Characterization and Antioxidant and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Gelatin Hydrolysates Prepared from Extrusion-Pretreated Milkfish (Chanos chanos) Scale

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
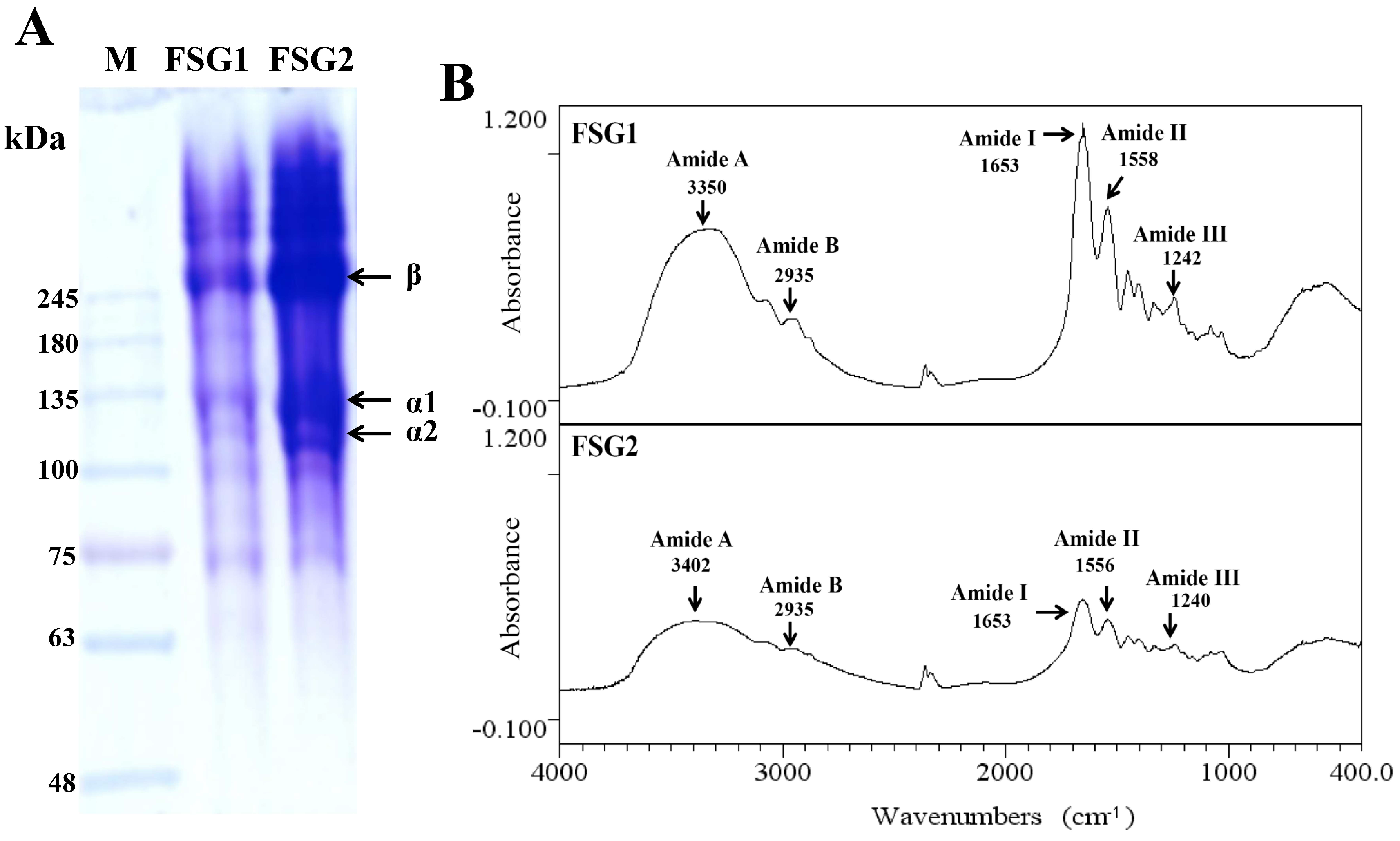
2.1. Preparation of Gelatins (FSG1 and FSG2) from Extrusion-Pretreated Milkfish Scale
2.2. Physicochemical Properties of Extracted Gelatins for FSG1 and FSG2
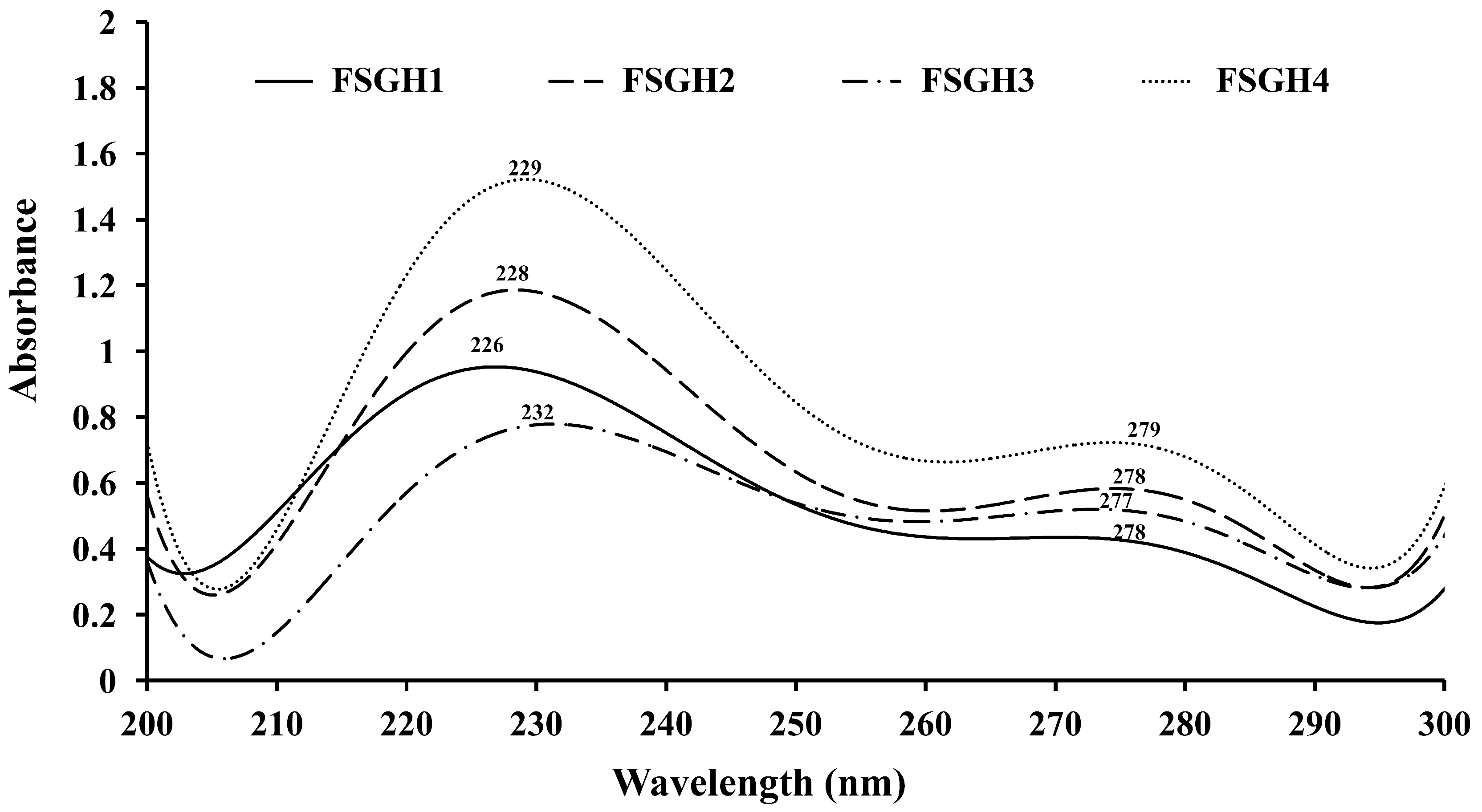
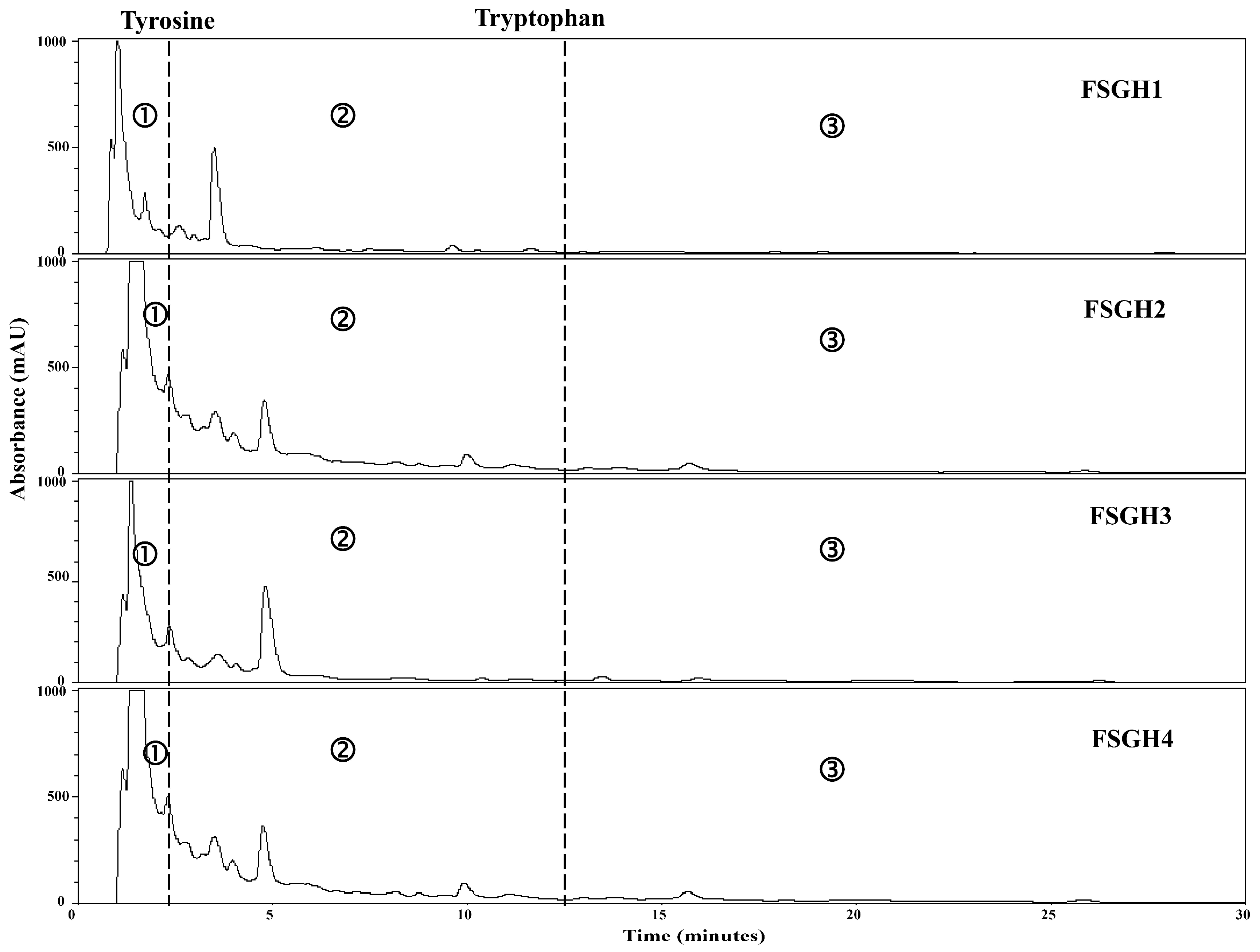
2.3. Preparation of Gelatin Hydrolysates (FSGH1, FSGH2, FSGH3, and FSGH4) from FSG1 and FSG2, and Characteristics of FSGH1, FSGH2, FSGH3, and FSGH4
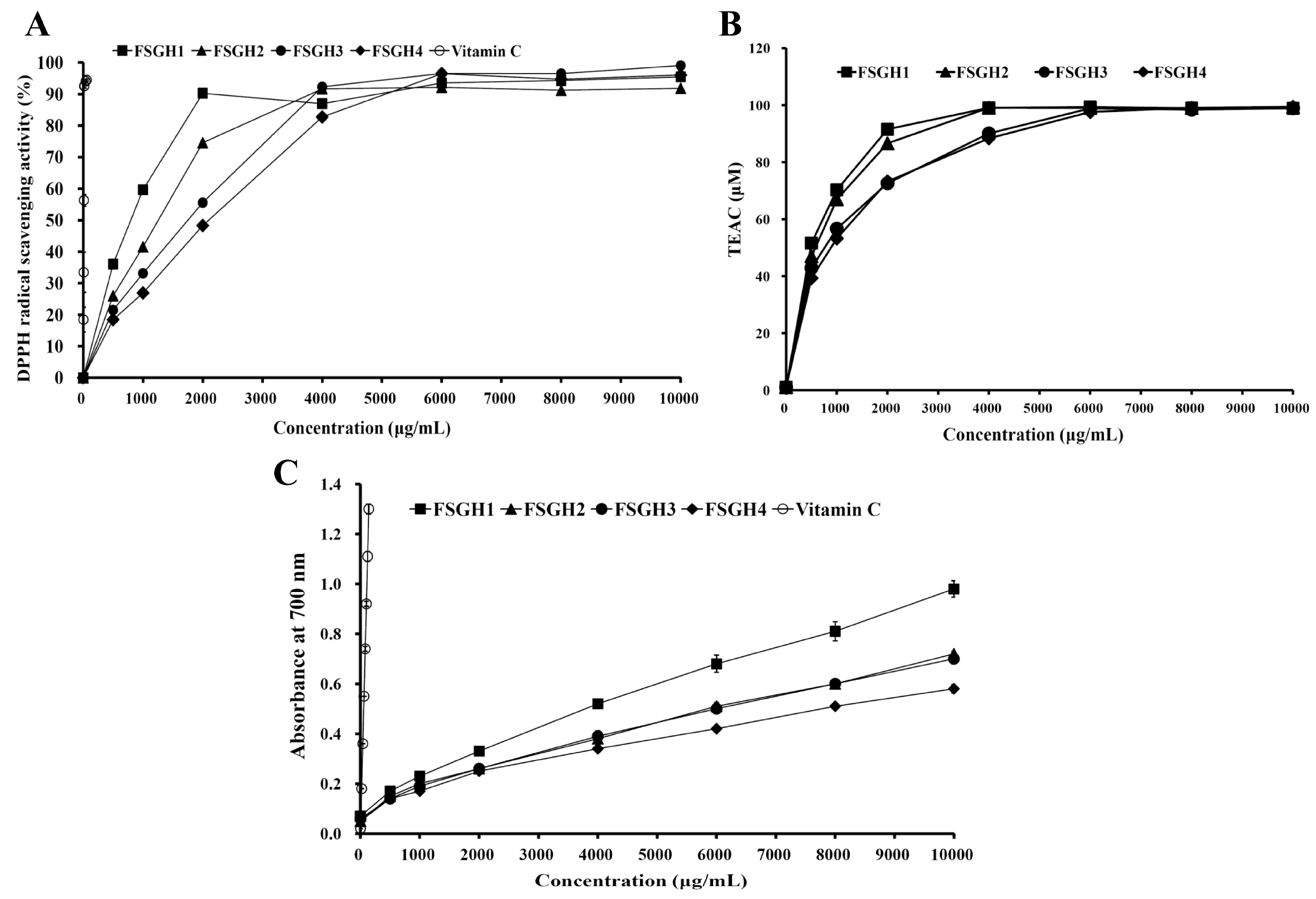
2.4. Antioxidant Activities of FSGH1, FSGH2, FSGH3, and FSGH4
2.5. ACE Inhibition Activity of FSGH1,FSGH2, FSGH3, and FSGH4
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Extrusion-Cooking Procedure
3.3. Determination of Protein Concentration
3.4. Extraction of Gelatin from Non-extruded Fish Scale and Fish Scale Extrudate
3.5. Chemical Composition Analyses
3.6. SDS-PAGE
3.7. Fourier Transform Infrared (FTIR) Spectroscopy
3.8. Preparation of Gelatin Hydrolysates
3.9. Evaluation of Degree of Hydrolysis
3.10. Determination of Amino acid Composition
3.11. Ultraviolet (UV) Absorption Spectrum
3.12. Molecular Weight Analysis
3.13. Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC)
3.14. LC-ESI/MS/MS and Data Analyses
3.15. DPPH Radical Scavenging Activity
3.16. ABTS Radical Scavenging Activity
3.17. Reducing Power Assay
3.18. ACE-Inhibiting Activity
3.19. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, K.W.; Lee, H.J. Biphasic effects of dietary antioxidants on oxidative stress-mediated carcinogenesis. Mech. Ageing Dev. 2006, 127, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A. Free radical biology: Xenobiotics, cancer, and aging. Ann. N. Y. Acad. Sci. 1982, 393, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Frölich, L.; Riederer, P. Free radical mechanisms in dementia of Alzheimer type and the potential for antioxidant treatment. Drug Res. 1995, 45, 443–446. [Google Scholar]
