
Running the Stop Sign: Readthrough of a Premature UAG Termination Signal in the Translation of a Zebrafish (Danio rerio) Taurine Biosynthetic Enzyme


Abstract
:1. Introduction
2. Results
2.1. Standard Feeds for Zebrafish Contain Taurine
2.2. Zebrafish Synthesize Sufficient Taurine for Homeostasis
2.3. Wild-type and sa9430 Fish Have Similar Fitness Despite Lower Taurine Levels in sa9430 Embryos
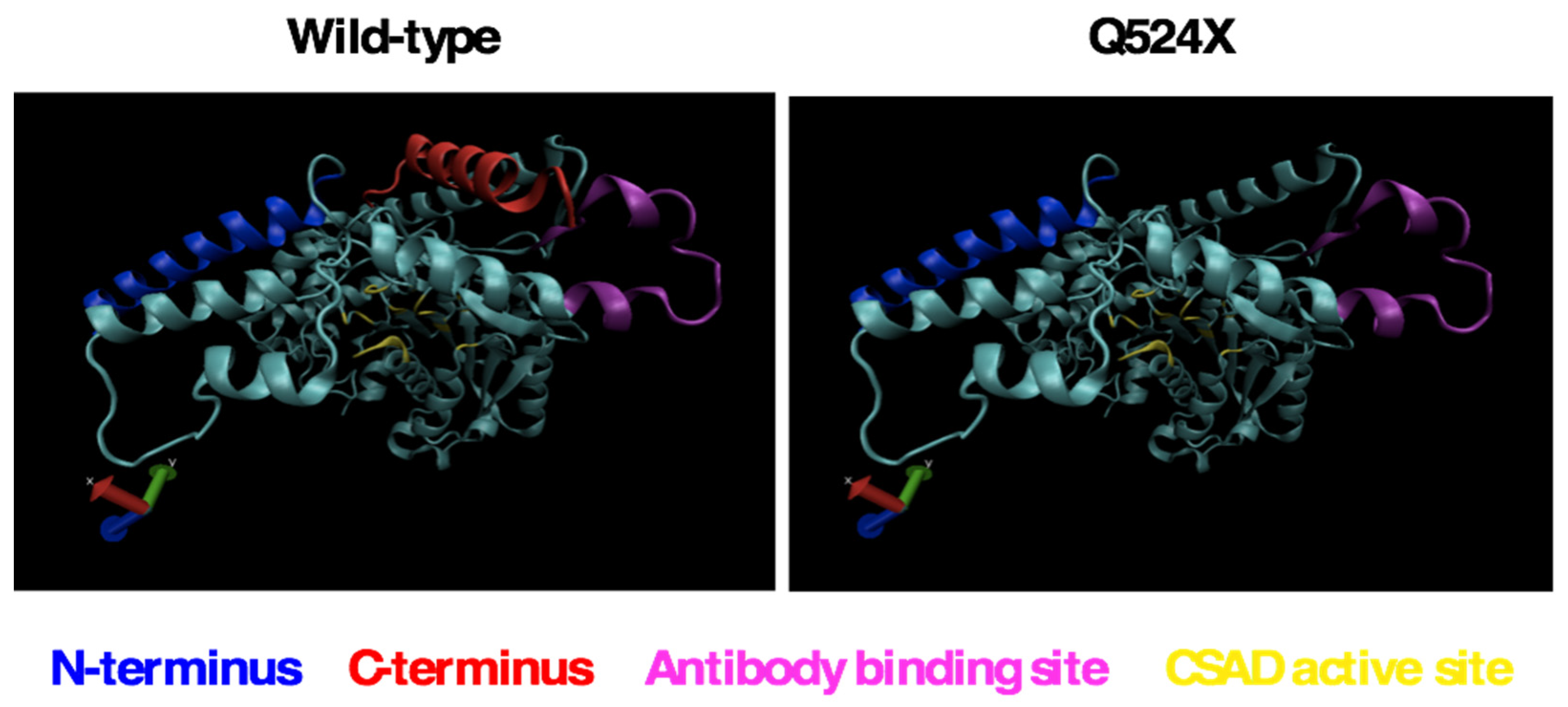
2.4. Wild-type and sa9430 CSAD Proteins Have Identical Modeled Conformations
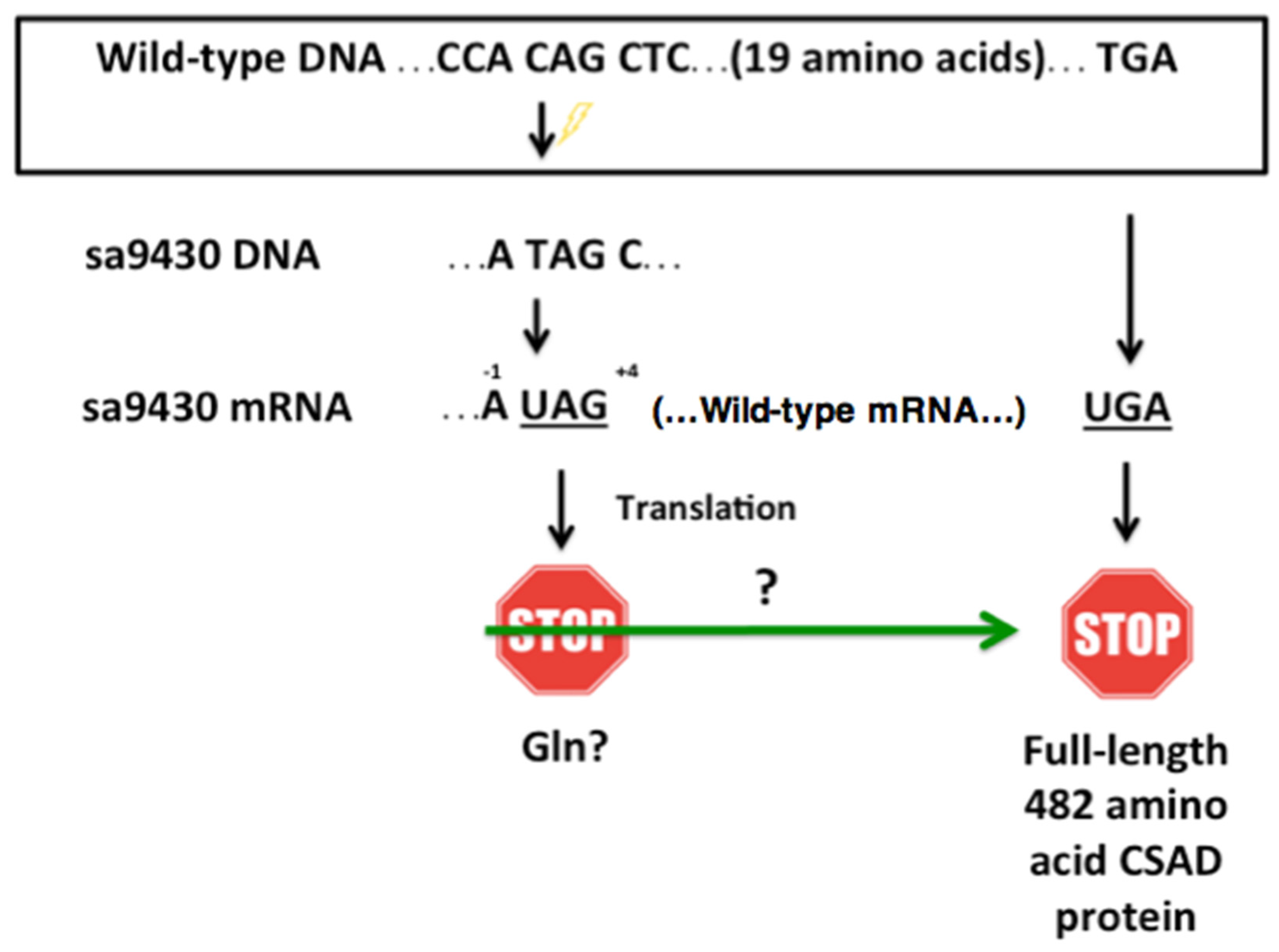
2.5. Nucleotides That Border the UAG May Increase Readthrough Frequency
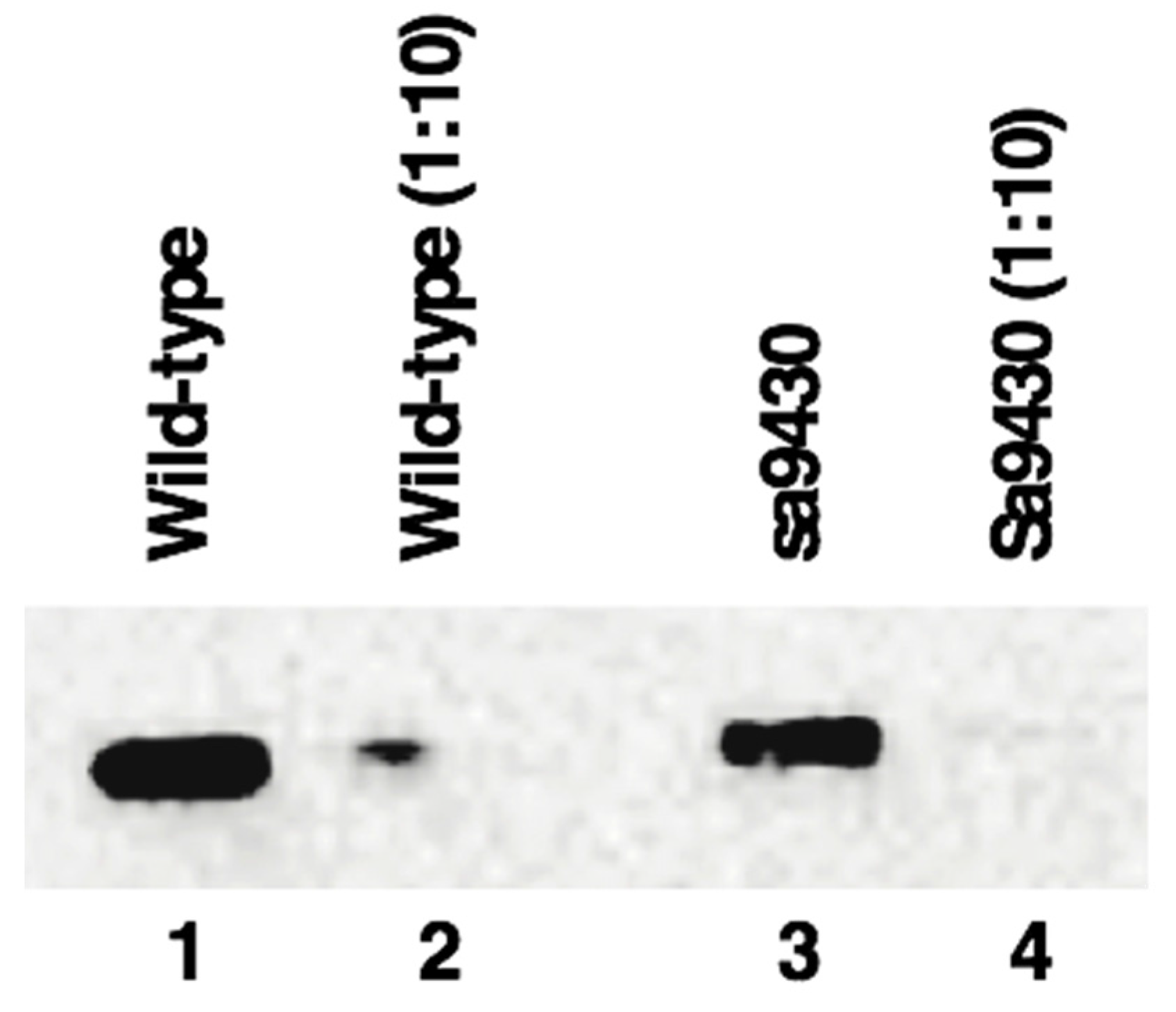
2.6. sa9430 CSAD Is the Same Size as the Wild-Type CSAD
2.7. Wild-Type and sa9430 CSAD Enzymes Catalyze the Conversion of Cysteic Acid to Taurine at Comparable Rates
