New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism

 ,
, 

Abstract
:1. Introduction
2. Results
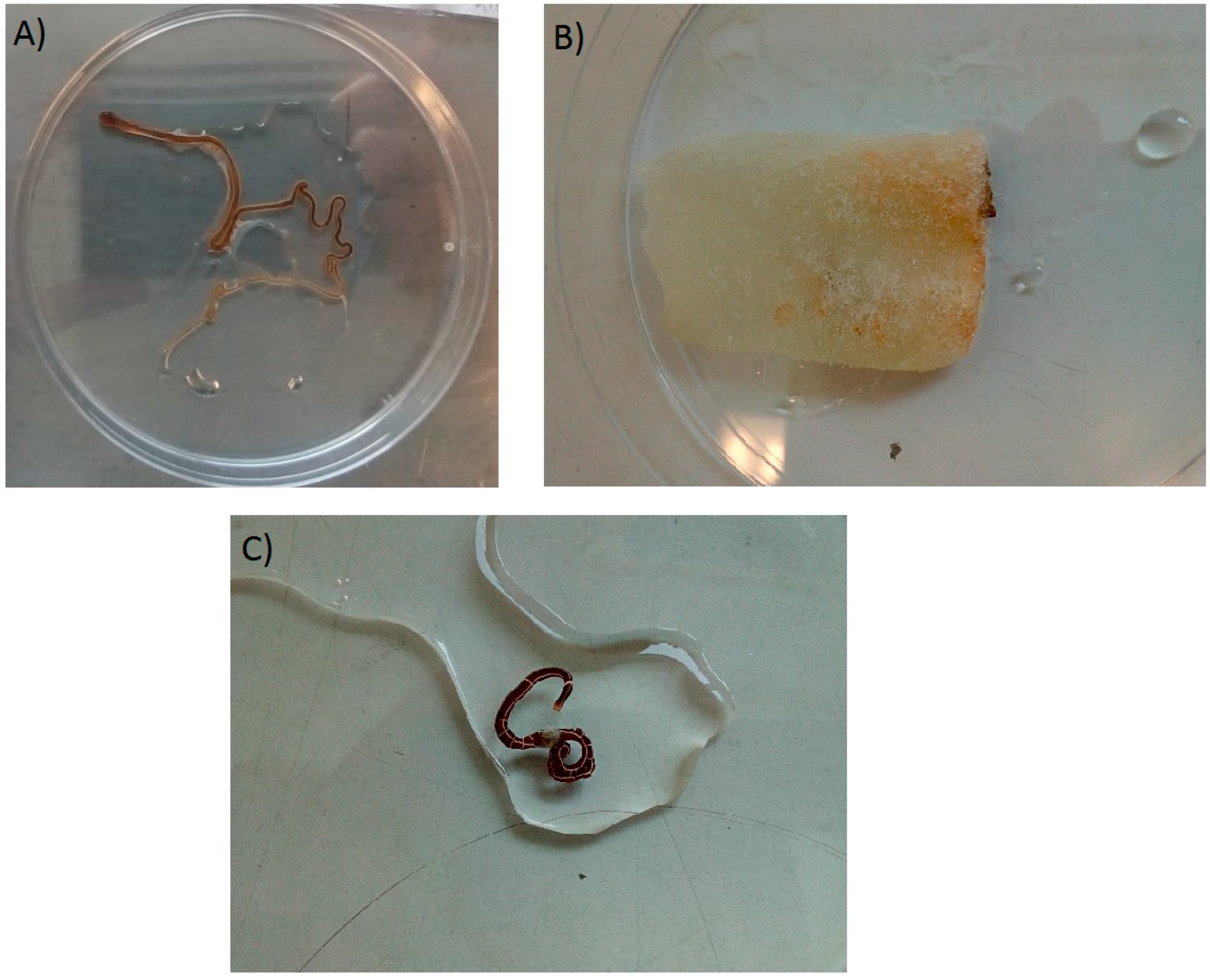
2.1. Samples
2.2. Species Identification
2.3. Microbiology
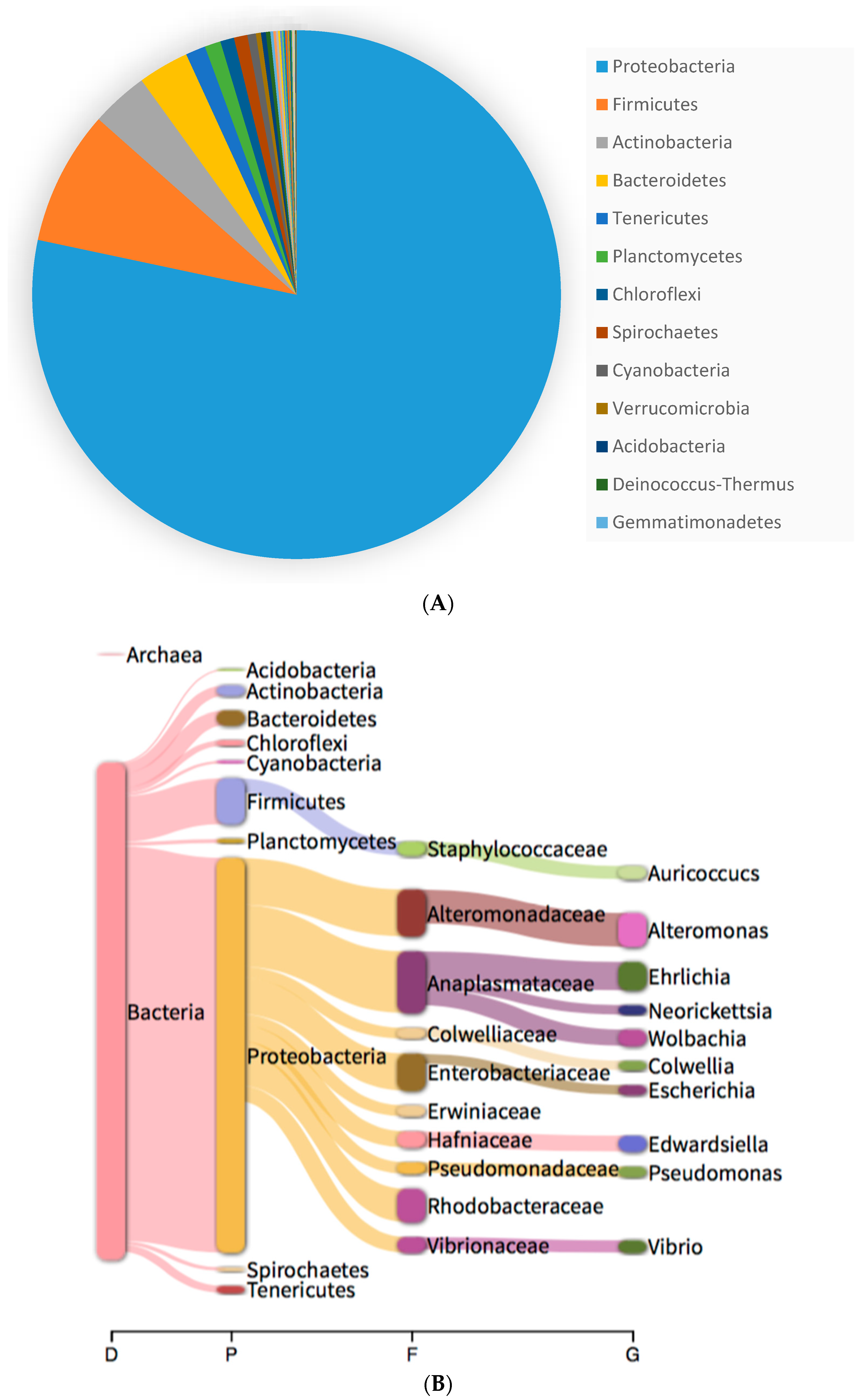
2.4. Microbiome
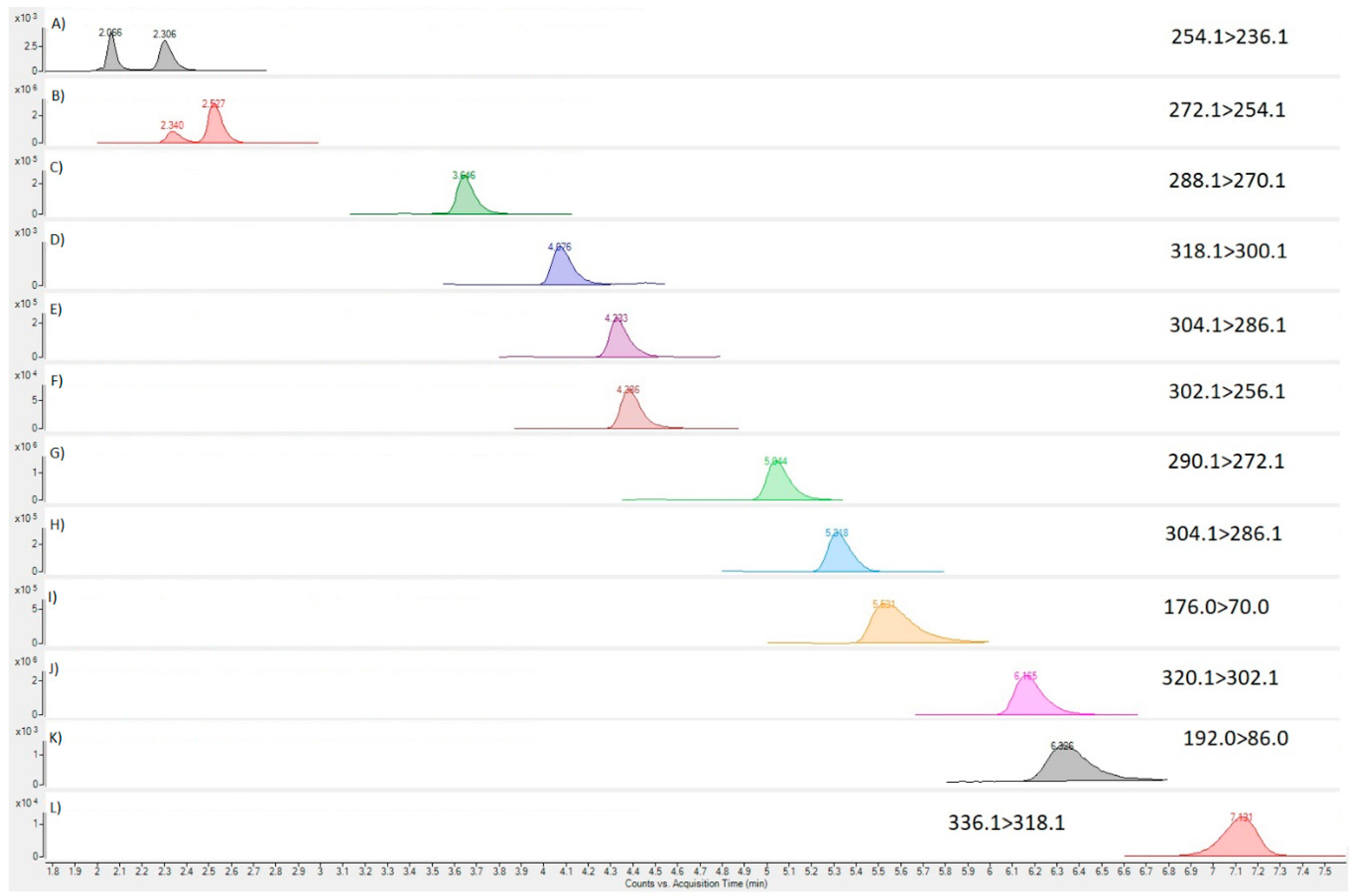
2.5. Toxin Analysis of Tissue
2.6. Toxin Analysis of Bacterial Cultures
3. Discussion
3.1. Invasive Species Implications
3.2. Microbiome
3.3. Toxicity and Toxin Profile
3.4. Human Safety
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Species Confirmation of Nemertean
4.3. Microbiological Culturing and Testing