- Chedea, V.S.; Braicu, C.; Socaciu, C. Antioxidant/prooxidant activity of a polyphenolic grape seed extract. Food Chem. 2010, 121, 132–139. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Lin, L.; Lv, S.; Li, B. Angiotensin-I-converting enzyme (ACE)-inhibitory and antihypertensive properties of squid skin gelatin hydrolysates. Food Chem. 2012, 131, 225–230. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Ondetti, M.A.; Rubin, B.; Cushman, D.W. Design of specific inhibitors of angiotensin-converting enzyme: New class of orally active antihypertensive agents. Science 1977, 196, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Patchett, A.A.; Harris, E.; Tristram, E.W.; Wyvratt, M.J.; Wu, M.T.; Taub, D.; Peterson, E.R.; Ikeler, T.J.; Ten Broeke, J.; Payne, L.G. A new class of angiotensin-converting enzyme inhibitors. Nature 1980, 288, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Gavras, H.; Gavras, I. Angiotensin converting enzyme inhibitors. Properties and side effects. Hypertension 1988, 11, II-37–II-41. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Suzuki, N. Isolation of collagen from fish waste material—skin, bone and fins. Food Chem. 2000, 68, 277–281. [Google Scholar] [CrossRef]
- Pati, F.; Adhikari, B.; Dhara, S. Isolation and characterization of fish scale collagen of higher thermal stability. Bioresour. Technol. 2010, 101, 3737–3742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, S.; Wang, Z. Pre-treatment optimization and properties of gelatin from freshwater fish scales. Food Bioprod. Process. 2011, 89, 185–193. [Google Scholar] [CrossRef]
- Sae-leaw, T.; O’callaghan, Y.C.; Benjakul, S.; O’brien, N.M. Antioxidant, immunomodulatory and antiproliferative effects of gelatin hydrolysates from seabass (Lates calcarifer) skins. Int. J. Food Sci. Technol. 2016, 51, 1545–1551. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, Y.T.; Byun, H.G.; Nam, K.S.; Joo, D.S.; Shahidi, F. Isolation and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin. J. Agric. Food Chem. 2001, 49, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Kim, S.K. Antioxidant properties of a radical-scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate. J. Agric. Food Chem. 2005, 53, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C. Aquaculture in Taiwan; Blackwell Scientific Publications: Oxford, UK, 1990. [Google Scholar]