2.8. Mass Spectrometry of Tryptic Peptides Indicates sa9430 Is Producing A Wild-Type CSAD Protein
3. Discussion
4. Materials and Methods
4.1. Computer Modeling of CSAD
4.2. Zebrafish Strains and Maintenance
4.3. Genotyping
- Forward: 5’ ACGTGGCGCCAGTCATTAAA 3’
- Reverse: 5’ GATGCCAATCGTTTGACCAGT 3’
4.4. Zebrafish Feeding Trials with and Without Supplemental Taurine
4.5. Liver Protein Isolation
4.6. Immunoblotting
4.7. CSAD Activity Assay
4.8. Amino Acid Analysis by HPLC
4.9. Preparation of Embryos for Amino Acid Analysis by HPLC
4.10. Preparation of Juveniles for Amino Acid Analysis by HPLC
4.11. Preparation of Paramecia and Artemia for Taurine Analysis
4.12. Preparation of GEMMA (Skretting) Feeds for HPLC Analysis
4.13. Mass Spectrometry
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ripps, H.; Shen, W. Review: Taurine: A “very Essential” Amino Acid. Mol. Vis. 2012, 18, 2673–2686. [Google Scholar] [PubMed]
- Knopf, K.; Sturman, J.A.; Armstrong, M.; Hayes, K.C. Taurine: An Essential Nutrient for the Cat. J. Nutr. 1978, 108, 773–778. [Google Scholar] [PubMed]
- Verbrugghe, A.; Bakovic, M. Peculiarities of One-Carbon Metabolism in the Strict Carnivorous Cat and the Role in Feline Hepatic Lipidosis. Nutrients 2013, 5, 2811–2835. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.M.; Barrows, F.T.; Place, A.R. Taurine Supplementation of Plant Derived Protein and n-3 Fatty Acids are Critical for Optimal Growth and Development of Cobia, Rachycentron Canadum. Lipids 2013, 48, 899–913. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Murata, H.; Goto, T.; Hatate, H.; Endo, M.; Yamashita, H.; Miyatake, H.; Ukawa, M. Role of Taurine Deficiency in Inducing Green Liver Symptom and Effect of Dietary Taurine Supplementation in Improving Growth in Juvenile Red Sea Bream Pagrus major Fed Non-Fishmeal Diets Based on Soy Protein Concentrate. Fish. Sci. 2011, 77, 235–244. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Endo, M.; Yamashita, H.; Ukawa, M. Taurine is an Essential Nutrient for Yellowtail Seriola quinqueradiata Fed Non-Fish Meal Diets Based on Soy Protein Concentrate. Aquaculture 2008, 280, 198–205. [Google Scholar] [CrossRef]
- Salze, G.P.; Davis, D.A. Taurine: A Critical Nutrient for Future Fish Feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- NOAA Fisheries. Nutrient Approval Is Victory for U.S. Fish Farmers and Seafood Lovers. Available online: http://www.nmfs.noaa.gov/stories/2017/02/taurine-QA.html (accessed 17 February 2017).
