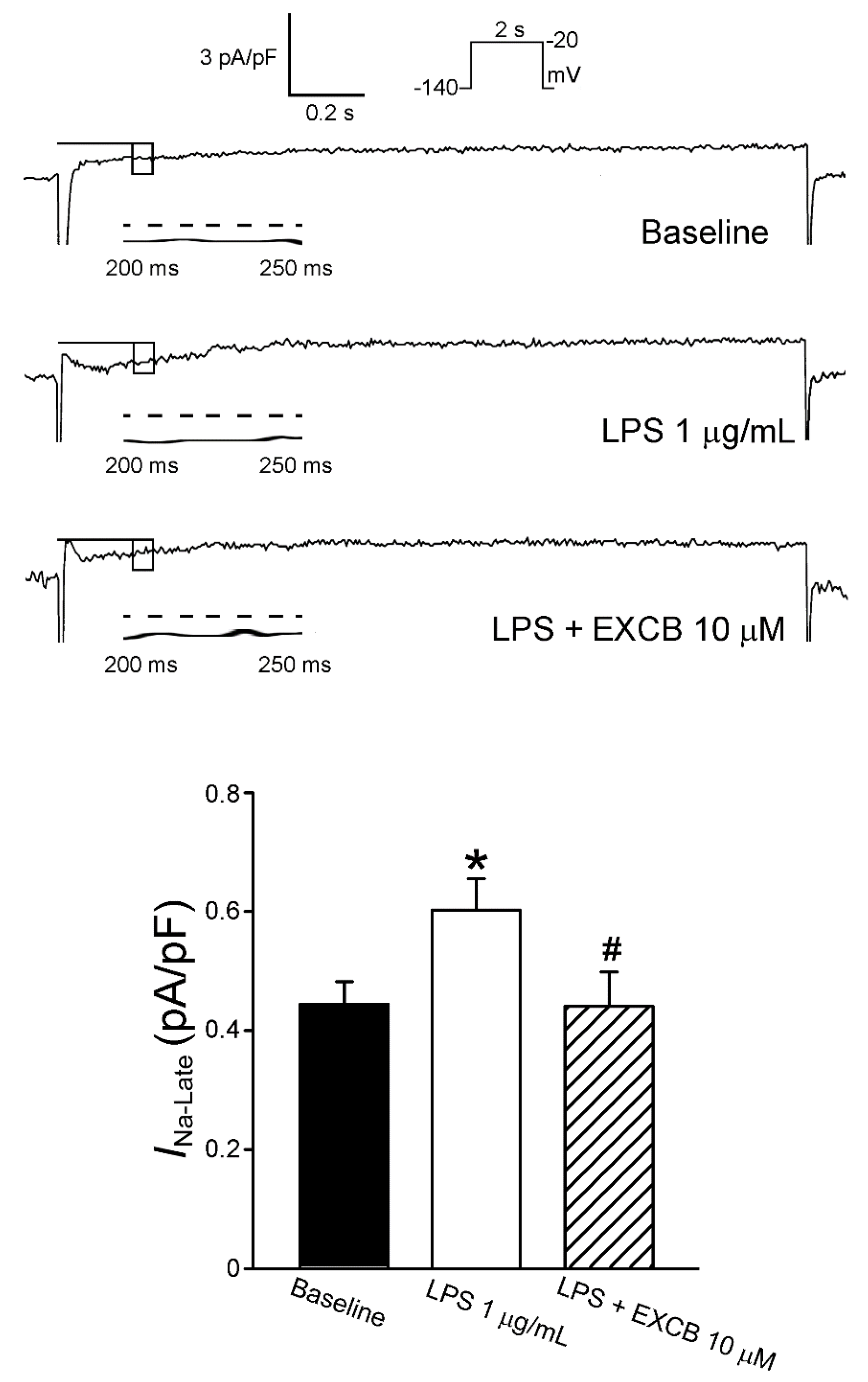
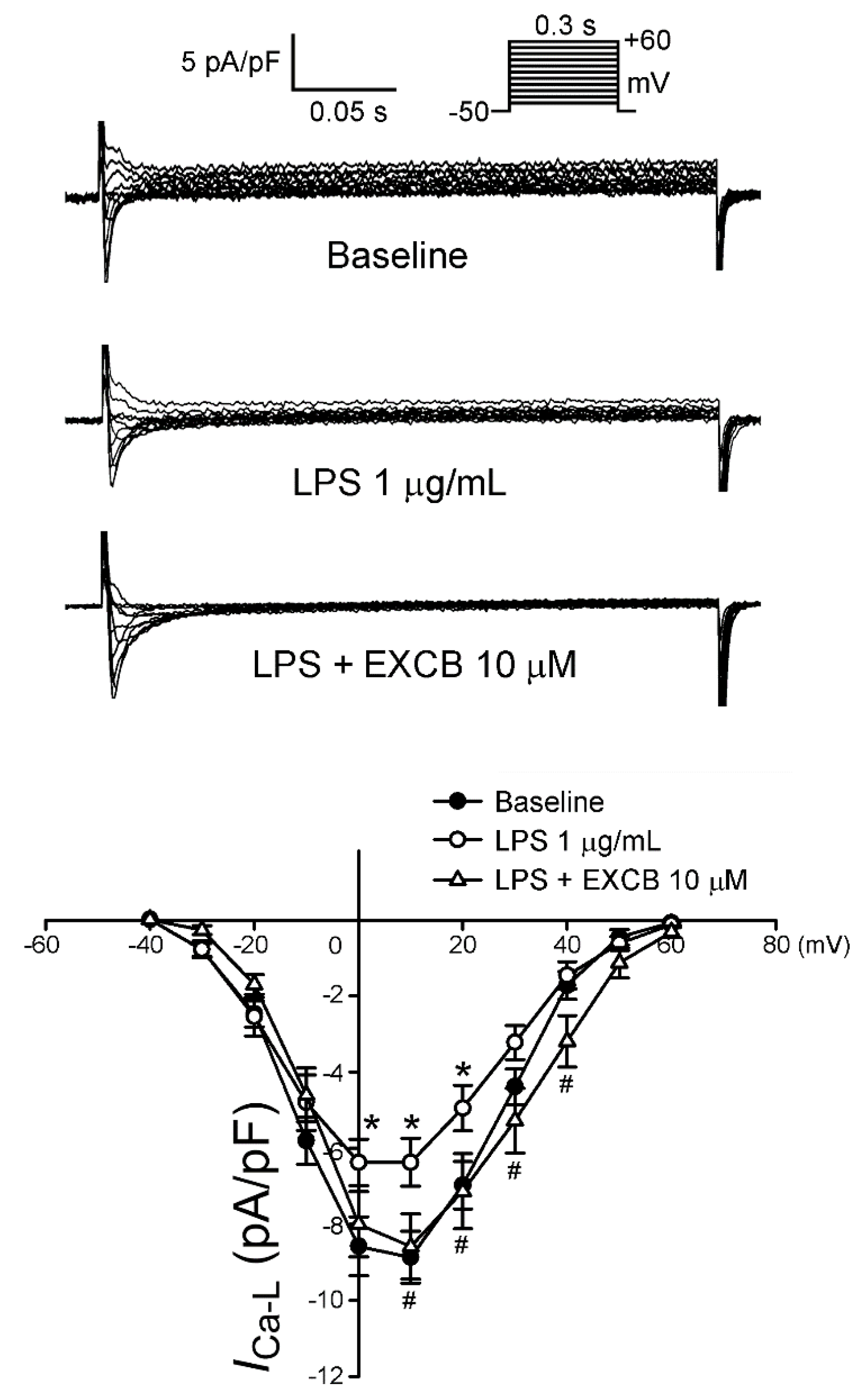
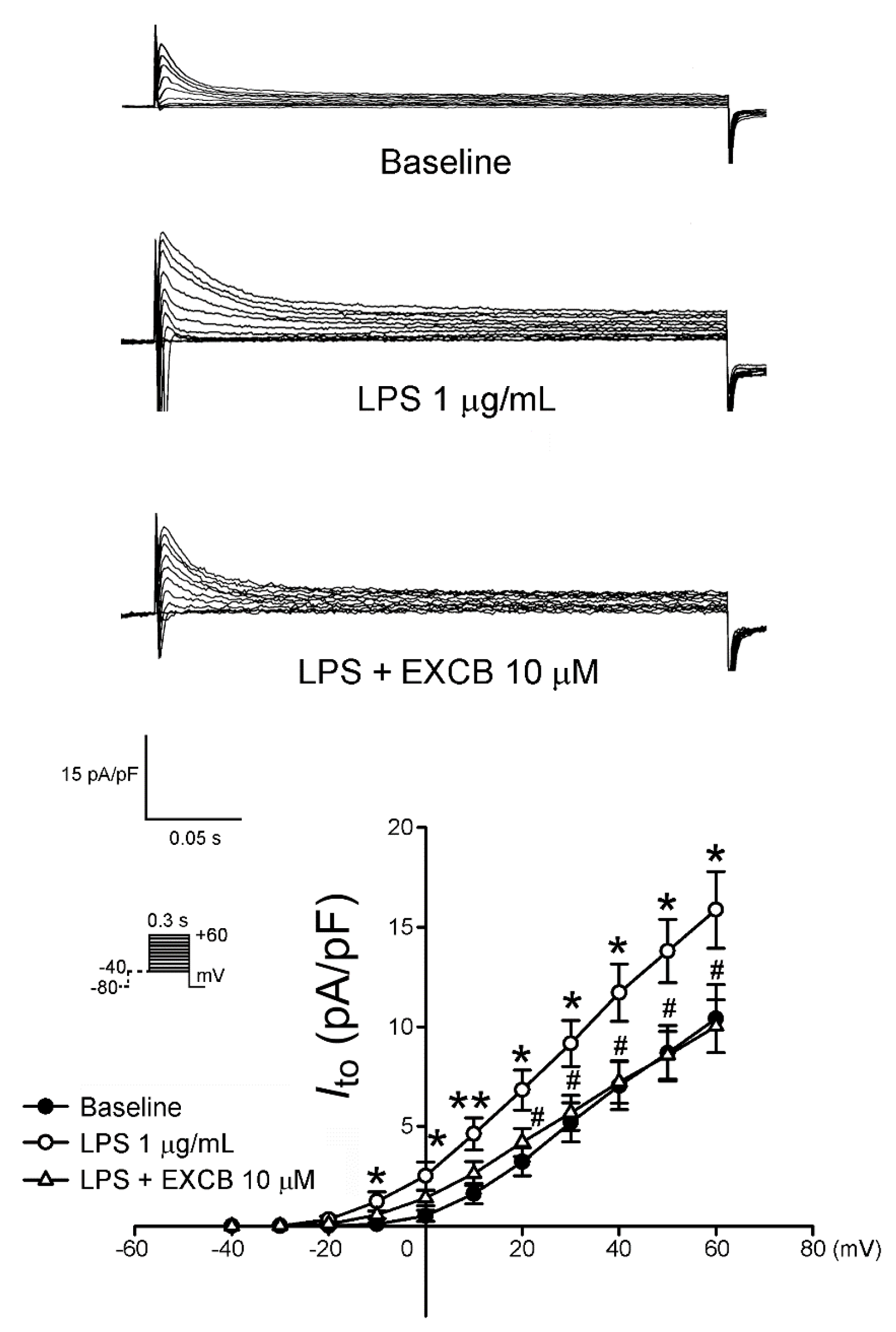
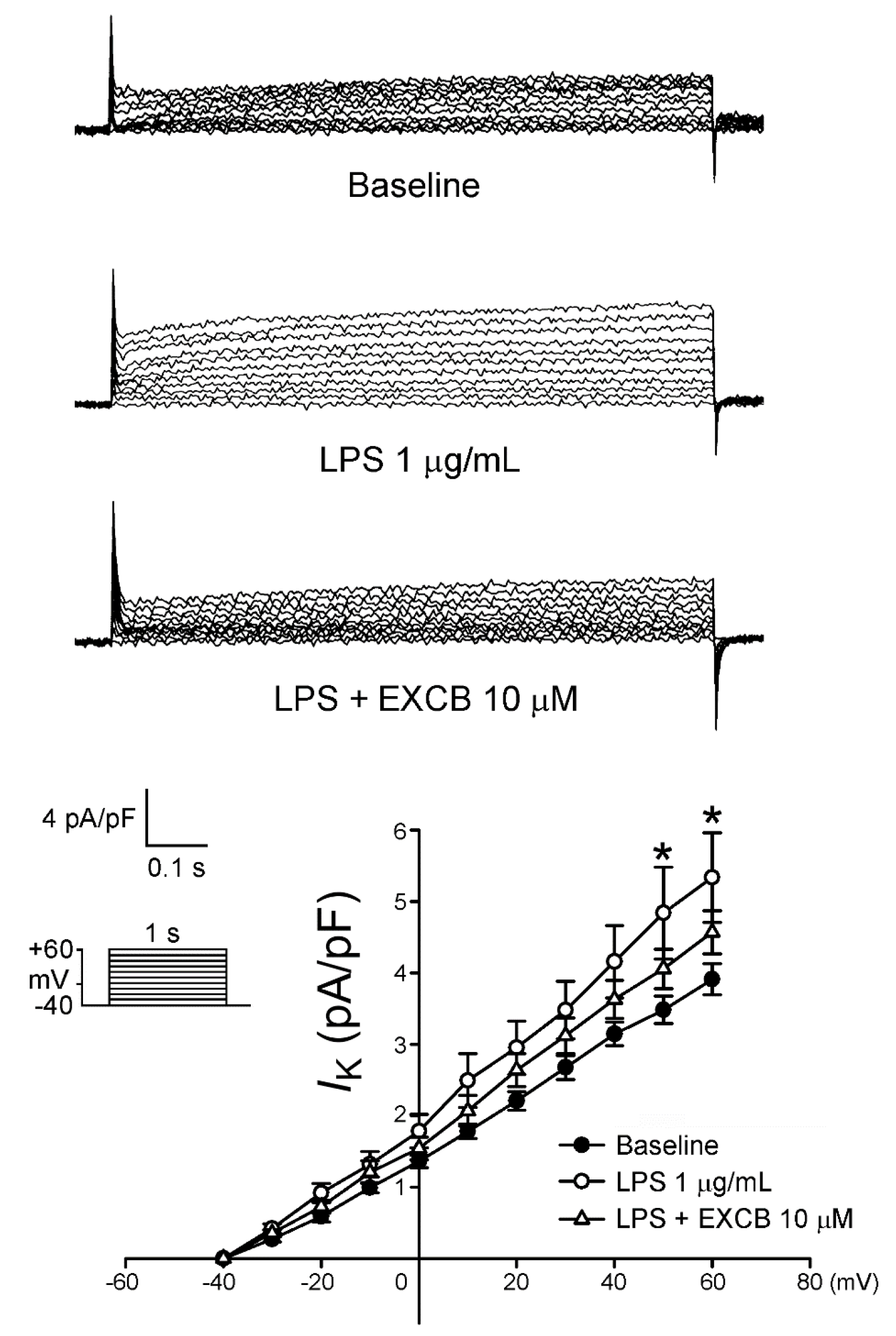
2.3. Effects of EXCB on the Membrane Currents of LA Myocytes
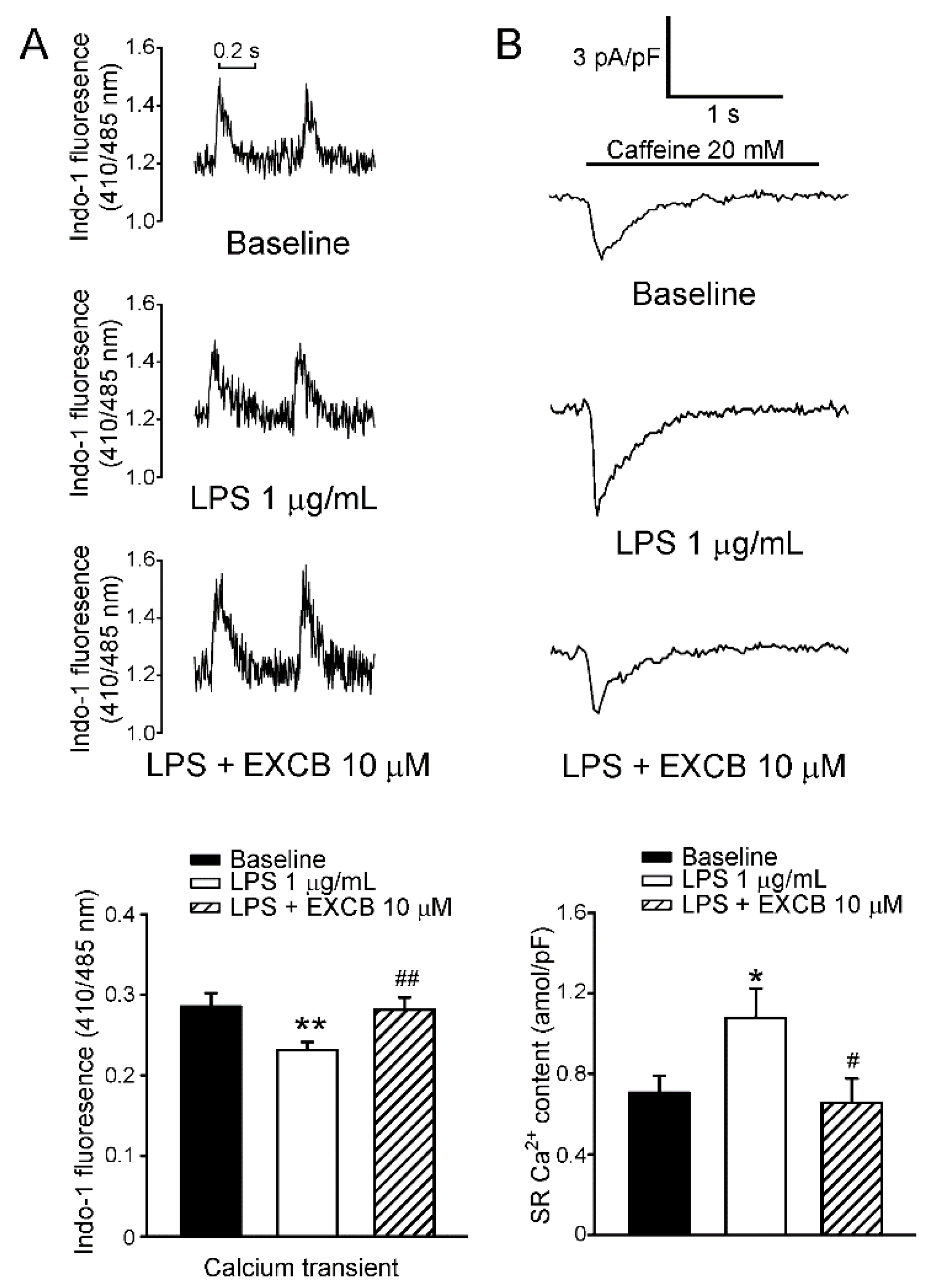
2.4. Effects of EXCB on Calcium Handling of LA Myocytes
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Isolation of Single LA Myocytes
4.3. Electrophysiological Study
4.4. Measurement of the Changes in the Intracellular Calcium Concentration
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Francisco, R.B.; Manuel, R.B.; Julián, G.C.; Maria, J.H.R. Sepsis-induced Cardiomyopathy. Curr. Cardiol. Rev. 2011, 7, 163–183. [Google Scholar]
- Van Amersfoort, E.S.; Van Berkel, T.J.; Kuiper, J. Receptors, mediators, and mechanisms involved in bacterial sepsis and septic shock. Clin. Microbiol. Rev. 2003, 16, 379–414. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M. Endotoxins and other sepsis triggers. Contrib. Nephrol. 2010, 167, 14–24. [Google Scholar] [PubMed]
- Jiang, S.; Zhu, W.; Li, C.; Zhang, X.; Lu, T.; Ding, Z.; Cao, K.; Liu, L. α-Lipoic acid attenuates LPS-induced cardiac dysfunction through a PI3K/Akt-dependent mechanism. Int. Immunopharmacol. 