- Huang, C.Y.; Kuo, J.M.; Wu, S.J.; Tsai, H.T. Isolation and characterization of fish scale collagen from tilapia (Oreochromis sp.) by a novel extrusion-hydro-extraction process. Food Chem. 2016, 190, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Goda, T.; Takeuchi, N.; Einaga, H.; Tanabe, T. Preparation of collagen/calcium phosphate multilayer sheet using enzymatic mineralization. Biomaterials 2004, 25, 5481–5489. [Google Scholar] [CrossRef] [PubMed]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnacion, A.B.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Wangtueai, S.; Noomhorm, A. Processing optimization and characterization of gelatin from lizardfish (Saurida spp.) scales. LWT-Food Sci. Technol. 2009, 42, 825–834. [Google Scholar] [CrossRef]
- Ogawa, M.; Portier, R.J.; Moody, M.W.; Bell, J.; Schexnayder, M.A.; Losso, J.N. Biochemical properties of bone and scale collagens isolated from the subtropical fish black drum (Pogonia cromis) and sheepshead seabream (Archosargus probatocephalus). Food Chem. 2004, 88, 495–501. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Comparative study on characteristics of gelatin from the skins of brownbanded bamboo shark and blacktip shark as affected by extraction conditions. Food Hydrocoll. 2010, 24, 164–171. [Google Scholar] [CrossRef]
- Wang, L.; An, X.; Yang, F.; Xin, Z.; Zhao, L.; Hu, Q. Isolation and characterisation of collagens from the skin, scale and bone of deep-sea redfish (Sebastes mentella). Food Chem. 2008, 108, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Characterisation of acid soluble collagen from skins of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 85, 81–89. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, K.H.; Shin, J.H.; Lee, J.S.; Heu, M.S.; Lee, D.H.; Kim, J.S. Antioxidant and ACE inhibiting activities of the rockfish Sebastes hubbsi skin gelatin hydrolysates produced by sequential two-step enzymatic hydrolysis. Fish Aquatic Sci. 2011, 14, 1–10. [Google Scholar] [CrossRef]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure-activity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, J.; Sionkowska, A.; Skopinska-Wisniewska, J.; Piechowicz, K. Northern pike (Esox lucius) collagen: Extraction, characterization and potential application. Int. J. Biol. Macromol. 2015, 81, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Chi, C.-F.; Wang, B.; Ding, G.-F.; Li, Z.-R. Characterization of acid-and pepsin-soluble collagens from spines and skulls of skipjack tuna (Katsuwonus pelamis). Chin. J. Nat. Med. 2014, 12, 712–720. [Google Scholar] [CrossRef]