4.4. Microbiome Sequencing
4.5. Toxin Testing
4.5.1. Nemertean Tissues
4.5.2. Bacterial Cultures
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bourque, D.; Miron, G.; Landry, T. Predation on soft-shell clams (Mya arenaria) by the nemertean Cerebratulus lacteus in Atlantic Canada: Implications for control measures. Hydrobiologia 2001, 456, 33–44. [Google Scholar] [CrossRef]
- Bourque, G.; Miron, G.; Landry, T. Predator-prey relationship between the nemertean Cerebratulus lacteus and the soft-shell clam, Mya arenaria: Surface-exploration activity and qualitative observations on feeding behaviour. Can. J. Zool. 2002, 80, 1204–1211. [Google Scholar] [CrossRef]
- Gibson, R. Nemerteans; Hutchinson: London, UK, 1972; p. 224. [Google Scholar]
- Brusca, R.; Brusca, G. Invertebrates, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Roe, P.; Norenburg, J.; Maslakova, S. Nemertea. In The Light and Smith Manual: Intertidal Invertebrates from Central Californica to Oregon, 4th ed.; Carlton, J., Ed.; University of California Press: Berkeley, CA, USA, 2007; pp. 182–196. [Google Scholar]
- Norenburg, J.; Gibson, R. Nemertea. World Register of Marine Species. 2013. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=152391 (accessed on 17 July 2018).