2013, 16, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Han, A.R.; Park, E.Y.; Kim, J.Y.; Cho, W.; Lee, J.; Seo, E.K.; Lee, K.T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-kappa B inactivation in RAW 264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.R.; Treuer, A.; Sun, Q.A.; Stamler, J.S.; Hare, J.M. S-Nitrosylation of cardiac ion channels. J. Cardiovasc. Pharmacol. 2009, 54, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Lin, S.C.; Feng, C.W.; Chen, P.C.; Su, Y.D.; Li, C.M.; Yang, S.N.; Jean, Y.H.; Sung, P.J.; Duh, C.Y.; et al. Anti-Inflammatory and Analgesic Effects of the Marine-Derived Compound Excavatolide B Isolated from the Culture-Type Formosan Gorgonian Briareum excavatum. Mar. Drugs 2015, 13, 2559–2579. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, B.K.; Yang, H.H.; Sung, P.J.; Weng, C.F. Excavatolide B inhibits nonsmall cell lung cancer proliferation by altering peroxisome proliferator activated receptor gamma expression and PTEN/AKT/NF-Kβ expression. Environ. Toxicol. 2017, 32, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Joyner, P.M.; Waters, A.L.; Williams, R.B.; Powell, D.R.; Janakiram, N.B.; Rao, C.V.; Cichewicz, R.H. Briarane Diterpenes Diminish COX-2 Expression in Human Colon Adenocarcinoma Cells. J. Nat. Prod. 2011, 74, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, R.B.; Larson, M.G.; Yamamoto, J.F.; Kathiresan, S.; Rong, J.; Levy, D.; Keaney, J.F., Jr.; Wang, T.J.; Vasan, R.S.; Benjamin, E.J. Relation of multiple inflammatory biomarkers to incident atrial fibrillation. Am. J. Cardiol. 