- Li, Z.R.; Wang, B.; Chi, C.F.; Zhang, Q.H.; Gong, Y.D.; Tang, J.J.; Luo, H.Y.; Ding, G.F. Isolation and characterization of acid soluble collagens and pepsin soluble collagens from the skin and bone of Spanish mackerel (Scomberomorous niphonius). Food Hydrocoll. 2013, 31, 103–113. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides-opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, L.; Piot, J.M.; Guillochon, D.; Amiot, J. Study of the efficiency of a mobile phase used in size-exclusion HPLC for the separation of peptides from a casein hydrolysate according to their hydrodynamic volume. Chromatographia 1991, 32, 499–504. [Google Scholar] [CrossRef]
- Mahmoud, M.I.; Malone, W.T.; Cordle, C.T. Enzymatic hydrolysis of casein: Effect of degree of hydrolysis on antigenicity and physical properties. J. Food Sci. 1992, 57, 1223–1229. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldrá, F.; Aristoy, M.-C.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods 2015, 13, 225–238. [Google Scholar] [CrossRef]
- Ngo, D.H.; Qian, Z.J.; Ryu, B.; Park, J.W.; Kim, S.K. In vitro antioxidant activity of a peptide isolated from Nile tilapia (Oreochromis niloticus) scale gelatin in free radical-mediated oxidative systems. J. Funct. Foods 2010, 2, 107–117. [Google Scholar] [CrossRef]
- Cai, L.; Wu, X.; Zhang, Y.; Li, X.; Ma, S.; Li, J. Purification and characterization of three antioxidant peptides from protein hydrolysate of grass carp (Ctenopharyngodon idella) skin. J. Funct. Foods 2015, 16, 234–242. [Google Scholar] [CrossRef]
- Leong, L.P.; Shui, G. An investigation of antioxidant capacity of fruits in Singapore markets. Food Chem. 2002, 76, 69–75. [Google Scholar] [CrossRef]
- Senevirathne, M.; Kim, S.H.; Siriwardhana, N.; Ha, J.H.; Lee, K.W.; Jeon, Y.J. Antioxidant potential of ecklonia cavaon reactive oxygen species scavenging, metal chelating, reducing power and lipid peroxidation inhibition. Food Sci. Technol. Int. 2006, 12, 27–38. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.J.; Vaughan, D.E. Angiotensin-converting enzyme inhibitors. Circulation 1998, 97, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Alemán, A.; Giménez, B.; Pérez-Santin, E.; Gómez-Guillén, M.C.; Montero, P. Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activities of peptide sequences isolated from squid gelatin hydrolysate. Food Chem. 2011, 125, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Iroyukifujita, H.; Eiichiyokoyama, K.; Yoshikawa, M. Classification and antihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar] [CrossRef]