- Park, E.; Park, S.Y.; Dobkin, C.; Schuller-Levis, G. Development of a Novel Cysteine Sulfinic Acid Decarboxylase Knockout Mouse: Dietary Taurine Reduces Neonatal Mortality. J. Amino Acids 2014, 2014, 346809. [Google Scholar] [CrossRef] [PubMed]
- Thisse, B.; Thisse, C. Fast Release Clones: A High Throughput Expression Analysis. ZFIN Direct Data Submiss. (http://zfin. org). 2004. [Google Scholar]
- Chang, Y.C.; Ding, S.T.; Lee, Y.H.; Wang, Y.C.; Huang, M.F.; Liu, I.H. Taurine Homeostasis Requires De Novo Synthesis Via Cysteine Sulfinic Acid Decarboxylase during Zebrafish Early Embryogenesis. Amino Acids 2013, 44, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, D.J.; Chen, Z.; Zhuang, L.; Fei, Y.J.; Navarre, S.; Ganapathy, V. Molecular Characterization and Expression Pattern of Taurine Transporter in Zebrafish during Embryogenesis. Life Sci. 2008, 82, 1004–1011. [Google Scholar] [CrossRef]
- Cetiner, M.; Sener, G.; Sehirli, A.O.; Eksioglu-Demiralp, E.; Ercan, F.; Sirvanci, S.; Gedik, N.; Akpulat, S.; Tecimer, T.; Yegen, B.C. Taurine Protects Against Methotrexate-Induced Toxicity and Inhibits Leukocyte Death. Toxicol. Appl. Pharmacol. 2005, 209, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Cha, Y.N. Taurine Chloramine Produced from Taurine Under Inflammation Provides Anti-Inflammatory and Cytoprotective Effects. Amino Acids 2014, 46, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Busch-Nentwich, E.; Kettleborough, R.; Dooley, C.M.; Scahill, C.; Sealy, I.; White, R.; Herd, C.; Mehroke, S.; Wali, N.; Carruthers, S.; et al. Sanger institute zebrafish mutation project mutant data submission. ZFIN Direct Data Submiss. 2013. [Google Scholar]
- Andrews, J.; Kuck, J. The Conversion of Cysteic Acid to Taurine. J. Elisha Mitchell Sci. Soc. 1963, 69, 35–41. [Google Scholar]
- Edgar, S.; Hickman, M.; Marsden, M.; Morris, J.; Rogers, Q. Dietary Cysteic Acid Serves as a Precursor of Taurine for Cats. Am. Inst. Nutr. 1994, 124, 103–109. [Google Scholar]
- Vitvitsky, V.; Garg, S.K.; Banerjee, R. Taurine Biosynthesis by Neurons and Astrocytes. J. Biol. Chem. 2011, 286, 32002–32010. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.M. Taurine: An Indispensible Ingredient in the Development of Sustainable Aquafeeds. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2013. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Cassan, M.; Rousset, J.P. UAG Readthrough in Mammalian Cells: Effect of Upstream and Downstream Stop Codon Contexts Reveal Different Signals. BMC Mol. Biol. 2001, 2, 3. [Google Scholar] [CrossRef]
- Brown, C.M.; Stockwell, P.A.; Trotman, C.N.; Tate, W.P. Sequence Analysis Suggests that Tetra-Nucleotides Signal the Termination of Protein Synthesis in Eukaryotes. Nucleic Acids Res. 1990, 18, 6339–6345. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Leszyk, J.D.; Mangus, D.A.; Jacobson, A. Nonsense Suppression by Near-Cognate tRNAs Employs Alternative Base Pairing at Codon Positions 1 and 3. Proc. Natl. Acad. Sci. USA 2015, 112, 3038–3043. [Google Scholar] [CrossRef] [PubMed]
- Beznoskova, P.; Wagner, S.; Jansen, M.E.; von der Haar, T.; Valasek, L.S. Translation Initiation Factor eIF3 Promotes Programmed Stop Codon Readthrough. Nucleic Acids Res. 2015, 43, 5099–5111. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, S.; Cornu, D.; Argentini, M.; Namy, O. New Insights into the Incorporation of Natural Suppressor tRNAs at Stop Codons in Saccharomyces Cerevisiae. Nucleic Acids Res. 2014, 42, 10061–10072. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.X.; Copeland, T.D.; Oroszlan, S.; Rein, A.; Levin, J.G. Identification of Amino Acids Inserted during Suppression of UAA and UGA Termination Codons at the Gag-Pol Junction of Moloney Murine Leukemia Virus. Proc. Natl. Acad. Sci. USA 1990, 87, 8860–8863. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, M.; Xu, J.; Luo, J. Relationships among Stop Codon Usage Bias, its Context, Isochores, and Gene Expression Level in various Eukaryotes. J. Mol. Evol. 2005, 61, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Yi, S.; Guo, X.; Wang, W. A Comprehensive Analysis of Codon Usage Patterns in Blunt Snout Bream (Megalobrama Amblycephala) Based on RNA-Seq Data. Int. J. Mol. Sci. 2015, 16, 11996–12013. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.M.; Tuohy, T.M.; Mosurski, K.R. Codon Usage in Yeast: Cluster Analysis Clearly Differentiates Highly and Lowly Expressed Genes. Nucleic Acids Res. 1986, 14, 5125–5143. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miyauchi, K.; Suzuki, T.; Yokobori, S.; Shigi, N.; Kondow, A.; Takeuchi, N.; Yamagishi, A.; Watanabe, K. Taurine-Containing Uridine Modifications in tRNA Anticodons are Required to Decipher Non-Universal Genetic Codes in Ascidian Mitochondria. J. Biol. Chem. 2011, 286, 35494–35498. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Suzuki, T.; Wada, T.; Saigo, K.; Watanabe, K. Taurine as a Constituent of Mitochondrial tRNAs: New Insights into the Functions of Taurine and Human Mitochondrial Diseases. EMBO J. 2002, 21, 6581–6589. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Kim, H.K.; Kim, W.B.; Yang, J.; Kim, B.K. Protective Effect of Taurine on Indomethacin-Induced Gastric Mucosal Injury. Adv. Exp. Med. Biol. 1996, 403, 147–155. [Google Scholar] [PubMed]
- Wass, M.N.; Kelley, L.A.; Sternberg, M.J. 3DLigandSite: Predicting Ligand-Binding Sites using Similar Structures. Nucleic Acids Res. 2010, 38, W469–W473. [Google Scholar] [CrossRef] [PubMed]
- Spicer, O.S.; Wong, T.T.; Zmora, N.; Zohar, Y. Targeted Mutagenesis of the Hypophysiotropic Gnrh3 in Zebrafish (Danio Rerio) Reveals no Effects on Reproductive Performance. PLoS One 2016, 11, e0158141. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.M.; Barrows, F.T.; Place, A.R. Leaching of Taurine from Commercial Type Aquaculture Feeds. Aquacult. Res. 2015, 46, 1510–1517. [Google Scholar] [CrossRef]
- Yokoyama, M.; Takeuchi, T.; Park, G.S.; Nakazoe, J. Hepatic Cysteinesulphinate Decarboxylase Activity in Fish. Aquacult. Res. 2001, 32, 216. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Farrah, T.; Lam, H.; Tasman, N.; Sun, Z.; Nilsson, E.; Pratt, B.; Prazen, B.; et al. A Guided Tour of the Trans-Proteomic Pipeline. Proteomics 2010, 10, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Eng, J.K.; Jahan, T.A.; Hoopmann, M.R. Comet: An Open-Source MS/MS Sequence Database Search Tool. Proteomics 2013, 13, 22–24. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | pmol Taurine/h·μg Protein |
|---|---|
| Wild-type | 5.83 ± 1.94 |
| sa9430 | 3.09 ± 0.45 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larkin, M.E.M.; Place, A.R. Running the Stop Sign: Readthrough of a Premature UAG Termination Signal in the Translation of a Zebrafish (Danio rerio) Taurine Biosynthetic Enzyme. Mar. Drugs 2017, 15, 162. https://doi.org/10.3390/md15060162
Larkin MEM, Place AR. Running the Stop Sign: Readthrough of a Premature UAG Termination Signal in the Translation of a Zebrafish (Danio rerio) Taurine Biosynthetic Enzyme. Marine Drugs. 2017; 15(6):162. https://doi.org/10.3390/md15060162
Chicago/Turabian StyleLarkin, Mary E.M., and Allen R. Place. 2017. "Running the Stop Sign: Readthrough of a Premature UAG Termination Signal in the Translation of a Zebrafish (Danio rerio) Taurine Biosynthetic Enzyme" Marine Drugs 15, no. 6: 162. https://doi.org/10.3390/md15060162
APA StyleLarkin, M. E. M., & Place, A. R. (2017). Running the Stop Sign: Readthrough of a Premature UAG Termination Signal in the Translation of a Zebrafish (Danio rerio) Taurine Biosynthetic Enzyme. Marine Drugs, 15(6), 162. https://doi.org/10.3390/md15060162