The Sea Urchin Arbacia lixula: A Novel Natural Source of Astaxanthin

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
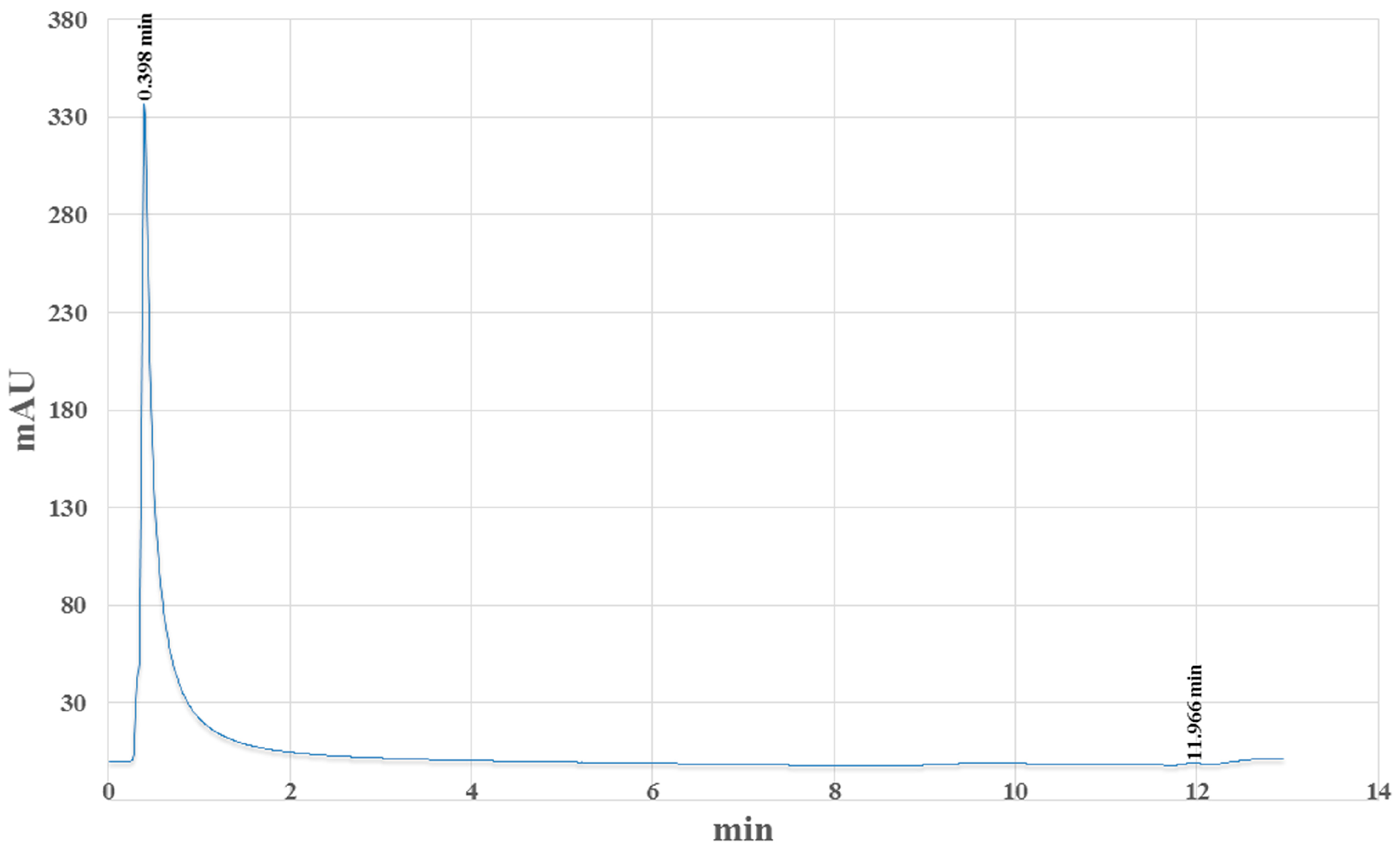
2.1. Chemical Analysis
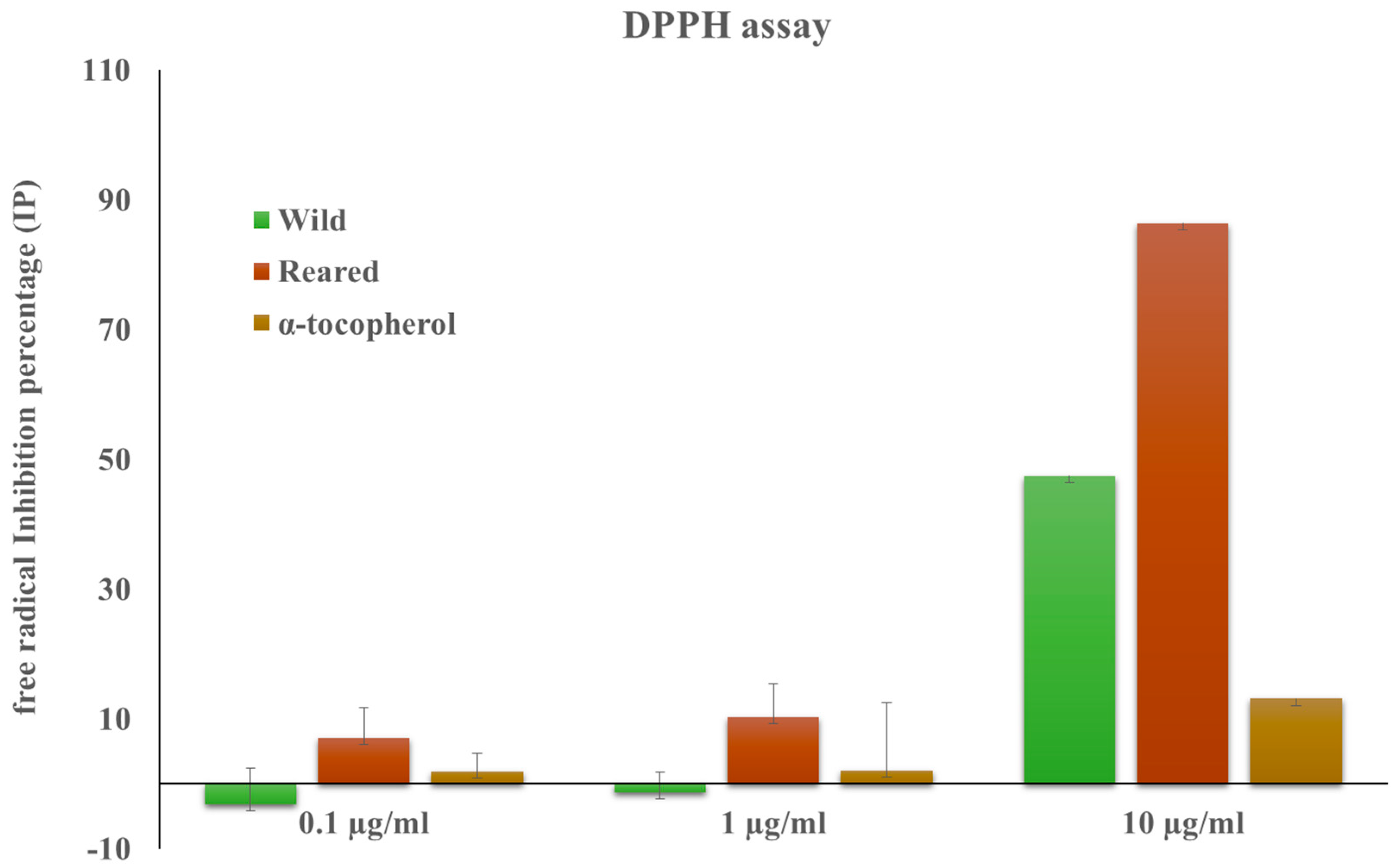
2.2. Radical Scavenging Activity
3. Discussion
4. Materials and Methods
4.1. Culture System
4.2. Feeding Practice
4.3. Eggs Collection
4.4. Chemical Extraction from Eggs
4.5. HPLC Analysis
4.6. DPPH-Assay
4.7. Experimental Design and Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bendich, A. Recent advances in clinical research involving carotenoids. Pure Appl. Chem. 1994, 66, 1017–1024. [Google Scholar] [CrossRef]
- Krinsky, N.I. The biological properties of carotenoids. Pure Appl. Chem. 1994, 66, 1003–1010. [Google Scholar] [CrossRef]
- Kelly, M.S.; Symonds, R.C. Carotenoids in sea urchins. In Sea Urchin: Biology and Ecology, 3rd ed.; Lawrence, J.M., Ed.; Elsevier Science Publishing Co Inc.: San Diego, CA, USA, 2013; pp. 171–177. [Google Scholar]
- Goodwin, T.W. The Biochemistry of the Carotenoids, 2nd ed.; Chapman and Hall: London, UK, 1984; Volume 2. [Google Scholar]
- Haug, T.; Kjuul, A.K.; Styrvold, O.B.; Sandsdalen, E.; Olsen, M.O.; Stensvåg, K. Antibacterial activity in Strongylocentrotus droebachiensis (Echinoidea), Cucumaria frondosa (Holothuroidea), and Asterias rubens (Asteroidea). J. Invert. Pathol. 2002, 81, 94–102. [Google Scholar] [CrossRef]
- Kuwahara, R.; Hatate, H.; Yuki, T.; Murata, H.; Tanaka, R.; Hama, Y. Antioxidant property of polyhydroxylated naphthoquinone pigments from shells of purple sea urchin Anthocidaris crassispina. Lwt-Food Sci. Technol. 2009, 42, 1296–1300. [Google Scholar] [CrossRef]
- Uma, B.; Parvathavarthini, R. Antibacterial effect of hexane extract of sea Urchin, Temnopleurus alexandri (Bell, 1884). Int. J. PharmTech Res. 2010, 2, 1677–1680. [Google Scholar]
- Abubakar, L.; Mwangi, C.; Uku, J.; Ndirangu, S. Antimicrobial activity of various extracts of the sea urchin Tripneustes gratilla (Echinoidea). Afr. J. Pharmacol. Ther. 2012, 1, 19–23. [Google Scholar]
- Artyukov, A.A.; Popova, A.M.; Tsybulskyb, A.V.; Krivoshapkoa, O.N.; Polyakovac, N.V. Pharmacological activity of Echinochrome A alone and in the biologically active additive Timarin. Biomed. Chem. 2013, 7, 237–242. [Google Scholar] [CrossRef]
- Bragadeeswaran, S.; Sri Kumaran, N.; Prasath Sankar, P.; Prabahar, R. Bioactive potential of sea urchin Temnopleurus toreumaticus from Devanampattinam, Southeast coast of India. J. Pharm. Altern. Med. 2013, 2, 9–17. [Google Scholar]