- Ikoma, T.; Kobayashi, H.; Tanaka, J.; Walsh, D.; Mann, S. Physical properties of type I collagen extracted from fish scales of Pagrus major and Oreochromis niloticas. Int. J. Biol. Macromol. 2003, 32, 199–204. [Google Scholar] [CrossRef]
- Stepnowski, P.; Ólafsson, G.; Helgason, H.; Jastorff, B. Preliminary study on chemical and physical principles of astaxanthin sorption to fish scales towards applicability in fisheries waste management. Aquaculture 2004, 232, 293–303. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- AOAC. Methods of Analysis, 14th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1984. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. An overview of peanut allergenicity and enzymatic treatment for reducing it. Med. Res. Arch. 2018, 6, 5. [Google Scholar]
- Pal, G.K.; Suresh, P. Comparative assessment of physico-chemical characteristics and fibril formation capacity of thermostable carp scales collagen. Mater. Sci. Eng. C 2017, 70, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.N.; Chen, P.W.; Huang, C.Y. Compositional characteristics and in vitro valuations of antioxidant and neuroprotective properties of crude extracts of fucoidan prepared from compressional puffing-pretreated Sargassum crassifolium. Mar. Drugs 2017, 15, 183. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Wu, T.C.; Hsieh, S.L.; Tsai, Y.H.; Yeh, C.W.; Huang, C.Y. Antioxidant activity and growth inhibition of human colon cancer cells by crude and purified fucoidan preparations extracted from Sargassum cristaefolium. J. Food Drug. Anal. 2015, 23, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Wu, S.J.; Yang, W.N.; Kuan, A.W.; Chen, C.Y. Antioxidant activities of crude extracts of fucoidan extracted from Sargassum glaucescens by a compressional-puffing-hydrothermal extraction process. Food Chem. 2016, 197, 1121–1129. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Analysis (%) | C. chanos |
|---|---|
| Moisture | 9.82 ± 0.42 1 |
| Crude protein | 53.6 ± 0.7 |
| Crude lipid | 0.25 ± 0.04 |
| Ash | 33.0 ± 0.2 |
| Carbohydrate | 3.34 ± 0.18 |
| Variables of Extrusion | FSG1 | FSG2 |
| Preconditioning solvent | — | ddH2O |
| Moisture content of fish scale powder (%) | — | 27 |
| Feed supply (kg/h) | — | 11.4 |