- Bacq, Z.M. Les poisons des Nemertiens. Bull. Cl. Sci. Acad. R. Belg. 1936, 22, 1072–1079. [Google Scholar]
- Bacq, Z.M. L’amphiporine’ et la ‘némertine’, poisons des vers némertiens. Arch. Int. Physiol. 1937, 44, 190–204. [Google Scholar]
- Jennings, J.B.; Gibson, R. Observations on the nutrition of seven species of rhynchocoelan worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- McDermott, J.J. The feeding biology of Nipponnemerte pulcher (Johnston) (Hoplonemertea), with some ecological implications. Ophelia 1984, 23, 1–21. [Google Scholar] [CrossRef]
- Ruppert, E.; Fox, R.; Barnes, R. Invertebrate Zoology: A Functional Evolutionary Approach, 7th ed.; Thomson-Brooks/Cole: Belmont, CA, USA, 2004. [Google Scholar]
- Wang, H.-Y.; Sun, S.-C.; Li, Q.-L. Laboratory observations on the feeding behaviour and feeding rate of the nemertean Procephalothrix simulus. Biol. Bull. 2008, 214, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Carefoot, T. Defenses of Ribbonworms. A Snail’s Odyssey. 2010. Available online: http://www.asnailsodyssey.com/LEARNABOUT/FLATWORM/flatPred.php accessed on-line (accessed on 17 July 2018).
- Asakawa, M.; Ito, K.; Kajihara, H. Highly toxic ribbon worm Cephalothrix simula containing tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Higashiyama, M.; Hori, K.; Noguchi, T.; Ito, K.; Hashimoto, K. Distribution of tetrodotoxin in various organs of the starfish Astopecten polyacanthus. Mar. Biol. 1987, 96, 385–390. [Google Scholar] [CrossRef]
- Isbister, G.K.; Kiernan, M.C. Neurotoxic marine poisoning. Lancet Neurol. 2005, 4, 219–228. [Google Scholar] [CrossRef]
- Yasumoto, T.; Hashimoto, Y.; Bagnis, R.; Randall, J.E.; Banner, A.H. Toxicity of the Surgeon fishes. Bull. Jpn. Soc. Sci. Fish. 1971, 37, 724–734. [Google Scholar] [CrossRef]
- Yasumoto, T.; Nakamura, M.; Oshima, Y.; Takahata, J. Construction of a continuous tetrodotoxins analyser. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 1481–1483. [Google Scholar] [CrossRef]
- Yasumoto, T.; Endo, A.; Yasumura, D.; Nagai, H.; Murata, M.; Yotsu, M. Bacterial production of Tetrodotoxin and its derivatives. Toxicon 1988, 26, 50. [Google Scholar]
- Yasumoto, T. Fish poisoning due to toxins of microalgal origins in the Pacific. Toxicon 1998, 36, 1515–1518. [Google Scholar] [CrossRef]
- Mebs, D.; Yotsu-Yamashita, M.; Yasumoto, T.; Lotters, S.; Schluter, A. Further report of the occurrence of tetrodotoxin in Atelopus species (Family: Bufonidae). Toxicon 1995, 33, 246–249. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D. Occurrence of 11-oxotetrodotoxin in the red-spotted newt, Notophthalmus viridescens, and further studies on the levels of tetrodotoxin and its analogues in the newt’s efts. Toxicon 2003, 41, 893–897. [Google Scholar] [CrossRef]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Norcardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Maran, B.A.V.; Honda, S.; Ohtsuka, S.; Arakawa, O.; Takatani, T.; Asakawa, M.; Boxshall, G.A. Accumulation of tetrodotoxin (TTX) in Pseudocaligus fugu, a parasitic copepod from panther puffer Takifugu pardalis, but without vertical transmission—Using an immunoenzymatic technique. Toxicon 2006, 48, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Yotsu-Yamashita, M. Distribution of Tetrodotoxin, saxitoxin and their analogs among the tissues of the puffer fish Fugu pardalis. Toxicon. 2006, 48, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Jen, H.-C.; Lin, S.-J.; Lin, S.-Y.; Huang, Y.W.; Liao, I.C.; Arakawa, O.; Hwang, D.F. Occurrence of tetrodotoxin and paralytic shellfish poisons in a gastropod implicated in food poisoning in Southern Taiwan. Food Addit. Contam. 2007, 24, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Jen, H.-C.; Nguyen, T.A.-T.; Wu, Y.-J.; Hoang, T.; Arakawa, O.; Lin, W.-F.; Hwang, D.-F. Tetrodotoxin and paralytic shellfish poisons in gastropod species from Vietnam analysed by high performance liquid chromatography-tandem mass spectrometry. J. Food Drug Anal. 2014, 22, 178–188. [Google Scholar] [CrossRef]