- Lennikov, A.; Kitaichi, N.; Noda, K.; Mizuuchi, K.; Ando, R.; Dong, Z.; Fukuhara, J.; Kinoshita, S.; Namba, K.; Ohno, S.; et al. Amelioration of endotoxin-induced uveitis treated with the sea urchin pigment echinochrome in rats. Mol. Vis. 2014, 20, 171–177. [Google Scholar] [PubMed]
- Suleria, H.R.; Osborne, S.; Masci, P.; Gobe, G. Marine-Based Nutraceuticals: An Innovative Trend in the Food and Supplement Industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Levitskaya, E.L.; Tikhonova, E.V.; Ivanova, M.V. Antioxidant properties, autoxidation and mutagenic activity of echinochrome A compared with its etherified derivative. Biochemistry 2001, 66, 885–893. [Google Scholar] [PubMed]
- Lederer, E. Echinenone and pentaxanthin; two new carotenoids in the sea urchin (Echinus esculentus). C. R. Acad. Sci. Paris 1935, 201, 300–302. [Google Scholar]
- Goodwin, T.W.; Taha, M.M. The carotenoids of the gonads of the limpets Patella vulgata and Patella depressa. Biochem. J. 1950, 47, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C. Seasonal changes in the biochemical composition of the edible sea urchin Paracentrotus lividus (Echinodermata: Echinoidea) in a lagoonal environment. Mar. Ecol. 1998, 1, 1–11. [Google Scholar] [CrossRef]
- Lawrence, J.M. Edible Sea Urchins: Biology and Ecology; Elsevier: Boston, MA, USA, 2007; p. 380. [Google Scholar]
- Hávardsson, B.; Imsland, A.K.; Christiansen, R. The effect of astaxanthin in feed and environmental temperature on carotenoid concentration in the gonads of green sea urchin Strongylocentrotus droebachiensis Muller. J. World Aquac. Soc. 1999, 30, 208–218. [Google Scholar] [CrossRef]
- McLaughlin, G.; Kelly, M.S. Effect of artificial diets containing carotenoid-rich microalgae on gonad growth and color in the sea urchin Psammechinus miliaris (Gmelin). J. Shellfish Res. 2001, 20, 377–382. [Google Scholar]
- Robinson, S.M.C.; Castell, J.D.; Kennedy, E.J. Developing suitable color in the gonads of cultured green sea urchins (Strongylocentrotus droebachiensis). Aquaculture 2002, 206, 289–303. [Google Scholar] [CrossRef]
- Robinson, S.M.; Lawrence, J.M.; Burridge, L.; Haya, K.; Martin, J.; Castell, J.; Lawrence, A.L. Effectiveness of different pigment sources in coloring the gonads of the green sea urchin (Strongylocentrotus droebachiensis). In Sea Urchins: Fisheries and Ecology; Lawrence, J.M., Guzmán, O., Eds.; DEStech Publication: Lancaster, UK, 2004; pp. 215–221. [Google Scholar]
- Shpigel, M.; McBride, S.C.; Marciano, S.S.; Ben-Amotz, A. Improving gonad colour and somatic index in the European sea urchin Paracentrotus lividus. Aquaculture 2005, 245, 101–109. [Google Scholar] [CrossRef]