| Die diameter (mm) | — | 5 |
| Screw speed (rpm) | — | 360 |
| Barrel temperature (°C) | — | 135 |
| Variables of Water Extraction | FSG1 | FSG2 |
| Extraction temperature (°C) | 50 | 50 |
| Extraction time (h) | 1 | 1 |
| Extraction Yield of Gelatins | FSG12 | FSG22 |
| Extraction yield (%) 1 | 4.03 ± 0.05 | 11.4 ± 0.2 *** |
| Process Variables | FSGH1 | FSGH2 | FSGH3 | FSGH4 |
| Gelatin source | FSG1 | FSG1 | FSG2 | FSG2 |
| Enzyme used | Flavourzyme | Alcalase + Flavourzyme | Flavourzyme | Alcalase + Flavourzyme |
| Digestion conditions | pH 7.5, 50 °C, 2 h | pH 7.5, 50 °C, 2 h | pH 7.5, 50 °C, 2 h | pH 7.5, 50 °C, 2 h |
| Ultrafiltration condition | <3 kDa | <3 kDa | <3 kDa | <3 kDa |
| Amino Acid Composition | FSGH1 | FSGH2 | FSGH3 | FSGH4 |
| Aspartic acid/asparagine | 58.8 ± 1.5 c | 50.9 ± 5.0 b | 37.1 ± 0.9 a | 51.8 ± 1.7 b |
| Glutamic acid/glutamine | 89.9 ± 2.0 d | 67.7 ± 2.6 c | 28.6 ± 0.7 a | 39.8 ± 1.3 b |
| Serine | 23.5 ± 1.6 ab | 28.3 ± 12.0 b | 10.0 ± 0.2 a | 13.9 ± 0.5 ab |
| Histidine | 18.2 ± 1.6 a | 18.3 ± 1.0 a | 27.9 ± 3.0 b | 19.1 ± 3.5 a |
| Glycine | 311 ± 1 ab | 292 ± 5 ab | 326 ± 21 b | 288 ± 21 a |
| Threonine | 22.5 ± 0.3 b | 27.7 ± 3.6 c | 9.25 ± 0.22 a | 12.9 ± 0.4 a |
| Arginine | 46.0 ± 5.0 a | 46.4 ± 2.6 a | 49.8 ± 3.3 a | 52.2 ± 3.6 a |
| Alanine | 134 ± 2 a | 140 ± 3 a | 151 ± 8 a | 143 ± 14 a |
| Tyrosine | 7.93 ± 0.24 a | 10.2 ± 1.9 a | 9.86 ± 0.25 a | 9.20 ± 0.26 a |
| Cystine | 2.04 ± 0.32 a | 6.71 ± 0.33 c | 3.83 ± 0.44 b | 6.16 ± 0.89 c |
| Valine | 23.2 ± 0.1 a | 37.5 ± 1.6 c | 30.5 ± 1.6 b | 39.4 ± 2.2 c |
| Methionine | 10.8 ± 1.2 a | 12.6 ± 0.6 a | 16.8 ± 2.5 b | 13.6 ± 2.0 ab |
| Phenylalanine | 16.5 ± 0.1 a | 32.1 ± 1.3 c | 19.9 ± 1.4 b | 32.3 ± 2.3 c |
| Isoleucine | 10.6 ± 0.2 a | 18.6 ± 1.3 b | 12.1 ± 0.9 a | 17.9 ± 1.5 b |
| Leucine | 21.6 ± 0.3 a | 39.2 ± 1.1 c | 29.0 ± 1.5 b | 42.3 ± 3.0 c |
| Lysine | 15.9 ± 0.2 ab | 13.5 ± 1.3 a | 18.5 ± 2.7 b | 15.8 ± 0.9 ab |
| Tryptophan | 6.26 ± 0.59 a | 9.74 ± 1.99 bc | 7.33 ± 0.79 ab | 10.7 ± 0.4 c |
| Proline | 126 ± 2 b | 102 ± 3 a | 146 ± 10 c | 125 ± 5 b |
| Hydroxylysine | 12.5 ± 0.4 a | 18.7 ± 1.0 c | 16.6 ± 0.5 b | 22.1 ± 0.5 d |
| Hydroxyproline | 42.0 ± 0.7 b | 26.7 ± 1.4 a | 49.9 ± 4.2 c | 44.9 ± 1.5 bc |
| Total | 1000 | 1000 | 1000 | 1000 |
| Hydrolysate | Protein Origin | Molecular Mass (+1) | Peptide Sequence | Modification |