- Hwang, D.F.; Noguchi, T. Tetrodotoxin poisoning. Adv. Food Nutr. Res. 2007, 52, 141–236. [Google Scholar] [PubMed]
- Hwang, P.-A.; Tsai, Y.-H.; Lin, H.-P.; Hwang, D.-F. The gastropod possessing TTX and/or PSP. Food Rev. Int. 2007, 23, 321–340. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Alonso, E.; Rubiolo, J.A.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.A.; Lekha Menon, M.; et al. The association of bacterial C9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 40880. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, O.; Hwang, D.-F.; Taniyama, S.; Takatani, T. Toxins of pufferfish that cause human intoxications. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; National Institute of Informatics: Tokyo, Japan, 2010; pp. 227–244. [Google Scholar]
- Fernandez-Ortega, J.F.; Morales-de los Santos, J.M.; Herrera-Gutierrez, M.E.; Fernandez-Sanchez, V.; Rodriguez Loureo, P.; Rancano, A.A.; Tellez-Andrade, A. Seafood intoxication by tetrodotoxin: First case in Europe. J. Emerg. Med. 2010, 39, 612–617. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, L.A.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of Tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculate, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 56, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Azavedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Higgins, C.; Higman, W.; Hungerford, J. Potential threats posed by tetrodotoxins in UK waters: Examination of detection methodology used in their control. Mar. Drugs 2015, 13, 7357–7376. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin Tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligam, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of Tetrodotoxin Shellfish Poisoning (TSP) Toxins and Causative Factors in Bivalve Molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef] [PubMed]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazacharious, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Leão, J.M.; Lozano-Leon, A.; Giraldez, J.; Vilarino, O.; Gago-Martinez, A. Preliminary results on the evaluation of the occurrence of tetrodotoxin associated to marine Vibrio spp. In bivalves from the Galician Rias (Northwest of Spain). Mar. Drugs. 2018, 16, 81. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of Tetrodotoxin in the bivalve Paphies autralis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without pre-column reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.-F.; Cheng, C.-A.; Chen, H.-C.; Jeng, S.-S.; Noguchi, T.; Ohwada, K.; Hashimoto, K. Microflora and Tetrodotoxin-producing bacteria in the lined moon shell Natica lineata. Fish. Sci. 1994, 60, 567–571. [Google Scholar] [CrossRef]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origin and biosynthesis of Tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Pratheepa, V.; Alex, A.; Silva, M.; Vasconcelos, V. Bacterial diversity and tetrodotoxin analysis in the viscera of the gastropods from Portuguese coast. Toxicon 2016, 19, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins 2017, 9, 166–186. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, M.; Marvin, H.J.P.; Kleter, G.A.; Battilani, P.; Brera, C.; Coni, E.; Cubadda, F.; Croci, L.; De Santis, B.; Dekkers, S.; et al. Climate change and food safety: An emerging issue with special focus on Europe. Food Chem. Toxicol. 2009, 47, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Tirado, M.C.; Clarke, R.; Jaykus, L.A.; McQuatters-Gollop, A.; Frank, J.M. Climate change and food safety: A review. Food Res. Int. 2010, 43, 1745–1765. [Google Scholar] [CrossRef]