- Shpigel, M.; Schlosser, S.; Ben-Amotz, A.; Lawrence, A.; Lawrence, J. Effects of dietary carotenoid on the gut and the gonad of the sea urchin Paracentrotus lividus. Aquaculture 2006, 261, 1269–1280. [Google Scholar] [CrossRef]
- Suckling, C.; Symonds, R.C.; Kelly, M.S.; Young, A.J. The effect of artificial diets on gonad color and biomass in the edible sea urchin Psammechinus miliaris. Aquaculture 2011, 318, 335–342. [Google Scholar] [CrossRef]
- Matsuno, T.; Tsushima, M. Carotenoids in sea urchins. In Edible Sea Urchins: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2001; pp. 115–138. [Google Scholar]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health promoting effects of astaxanthin, a high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Grimmig, B.; Kim, S.; Nash, K.; Bickford, P.C.; Shytle, R.D. Neuroprotective mechanisms of astaxanthin: A potential therapeutic role in preserving cognitive function in age and neurodegeneration. GeroScience 2017, 39, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Phang, S.M.; Sarada, R.; Ravishankar, G.A. Astaxanthin: Sources, extraction, stability, biological activities and its commercial application—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2016, 128, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, O.; Imperatore, C.R.; Tomas, V.; Costantino, V.; Saggiomo, V.; Mangoni, A. The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae. Mar. Drugs 2011, 9, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Lessios, H.A.; Lockhart, S.; Collin, R.; Sotil, G.; Sanchez-Jerez, P.; Zigler, K.S.; Perez, A.F.; Garrido, M.J.; Geyer, L.B.; Bernardi, G.; et al. Phylogeography and bindin evolution in Arbacia, a sea urchin genus with an unusual distribution. Mol. Ecol. 2012, 21, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Gianguzza, P.; Agnetta, D.; Bonaviri, C.; Di Trapani, F.; Visconti, G.; Gianguzza, F.; Riggio, S. The rise of thermophilic sea urchins and the expansion of barren grounds in the Mediterranean Sea. Chem. Ecol. 2011, 27, 129–134. [Google Scholar] [CrossRef]
- Agnetta, D.; Bonaviri, C.; Badalamenti, F.; Scianna, C.; Vizzini, S.; Gianguzza, P. Functional traits of two co-occurring sea urchins across a barren/forest patch system. J. Sea Res. 2013, 76, 170–177. [Google Scholar] [CrossRef]
- Tsushima, M.; Matsuno, T. Comparative biochemical studies of carotenoids in sea urchins. Comp. Biochem. Physiol. 1990, 96, 801–810. [Google Scholar] [CrossRef]