|---|---|---|---|---|
| FSGH1 | Collagen 1a1-like | 1354.68 | M.GPRGPPGPPGPSGPQ.G | |
| 1411.70 | M.GPRGPPGPPGPSGPQG.F | |||
| Collagen alpha-5(VI) chain (Fragment) | 1443.69 | V.GPRGSPGPPGQPGPQG.F | 1 Deamidated (NQ) | |
| FSGH2 | collagen alpha-1(I) chain-like | 1072.55 | M.GPRGPPGPPGPS.G | |
| 1354.68 | M.GPRGPPGPPGPSGPQ.G | |||
| 1401.73 | P.AGPSGPRGPAGPAGPR.G | |||
| 1411.70 | M.GPRGPPGPPGPSGPQG.F | |||
| collagen alpha-1(I) chain | 1370.71 | A.GPRGLPGPPGSPGPQ.G | ||
| Collagen alpha-1(I) chain (Fragments) | 1370.71 | S.GPRGIPGPPGSPGPQ.G | ||
| collagen alpha-1(I) chain-like | 1427.73 | M.GPRGLPGPPGPSGPQG.F | ||
| Collagen alpha-1(IV) chain (Fragment) | 1443.69 | L.GPRGSPGPPGQPGPQG.P | 1 Deamidated (NQ) | |
| Collagen alpha-4(IV) chain | 1087.64 | I.GPLGPLGPIGIP.G | ||
| Collagen alpha-1(III) chain (Fragment) | 1329.64 | P.GNPGSPGPPGPLGPQ.G | 1 Deamidated (NQ) | |
| Collagen alpha-1(V) chain | 1386.67 | T.GPPGRSGPQGPPGPAG.E | 1 Deamidated (NQ) | |
| Collagen | 1174.54 | S.DGAPGGPGAPGPAGP.Q | ||
| Collagen type IX alpha 1 chain | 985.51 | D.PGRGPPGPPGP.P | ||
| Putative fibril-forming collagen alpha chain-like protein (Fragment) | 976.50 | R.DGLPGPPGPIG.I | ||
| Putative Collagen iv alpha 1 chain (Fragment) | 1245.58 | L.NGAPGAPGGAPGPAGP.K | 1 Deamidated (NQ) | |
| Collagen-like surface protein | 1129.69 | V.APVIPVAPVAPV.S | ||
| Collagen alpha-1(XXI) chain (Fragment) | 974.56 | E.GPPGIPGPIGL.P | ||
| Collagen alpha-1(XXVI) chain | 1362.27 | P.GPSGQNGRPGIPGAP.G | 1 Deamidated (NQ) | |
| Collagen alpha-1, IV, chain precursor, putative | 958.53 | E.PGIPGIGPPGP.I | ||
| Collagen type IV alpha 6 chain | 974.56 | V.PGIGLPGPLGP.R | ||
| collagen alpha-5(IV) chain isoform X1 | 974.56 | M.GPPGLPGPLGI.P | ||
| Collagen alpha-2(XI) chain | 974.56 | E.GPPGPIGPIGI.P | ||
| Collagen alpha-1(XXVIII) chain (Fragment) | 974.56 | F.GPPGLPGPIGL.P | ||
| FSGH3 | collagen alpha-1(I) chain-like | 1354.68 | M.GPRGPPGPPGPSGPQ.G | |
| 1411.70 | M.GPRGPPGPPGPSGPQG.F | |||
| collagen alpha-1(VII) chain | 994.50 | A.GPAGAAGSPGPR.G | ||
| collagen alpha-1(XII) chain | 1443.69 | I.GSPGPRGQPGPPGPQG.E | 1 Deamidated (NQ) | |
| Collagen alpha-1(V) chain | 1386.67 | T.GPPGRSGPQGPPGPAG.E | 1 Deamidated (NQ) | |
| Collagen | 1370.67 | P.GPRGPPGPPGEAGQP.G | ||
| 1427.70 | P.GPRGPPGPPGEAGQPG.P | |||
| Collagen, type I, alpha 1a | 1027.47 | G.MPIPGPMGPM.G | ||