2009, 104, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, R.; Iwasaki, Y.K.; Miyauchi, Y.; Hirayama, Y.; Kobayashi, Y.; Katoh, T.; Mizuno, K.; Sekiguchi, A.; Yamashita, T. lipopolysaccharide induces atrial arrhythmogenesis via down-regulation of l-type Ca2+ channel genes in rats. Int. Heart J. 2009, 50, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Dernellis, J.; Panaretou, M. Relationship between C-reactive protein concentrations during glucocorticoid therapy and recurrent atrial fibrillation. Eur. Heart J. 2004, 25, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Siu, C.W.; Lau, C.P.; Tse, H.F. Prevention of atrial fibrillation recurrence by statin therapy in patients with lone atrial fibrillation after successful cardioversion. Am. J. Cardiol. 2003, 92, 1343–1345. [Google Scholar] [CrossRef] [PubMed]
- Boos, C.J.; Lip, G.Y. The Role of Inflammation in Atrial Fibrillation. Int. J. Clin. Pract. 2005, 59, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Seguin, P.; Signouret, T.; Laviolle, B.; Branger, B.; Mallédant, Y. Incidence and risk factors of atrial fibrillation in a surgical intensive care unit. Crit. Care Med. 2004, 32, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Seguin, P.; Laviolle, B.; Maurice, A.; Leclercq, C.; Mallédant, Y. Atrial fibrillation in trauma patients requiring intensive care. Intensive Care Med. 2006, 32, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Court, O.; Kumar, A.; Parrillo, J.E.; Kumar, A. Clinical review: Myocardial depression in sepsis and septic shock. Crit. Care 2002, 6, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Hatakeyama, N.; Yamamoto, S.; Kinoshita, H.; Matsuda, N.; Hattori, Y.; Yamazaki, M. Role of ion channels in sepsis-induced atrial tachyarrhythmias in guinea pigs. Br. J. Pharmacol. 2012, 166, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, D.R.; Pond, A.L.; Lamorgese, M.; Rossie, S.S.; McCarthy, P.M.; Nerbonne, J.M. Atrial l-type Ca2+ currents and human atrial fibrillation. Circ. Res. 1999, 85, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Melnyk, P.; Feng, J.; Wang, Z.; Petrecca, K.; Shrier, A.; Nattel, S. Effects of experimental heart failure on atrial cellular and ionic electrophysiology. Circulation 2000, 101, 2631–2638. [Google Scholar] [CrossRef] [PubMed]
- Le Grand, B.L.; Hatem, S.; Deroubaix, E.; Couétil, J.P.; Coraboeuf, E. Depressed transient outward and calcium currents in dilated human atria. Cardiovasc. Res. 1994, 28, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Makielski, J.C.; Farley, A.L. Na(+) current in human ventricle: Implications for sodium loading and homeostasis. J. Cardiovasc. Electrophysiol. 2006, 17, S15–S20. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.K.; Chen, Y.C.; Chen, J.H.; Chen, S.A.; Chen, Y.J. Adipocytes modulate the electrophysiology of atrial myocytes: Implications in obesity-induced atrial fibrillation. Basic Res. Cardiol. 2012, 107, 293. [Google Scholar] [CrossRef] [PubMed]