- Symonds, R.C.; Caris-Veyrat, C.; Kelly, M.S.; Young, A.J. Carotenoids in the sea urchin Paracentrotus lividus: Occurrence of 9′-cis echinenone as the dominant carotenoid in gonad colour determination. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 148, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Symonds, R.C.; Kelly, M.S.; Suckling, C.C.; Young, A.J. Carotenoids in the gonad and gut of the edible sea urchin Psammechinus miliaris. Aquaculture 2009, 288, 120–125. [Google Scholar] [CrossRef]
- Steinbrenner, J.; Linden, H. Regulation of two carotenoid biosynthesis genes coding for phytoene synthase and carotenoid hydroxylase during stress-induced astaxanthin formation in the green alga Haematococcus pluvialis. Plant Physiol. 2001, 125, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, T. The biosynthetic pathway of carotenoids in the astaxanthin-producing green alga Chlorella zofingiensis. World J. Microbiol. Biotechnol. 2008, 24, 2927–2932. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in Marine Animals. Mar Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Kuwahara, T.; Narita, M. Carotenoids of sea angels Clione limacina and Paedoclione doliiformis from the perspective of the food chain. Mar. Drugs 2014, 12, 1460–1470. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, J.; Nagata, S.; Iwahashi, M.; Koike, T.; Okada, M. Intensification of color of fancy red carp with zeaxanthin and microxanthophyll, major carotenoid constituents of Spirulina. Bull. Jpn. Soc. Sci. Fish 1979, 45, 627–632. [Google Scholar] [CrossRef]
- Jalal, K.C.A.; Ambak, M.A.; Haji-Hassan, T.; Sorgeloos, P.; Mohd Zaki, S.; Saad, C.R. Effects of Spirulina on the growth and carotenoid deposition in tropical sport fish Tor tombroides fry. In European Aquaculture Society Special Publication; No. 30; European Aquaculture Society: Oostende, Belgium, 2001; pp. 258–261. [Google Scholar]
- Nandeesha, M.C.; Gangadhara, B.; Manissery, J.K.; Venkataraman, L.V. Growth performance of two Indian major carps, catla (Catla catla) and rohu (Labeo rohita) fed diets containing different levels of Spirulina platensis. Bioresour. Technol. 2001, 80, 117–120. [Google Scholar] [CrossRef]
- Gouveia, L.; Rema, P.; Pereira, O.; Empis, J. Colouring ornamental fish (Cyprinus carpio and Carassius auratus) with microalgal biomass. Aquac. Nutr. 2003, 9, 123–129. [Google Scholar] [CrossRef]
- Liao, W.L.; Nur-E-Borhan, S.A.; Shigera, O.; Matsui, T.; Yamaguchi, K. Pigmentation of cultured black tiger prawn by feeding with Spirulina supplementation diet. Nippon Suisan Gakkai Shi 1993, 59, 165–169. [Google Scholar] [CrossRef]
- Chien, Y.H.; Shiau, W.C. The effects of Haematococcus pluvialis, Spirulina pacific and synthetic astaxanthin on the pigmentation, survival, growth and oxygen consumption of kuruma prawn Penaeus japonicus Bate. In Book of Abstracts of World Aquaculture; World Aquaculture Society: Baton Rouge, LA, USA, 1998; p. 156. [Google Scholar]