| Type 1 collagen alpha 1 | 1339.67 | T.GFPGSAGRVGPPGPS.G | ||
| Collagen triple helix repeat-containing protein | 1174.61 | G.IQGVPGPQGPAGP.Q | ||
| FSGH4 | collagen alpha-1(I) chain-like | 1044.50 | R.GPPGPPGPSGPQ.G | |
| 1101.53 | R.GPPGPPGPSGPQG.F | |||
| 1354.68 | M.GPRGPPGPPGPSGPQ.G | |||
| 1411.70 | M.GPRGPPGPPGPSGPQG.F | |||
| Collagen alpha-4(IV) chain | 985.51 | A.GPPGRPGPPGP.A | ||
| Collagen alpha-1(IV) chain (Fragment) | 1258.61 | L.GPRGSPGPPGQPGP.Q | 1 Deamidated (NQ) | |
| 1443.69 | L.GPRGSPGPPGQPGPQG.P | 1 Deamidated (NQ) | ||
| Putative Collagen iv alpha 1 chain (Fragment) | 1245.58 | L.NGAPGAPGGAPGPAGP.K | 1 Deamidated (NQ) | |
| Collagen, type XXVII, alpha 1 (Fragment) | 1258.61 | H.GPRGSPGPQGPPGP.P | 1 Deamidated (NQ) | |
| Collagen type VII alpha 1 chain | 976.50 | P.LGDPGPPGPLG.P | ||
| Collagen type XVIII alpha 1 chain | 1101.53 | P.GPPGPPGPSGPAN.T | ||
| Collagen | 963.44 | G.GPGEPGPAGAPG.T |
| IC50 Value 1 (μg/mL) | |
|---|---|
| FSGH1 | 762 ± 8 e |
| FSGH2 | 472 ± 12 b |
| FSGH3 | 592 ± 0 d |
| FSGH4 | 547 ± 1 c |
| captopril | 0.002 ± 0.000 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-Y.; Tsai, Y.-H.; Hong, Y.-H.; Hsieh, S.-L.; Huang, R.-H. Characterization and Antioxidant and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Gelatin Hydrolysates Prepared from Extrusion-Pretreated Milkfish (Chanos chanos) Scale. Mar. Drugs 2018, 16, 346. https://doi.org/10.3390/md16100346
Huang C-Y, Tsai Y-H, Hong Y-H, Hsieh S-L, Huang R-H. Characterization and Antioxidant and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Gelatin Hydrolysates Prepared from Extrusion-Pretreated Milkfish (Chanos chanos) Scale. Marine Drugs. 2018; 16(10):346. https://doi.org/10.3390/md16100346
Chicago/Turabian StyleHuang, Chun-Yung, Yung-Hsiang Tsai, Yong-Han Hong, Shu-Ling Hsieh, and Ren-Han Huang. 2018. "Characterization and Antioxidant and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Gelatin Hydrolysates Prepared from Extrusion-Pretreated Milkfish (Chanos chanos) Scale" Marine Drugs 16, no. 10: 346. https://doi.org/10.3390/md16100346
APA StyleHuang, C. -Y., Tsai, Y. -H., Hong, Y. -H., Hsieh, S. -L., & Huang, R. -H. (2018). Characterization and Antioxidant and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Gelatin Hydrolysates Prepared from Extrusion-Pretreated Milkfish (Chanos chanos) Scale. Marine Drugs, 16(10), 346. https://doi.org/10.3390/md16100346