Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical composition
2.2. Analysis of the Elements and Organic Pollutants
2.3. Molecular Weight
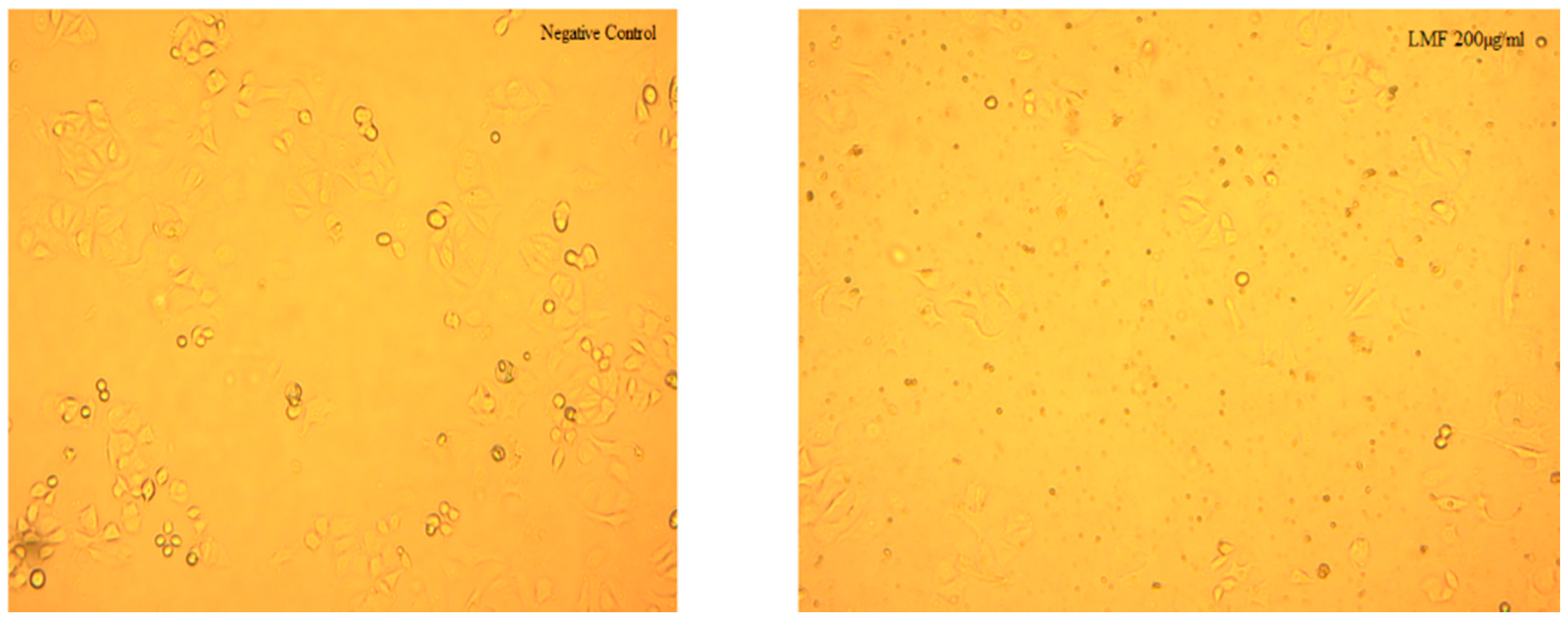
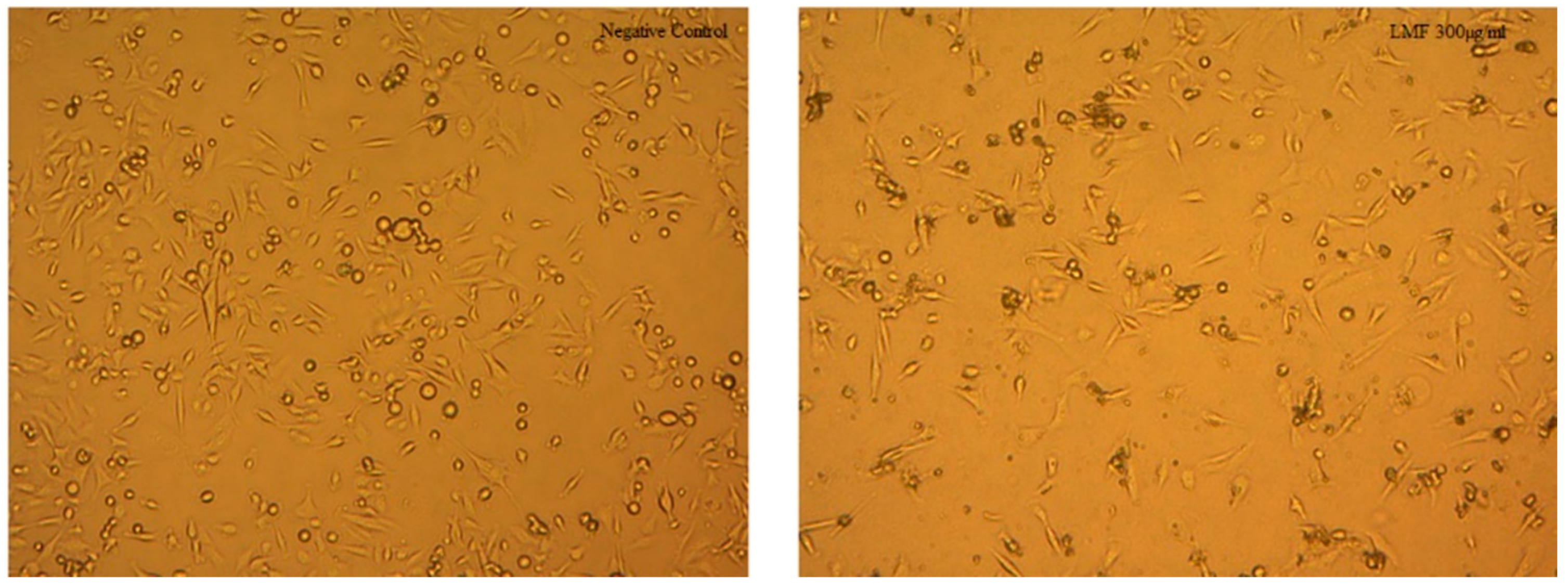
2.4. Molecular Weight Related Bioactivity of Fucoidans
2.5. Cytotoxicity Mechanisms
2.5.1. Examination of the LMWF-Induced Apoptosis
2.5.2. Examination of the Nitrosative Stress Parameters in the LMWF-Treated MDA-MB-231 Cells
2.6. Discussion
3. Materials and Methods
3.1. Materials
3.2. Extraction of the Crude Fucoidan from Local Pilot-Scale Production
3.3. Determination of the Chemical Composition
3.4. Determination of the Monosaccharide Composition
3.5. Molecular Weight Distribution
3.6. Elemental and Organic Pollutants Analysis
3.7. Preparation of Fucoidan Fractions and Stock Solution
3.8. Cell Lines
3.9. Cell Proliferation Assay
3.10. Cell Cycle Assay
3.11. Cell Mechanism Assays
3.11.1. Cell Samples Preparation
3.11.2. Cell Viability Assay
3.11.3. Cell Apoptosis Assay
Annexin V and Dead Cell Assay
Multi-Caspase Assay
MitoPotential Assay
3.11.4. Nitrosative Stress Measurement
3.12. Statistical Analysis
4. Conclusions
5. Future Research
Author Contributions
Funding
Conflicts of Interest
References
- Liu, M.; Liu, Y.; Cao, M.J.; Liu, G.M.; Chen, Q.; Sun, L.; & Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Kloareg, B. Isolation and analysis of cell walls of the brown marine algae Pelvetia canaliculata and Ascophyllum nodosum. Physiologie Végétale 1984, 22, 47–56. [Google Scholar]
- Skriptsova, A.V.; Shevchenko, N.M.; Zvyagintseva, T.N.; Imbs, T.I. Monthly changes in the content and monosaccharide composition of fucoidan from Undaria pinnatifida (Laminariales, Phaeophyta). J. Appl. Phycol. 2010, 22, 79–86. [Google Scholar] [CrossRef]
- Xu, X.; Xue, C.; Chang, Y.; Wang, J.; Jiang, K. Chain conformational and physicochemical properties of fucoidans from sea cucumber. Carbohydr. Polym. 2016, 152, 433–440. [Google Scholar] [CrossRef] [PubMed]