- Regunathan, C.; Wesley, S.G. Pigment deficiency correction in shrimp broodstock using Spirulina as a carotenoid source. Aquac. Nutr. 2006, 12, 425–432. [Google Scholar] [CrossRef]
- Ghaeni, M.; Roomiani, L.; Moradi, Y. Evaluation of Carotenoids and Chlorophyll as Natural Resources for Food in Spirulina Microalgae. Appl. Food Biotechnol. 2015, 2, 39–44. [Google Scholar]
- Tanaka, Y.; Matsuguchi, H.; Katayama, T.; Simpson, K.L.; Chichester, C.O. The biosynthesis of astaxanthin. XVIII. The metabolism of the carotenoids in the prawn, Peneus japonicus Bate. Bull. Jpn. Soc. Sci. Fish 1976, 42, 197–202. [Google Scholar] [CrossRef]
- Perry, A.; Rasmussen, H.; Johnson, E.J. Xanthophyll (Lutein, Zeaxanthin) content in fruits, vegetables and corn and egg products. J. Food Compos. Anal. 2009, 22, 9–15. [Google Scholar] [CrossRef]
- Adb El Baky, H.; El Baz, F.K.; Baroty, L. Evaluation of Marine Alga Ulva lactuca L. as A Source of Natural Preservative Ingredient. Electron. J. Environ. Agric. Food Chem. 2008, 3, 434–444. [Google Scholar]
- Razi Parjikolaei, B.; Cardoso, L.C.; Fernandez-Ponce, M.T.; Serano, C.M.; Bruhn, A.; Fretté, X.; Christensen, K.V. Supercritical fluid extraction of carotenoids from Ulva lactuca (Chlorophyta). Planta Med. 2014, 80, 151–152. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The green microalga Tetraselmis suecica reduces oxidative stress and induces repairing mechanisms in human cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Nuzzo, G.; Ianora, A.; Luongo, E.; Romano, G.; Gallo, C.; Sansone, C.; Aprea, S.; Mancini, F.; D’Oro, U.; et al. Development and Application of a Novel SPE-Method for Bioassay-Guided Fractionation of Marine Extracts. Mar. Drugs 2015, 13, 5736–5749. [Google Scholar] [CrossRef] [PubMed]
- Brunet, C.; Chandrasekaran, R.; Barra, L.; Giovagnetti, V.; Corato, F.; Ruban, A.V. Spectral radiation dependent photoprotective mechanism in the diatom Pseudo-nitzschia multistriata. PLoS ONE 2014, 9, e87015. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.W.; Wright, S.W. Qualitative and quantitative HPLC analysis of SCOR reference algal cultures. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 343–360. [Google Scholar]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Regression Models, 4th ed.; McGraw-Hill: New York, NY, USA, 1996. [Google Scholar]