- Burashnikov, A.; Di Diego, J.M.; Zygmunt, A.C.; Belardinelli, L.; Antzelevitch, C. Atrium-selective sodium channel block as a strategy for suppression of atrial fibrillation: Differences in sodium channel inactivation between atria and ventricles and the role of ranolazine. Circulation 2007, 116, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Shryock, J.C.; Belardinelli, L. A slowly inactivating sodium current contributes to spontaneous diastolic depolarization of atrial myocytes. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1254–H1262. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Y.; Cheng, C.C.; Chen, Y.C.; Chen, S.A.; Chen, Y.J. ATX-II-induced pulmonary vein arrhythmogenesis related to atrial fibrillation and long QT syndrome. Eur. J. Clin. Investig. 2012, 42, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.S. A novel mechanism for the treatment of angina, arrhythmias, and diastolic dysfunction: Inhibition of late I(Na) using ranolazine. J. Cardiovasc. Pharmacol. 2009, 54, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Tani, M. Mechanisms of Ca2+ overload in reperfused ischemic myocardium. Annu. Rev. Physiol. 1990, 52, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Wongcharoen, W.; Chen, Y.C.; Chen, Y.J.; Chang, C.M.; Yeh, H.I.; Lin, C.I.; Chen, S.A. Effects of a Na+/Ca2+ exchanger inhibitor on pulmonary vein electrical activity and ouabain-induced arrhythmogenicity. Cardiovasc. Res. 2006, 70, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Shigekawa, M.; Iwamoto, T. Cardiac Na(+)-Ca(2+) exchange: Molecular and pharmacological aspects. Circ. Res. 2001, 88, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, V.A.; Reznikov, V.; Undrovinas, N.A.; Sabbah, H.N.; Undrovinas, A. Modulation of late sodium current by Ca2+, calmodulin, and CaMKII in normal and failing dog cardiomyocytes: Similarities and differences. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1597–H1608. [Google Scholar] [CrossRef] [PubMed]
- Noble, D.; Noble, P.J. Late sodium current in the pathophysiology of cardiovascular disease: Consequences of sodium-calcium overload. Heart 2006, 92 (Suppl. S4), iv1–iv5. [Google Scholar] [CrossRef] [PubMed]
- Bers, D.M.; Bassani, J.W.; Bassani, R.A. Na-Ca exchange and Ca fluxes during contraction and relaxation in mammalian ventricular muscle. Ann. N. Y. Acad. Sci. 1996, 779, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.W. Digitalis: Mechanisms of action and clinical use. N. Engl. J. Med. 1988, 318, 358–365. [Google Scholar] [PubMed]