- Vezzulli, L.; Colwell, R.R.; Pruzzo, C. Ocean warming and spread of pathogenic Vibrios in the aquatic environment. Microb. Ecol. 2013, 65, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Brettar, I.; Pezzati, E.; Reid, P.C.; Colwell, R.R.; Hofle, M.G.; Pruzzo, C. Long-term effects of ocean warming on the prokaryotic community: Evidence from the vibrios. ISME J. 2012, 6, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonene, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2012, 3, 73–77. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trianes, J.A.; Salmenlinna, S.; Lofdahl, M.; Siitonen, A.; Taylorm, N.G.H.; Martinez-Urtaza, J. Heat wave-associated Vibriosis, Sweden and Finland. Emerg. Infect. Dis. 2016, 22, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.; Baker-Austin, C.; Wagley, S.; Bayley, A.; Hartnell, R. Isolation of Pandemic Vibrio parahaemolyticus from UK Water and Shellfish Produce. Microb. Ecol. 2013, 65, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Botana, L. A toxicological perspective to climate change: Aquatic toxins. Chem. Res. Toxicol. 2016, 29, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Beleneva, I.A.; Chernyshev, A.V.; Kuhlevsky, A.D. Tetrodotoxin-producing Bacillus sp. From the ribbon worm (Nemertea) Cepthalothris simula (Iwata, 1952). Toxicon 2014, 85, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Do, H.K.; Kogure, K.; Simidu, U. Identification of deep-sea-sediment bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1990, 56, 1162–1163. [Google Scholar] [PubMed]
- Do, H.K.; Hamasaki, K.; Ohwada, K.; Simidu, U.; Noguchi, T.; Shida, Y.; Kogure, K. Presence of tetrodotoxin and tetrodotoxin producing bacteria in freshwater sediments. Appl. Environ. Microbiol. 1993, 59, 3934–3937. [Google Scholar] [PubMed]
- Yan, L.U.; Ruizao, Y.I. Bacillus horikoshii, a tetrodotoxin-producing bacterium isolated from the liver of puffer fish. Ann. Microbiol. 2009, 59, 453–458. [Google Scholar]
- Magarlamov, T.Y.; Shokur, O.A.; Chernyshev, A.V. Distribution of tetrodotoxin in the ribbon worm Lineus alborostratus (Takakura, 1898) (nemertea): Immunoelectron and immunofluorescence studies. Toxicon 2016, 112, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The bacterial (Vibrio alginolyticus) production of tetrodotoxin in the ribbon worm Lineus longissimus—Just a false positive? Mar. Drugs 2016, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, H.; Sun, S.-C.; Chernyshev, A.V.; Chen, H.-X.; Ito, K.; Asakawa, M.; Maslakova, S.A.; Norenburg, J.L.; Strand, M.; Sundberg, P.; et al. Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm Cephalothrix simula (Nemertea: Palaeonemertea): A species artificially introduced from the Pacific to Europe. Zool. Sci. 2013, 30, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Strand, M.; Norenburg, J.L.; Sun, S.; Kajihara, H.; Chernyshev, A.V.; Maslakova, S.A.; Sundberg, P. Statistical parsimony networks and species assemblages in cephalotrichid nemerteans (Nemertea). PLoS ONE 2010, 5, e12885. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Alvarez, F.A.; Machordom, A. DNA barcoding reveals a cryptic nemertean invasion in Atlantic and Mediterranean waters. Helgol. Mar. Res. 2013, 67, 599–605. [Google Scholar] [CrossRef] [Green Version]
- International Council for the Exploration of the Sea (ICES). Report of the ICES/IOC/IMO Working Group on Ballast and Other Ship Vectors (WGBOSV), 16–18 March 2015, Bergen, Norway; ICES CM 2015/SSGEPI:09; ICES: Copenhagen, Denmark, 2015; 102p. [Google Scholar]
- Herrera-Bachiller, A. Los Nemertinos de España y Portugal. Ph.D. Thesis, Universidad de Alcalá, Alcalá de Henares, Spain, 2016. [Google Scholar]
- Faasse, M.; Turbeville, J. The first record of the north-west Pacific nemertean Cephalothrix simula in northern Europe. Mar. Biodivers. Rec. 2015, 8, E17. [Google Scholar] [CrossRef]
- Carroll, S.; McEvoy, E.G.; Gibson, R. The production of tetrodototoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Biolg. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-laboratory validation of a multitoxin ultra-performance LC-hydrophilic interaction LC-MS/MS method for quantitation of Paralytic Shellfish Toxins in bivalve shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, P.; Strand, M. Nemerteantaxonomy—Time to change lane? J. Zool. Syst. Evol. Res. 2010, 48, 283–284. [Google Scholar]