- Rouxton, G.D.; Beauchamps, G. Time for some a priori thinking about post hoc testing. Behav. Ecol. 2008, 19, 690–693. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Egger, M.; Smith, G.D. Investigating and dealing with publication and other biases in meta-analysis. BMJ 2001, 323, 101. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Wild (n = 5) | Farmed (n = 5) |
| 1.5 ± 1.8 µg/mg | 27.0 ± 7.5 µg/mg |
| Mean Fresh Weight of Eggs | Mean Dry Weight of Eggs | Astaxanthin Weight per g of Dry Eggs |
| 5.5 ± 1.5 g | 0.5 ± 0.250 g | 0.014 ± 0.004 g |
| Main Effects | SS | df | MS | F | p | G-G e | G-G p | H-F e | H-F p | |
|---|---|---|---|---|---|---|---|---|---|---|
| Wild vs. Reared | WvsR | 2313.76 | 1 | 2313.76 | 78.80 | 2.05 × 10−5 | ||||
| Error | 234.91 | 8 | 29.36 | |||||||
| Concentration | C | 31,520.68 | 3 | 10,506.89 | 430.92 | <0.01 × 10−15 | 0.72 | 1.11 × 10−15 | 1.00 | <0.01 × 10−15 |
| C × WvsR | 2082.90 | 3 | 694.30 | 28.48 | 4.46 × 10−8 | 0.72 | 2.46 × 10−6 | 1.00 | 4.46 × 10−8 | |
| Error | 585.18 | 24 | 24.38 | |||||||
| Planned Comparisons within R | SS | df | MS | F | p |
|---|---|---|---|---|---|
| L vs. C | 120.37 | 1 | 120.37 | 11.92 | 0.0087 |
| Error | 80.81 | 8 | 10.10 | ||
| M vs. C | 259.28 | 1 | 259.28 | 21.55 | 0.0017 |
| Error | 96.27 | 8 | 12.03 | ||
| H vs. C | 18,608.15 | 1 | 18,608.15 | 639.05 | 6.42 × 10−9 |
| Error | 232.95 | 8 | 29.12 | ||
| Planned Comparisons within W | SS | df | MS | F | p |
| L vs. C | 27.84 | 1 | 27.84 | 2.76 | 0.1355 |
| Error | 80.81 | 8 | 10.10 | ||
| M vs. C | 5.01 | 1 | 5.01 | 0.42 | 0.5367 |
| Error | 96.27 | 8 | 12.03 | ||
| H vs. C | 5594.73 | 1 | 5594.73 | 192.14 | 7.10 × 10−7 |
| Error | 232.95 | 8 | 29.12 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirino, P.; Brunet, C.; Ciaravolo, M.; Galasso, C.; Musco, L.; Vega Fernández, T.; Sansone, C.; Toscano, A. The Sea Urchin Arbacia lixula: A Novel Natural Source of Astaxanthin. Mar. Drugs 2017, 15, 187. https://doi.org/10.3390/md15060187
Cirino P, Brunet C, Ciaravolo M, Galasso C, Musco L, Vega Fernández T, Sansone C, Toscano A. The Sea Urchin Arbacia lixula: A Novel Natural Source of Astaxanthin. Marine Drugs. 2017; 15(6):187. https://doi.org/10.3390/md15060187
Chicago/Turabian StyleCirino, Paola, Christophe Brunet, Martina Ciaravolo, Christian Galasso, Luigi Musco, Tomás Vega Fernández, Clementina Sansone, and Alfonso Toscano. 2017. "The Sea Urchin Arbacia lixula: A Novel Natural Source of Astaxanthin" Marine Drugs 15, no. 6: 187. https://doi.org/10.3390/md15060187
APA StyleCirino, P., Brunet, C., Ciaravolo, M., Galasso, C., Musco, L., Vega Fernández, T., Sansone, C., & Toscano, A. (2017). The Sea Urchin Arbacia lixula: A Novel Natural Source of Astaxanthin. Marine Drugs, 15(6), 187. https://doi.org/10.3390/md15060187