- Li, G.R.; Feng, J.; Yue, L.; Carrier, M.; Nattel, S. Evidence for two components of delayed rectifier K+ current in human ventricular myocytes. Circ. Res. 1996, 78, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Li, G.R.; Feng, J.; Wang, Z.; Fermini, B.; Nattel, S. Comparative mechanisms of 4-aminopyridine-resistant Ito in human and rabbit atrial myocytes. Am. J. Physiol. 1995, 269, H463–H472. [Google Scholar] [PubMed]
- Nattel, S. New ideas about atrial fibrillation 50 years on. Nature 2002, 415, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, K.; Brendel, J.; Rosenstein, B.; Bleich, M.; Busch, A.E.; Wirth, K.J. Atrial-selective antiarrhythmic actions of novel Ikur vs. Ikr, Iks, and IKAch class Ic drugs and beta blockers in pigs. Med. Sci. Monit. 2004, 10, BR221–BR228. [Google Scholar] [PubMed]
- Matalon, S.; Hardiman, K.M.; Jain, L.; Eaton, D.C.; Kotlikoff, M.; Eu, J.P.; Sun, J.; Meissner, G.; Stamler, J.S. Regulation of ion channel structure and function by reactive oxygen-nitrogen species. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, L1184–L1189. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. USA 2004, 101, 4003–4008. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.R.; Kang, M.; Shirwany, N.; Malykhina, A.P.; Drozd, M.; Akbarali, H.I. Nitrotyrosylation of Ca2+ channels prevents c-Src kinase regulation of colonic smooth muscle contractility in experimental colitis. J. Pharmacol. Exp. Ther. 2007, 322, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Guide for the Care and Use of Laboratory Animals; United States National Institutes of Health: Bethesda, MD, USA, 1996; Publication No. 85-23.