- Sundberg, P.; Chernyshev, A.V.; Kajihara, H.; Kånneby, T.; Strand, M. Character-matrix based descriptions of two new nemertean (Nemertea) species. Zool. J. Linn. Soc. 2009, 157, 264–294. [Google Scholar] [CrossRef] [Green Version]
- Puerta, P.; Andrade, S.C.S.; Junoy, J. Redescription of Lineus acutifrons Southern, 1913 (Nemertea: Pilidiophora) and comments on its phylogenetic position. J. Nat. Hist. 2010, 44, 2363–2378. [Google Scholar] [CrossRef]
- Sundberg, P.; Turbeville, J.M.; Lindh, S. Phylogenetic relationships among higher nemertean taxa inferred from 18S rDNA sequences. Mol. Phylogenet. Evol. 2001, 20, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, P.; Gibson, R.; Strand, M. Swedish nemerteans (phylum Nemertea), with description of a new hoplonemertean genus and species. J. Nat. Hist. 2007, 41, 2287–2299. [Google Scholar] [CrossRef]
- Junoy, J.; Andrade, S.C.S.; Giribet, G. Phylogenetic placement of a new hoplonemertean species commensal on ascidians. Invertebr. Syst. 2010, 24, 616–629. [Google Scholar] [CrossRef]
- Caballer, M.; Ortea, J. New data on the genus Hermaea (Mollusca: Sacoglossa) in Europe, with the description of a new species from Spain. Rev. Acad. Canar. Sienc. 2015, 26, 125–136. [Google Scholar]
- Gainey, P.; Guiry, M.D.; Fenwick, D.; Alder, A. Anadyomene stellata (Wulfen) C. Agardh (Anadyomenaceae, Chlorophyta), a vagrant new to Britain. Not. Algarum 2016, 3, 1–4. [Google Scholar]
- Tidbury, H.J.; Taylor, N.G.; Copp, G.H.; Garnacho, E.; Stebbing, P.D. Predicting and mapping the risk of introduction of marine non-indigenous species into Great Britain and Ireland. Biol. Invasions 2016, 18, 3277–3292. [Google Scholar] [CrossRef]
- Townhill, B.; Pinnegar, J.; Tinker, J.; Jones, M.; Simpson, S.; Stebbing, P.; Dye, S. Non-native marine species in North-West Europe: Developing an approach to assess future spread using regional downscaled climate projections. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 1035–1050. [Google Scholar] [CrossRef]
- Pingree, R.D.; Le Cann, B. Structure, strength and seasonality of the slope currents in the Bay of Biscay region. J. Mar. Biol. Assoc. UK 1990, 70, 857–885. [Google Scholar] [CrossRef]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Noguchi, T.; Jeon, J.K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.C.; Yu, P.H.; Ho, K.C.; Lee, F.W. Isolation and identification of a new tetrodotoxin-producing bacterial species, Raoultella terrigena, from Hong Kong marine puffer fish Takifugu niphobles. Mar. Drugs 2011, 9, 2384–2396. [Google Scholar] [CrossRef] [PubMed]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ. Toxicol. Pharmacol. 2013, 36, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Salvitti, L.; Wood, S.A.; Taylor, D.I.; McNabb, P.; Cary, S.C. First identification of tetrodotoxin in the flatworm Stylochoplana sp.; a source of TTX for the sea slug Pleurobranchaea maculate. Toxicon 2015, 95, 23–29. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Technical Paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; FAO/WHO: Rome, Italy, 2016; 108p. [Google Scholar]
- Kawabata, T. Puffer toxin. In The Manual for the Methods of Food Sanitation Tests; Environmental Health Bureau, Japan Food Hygienic Association: Tokyo, Japan, 1978; Volume 2, p. 232. [Google Scholar]
- Noguchi, T.; Narita, H.; Maruyama, J.; Hasimoto, K. Tetrodotoxin in the starfish Astropecten polyacanthus, in association with toxification of a trumpet shell, “Boshubora” Charonia sauliae. Nippon Suisan Gakkaishi 1982, 48, 1173–1177. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Narita, H.; Noguchi, T.; Maruyama, J.; Nara, M.; Hashimoto, K. Occurrence of tetrodotoxin associated substances in a gastropod “Hanamushirogai” Zerrxis siquijorensis. Nippon Suisan Gakkaishi 1984, 50, 85–88. [Google Scholar] [CrossRef]