- Monnerat-Cahli, G.; Alonso, H.; Gallego, M.; Alarcón, M.L.; Bassani, R.A.; Casis, O.; Medei, E. Toll-like receptor 4 activation promotes cardiac arrhythmias by decreasing the transient outward potassium current (Ito) through an IRF3-dependent and MyD88-independent pathway. J. Mol. Cell. Cardiol. 2014, 76, 116–125. [Google Scholar] [CrossRef] [PubMed]









© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, H.-R.; Tai, B.-Y.; Cheng, P.-Y.; Chen, P.-N.; Sung, P.-J.; Wen, Z.-H.; Hsu, C.-H. Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes. Mar. Drugs 2017, 15, 25. https://doi.org/10.3390/md15020025
Hwang H-R, Tai B-Y, Cheng P-Y, Chen P-N, Sung P-J, Wen Z-H, Hsu C-H. Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes. Marine Drugs. 2017; 15(2):25. https://doi.org/10.3390/md15020025
Chicago/Turabian StyleHwang, Hwong-Ru, Buh-Yuan Tai, Pao-Yun Cheng, Ping-Nan Chen, Ping-Jyun Sung, Zhi-Hong Wen, and Chih-Hsueng Hsu. 2017. "Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes" Marine Drugs 15, no. 2: 25. https://doi.org/10.3390/md15020025
APA StyleHwang, H. -R., Tai, B. -Y., Cheng, P. -Y., Chen, P. -N., Sung, P. -J., Wen, Z. -H., & Hsu, C. -H. (2017). Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes. Marine Drugs, 15(2), 25. https://doi.org/10.3390/md15020025