- Halstead, B.W. Phylum Rhychocoela. In Poisonous and Venomous Marine Animals of the World; Darwin Press: Princeton, NJ, USA, 1978; pp. 334–343. [Google Scholar]
- EFSA. Scientific opinion on the risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar] [CrossRef]
- Codex. CODEX, CODEX STAN; FAO/WHO: Rome, Italy, 2008; pp. 292–2008. [Google Scholar]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.; Agudelo, N.; Ranwez, V.; Boehm, J.; Ryuji, J.M. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Di Pinto, A.; Ciccarese, G.; Tantillo, G.; Catalano, D.; Forte, V.T. A collagenase-targeted multiplex PCR assay for identification of Vibrio alginolyticus, Vibrio cholerae, and Vibrio parahaemolyticus. J. Food Prot. 2005, 68, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Hu, C. Vibrio alginolyticus gyrB sequence analysis and gyrB-targeted PCR identification in environmental isolates. Dis. Aquat. Organ. 2008, 82, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Song, L.; Breitwieser, F.P.; Salzberg, S.L. Centrifuge: Rapid and sensitive classification of metagenomic sequences. Genome Res. 2016, 26, 1721–1729. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Boundy, M.J.; Dhanji-Rapkova, M. Development and single-laboratory validation of a liquid chromatography tandem mass spectrometry method for quantitation of Tetrodotoxin in mussels and oysters. J. AOAC Int. 2017, 100, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitized carbon solid phase extraction. J. Chrom. A 2015, 1387, 1012. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Matrix a | Date Collected | Location |
|---|---|---|---|
| Worm 1a | C. simula (in seawater) | 18 May 2018 | Godrevy Point, Cornwall |
| Worm 1b | C. simula (in ethanol) | 18 May 2018 | Godrevy Point, Cornwall |
| Worm 2 | C. rubifrons (in seawater with sponge) | 29 May 2018 | Skilly, Newlyn, Cornwall |
| Worm 3 | T. annulatus (in seawater) | 29 May 2018 | Skilly, Newlyn, Cornwall |
| Name | TID | Relative Abundance | Sample1 | Sample2 |
|---|---|---|---|---|
| Alteromonas | 226 | 5.8055 | 7.102 | 4.509 |
| Ehrlichia | 943 | 4.1325 | 5.933 | 2.332 |
| Edwardsiella | 635 | 2.94 | 3.322 | 2.558 |
| Vibrio | 662 | 2.5565 | 2.579 | 2.534 |
| Wolbachia | 953 | 2.524 | 3.383 | 1.665 |
| Colwellia | 28,228 | 2.3225 | 1.933 | 2.712 |
| Pseudomonas | 286 | 2.2095 | 2.206 | 2.213 |
| Escherichia | 561 | 2.0995 | 2.034 | 2.165 |
| Auricoccucs | 2,005,363 | 1.983 | 2.717 | 1.249 |
| Sulfitobacter | 60,136 | 1.8645 | 1.481 | 2.248 |
| TTX | 6,11-dideoxy TTX | 5,6,11-trideoxy TTX | 11-oxo TTX | 5-deoxy TTX/11-deoxy TTX | 4,9-anhydro TTX | 11-nor TTX-6-ol | 4,9-anhydro-5,6,11-trideoxy TTX | 4,9-anhydro-11-oxo TTX | |
|---|---|---|---|---|---|---|---|---|---|
| Retention time (min) | 6.17 | 3.65 | 2.53 | 7.13 | 4.33/5.32 | 4.39 | 5.04 | 2.31 | 4.68 |
| ng/mL | 580 | 186 | 86 | 41 | 8.4 | 3.2 | 0.57 | 0.47 | 0.21 |
| ng/g | 34,791 | 11,170 | 5146 | 2454 | 504 | 193 | 35 | 28 | 12 |
| Proportion | 64% | 21% | 9% | 5% | 0.9% | 0.4% | 0.1% | 0.1% | 0.02% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Mar. Drugs 2018, 16, 452. https://doi.org/10.3390/md16110452
Turner AD, Fenwick D, Powell A, Dhanji-Rapkova M, Ford C, Hatfield RG, Santos A, Martinez-Urtaza J, Bean TP, Baker-Austin C, et al. New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Marine Drugs. 2018; 16(11):452. https://doi.org/10.3390/md16110452
Chicago/Turabian StyleTurner, Andrew D., David Fenwick, Andy Powell, Monika Dhanji-Rapkova, Charlotte Ford, Robert G. Hatfield, Andres Santos, Jaime Martinez-Urtaza, Tim P. Bean, Craig Baker-Austin, and et al. 2018. "New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism" Marine Drugs 16, no. 11: 452. https://doi.org/10.3390/md16110452
APA StyleTurner, A. D., Fenwick, D., Powell, A., Dhanji-Rapkova, M., Ford, C., Hatfield, R. G., Santos, A., Martinez-Urtaza, J., Bean, T. P., Baker-Austin, C., & Stebbing, P. (2018). New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Marine Drugs, 16(11), 452. https://doi.org/10.3390/md16110452