- White, W.L.; White, L.N. World seaweed utilization A2—Tiwari. In Brijesh K. In Seaweed Sustainability; Troy, D.J., Ed.; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- MAF Biosecurity. The commercial use of Undaria pinnatifida—An exotic Asian seaweed. MAF Biosecurity New Zealand Information: Wellington, New Zealand, 2010. [Google Scholar]
- White, L.N.; Lu, J.; White, W.L. Scoping Assessment of the Economic Viability of Harvesting Undaria Pinnatifida from NZ Mussel Lines and Potential Uses of the Collected Material; Report Number 14/01; Applied Ecology New Zealand: Auckland, New Zealand, May 2014. [Google Scholar]
- Mak, W.; Hamid, N.; Liu, T.; Lu, J.; White, W. Fucoidan from New Zealand Undaria pinnatifida: Monthly variations and determination of antioxidant activities. Carbohydr. Polym. 2013, 95, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, Q.; Wang, J.; Zhang, W. A comparative study of the anticoagulant activities of eleven fucoidans. Carbohydr. Polym. 2013, 91, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, S.; Menon, T.; Hanna, L.E.; Suresh, V.; Sathuvan, M.; Manikannan, M. In vitro anti-HIV-1 activity of fucoidan from Sargassum swartzii. Int. J. Biol. Macromol. 2016, 82, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Vishchuk, O.S.; Sun, H.; Wang, Z.; Ermakova, S.P.; Xiao, J.; Lu, T.; Xue, P.; Zvyagintseva, T.N.; Xiong, H.; Shao, C. PDZ-binding kinase/T-LAK cell-originated protein kinase is a target of the fucoidan from brown alga Fucus evanescens in the prevention of EGF-induced neoplastic cell transformation and colon cancer growth. Oncotarget 2016, 7, 18763. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-J.; Lee, B.-Y. Fucoidan from the sporophyll of Undaria pinnatifida suppresses adipocyte differentiation by inhibition of inflammation-related cytokines in 3T3-L1 cells. Nutr. Res. 2012, 32, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.K.; Cheng, J.J.; Lin, C.Y.; Chang, C.C. Purification, structural elucidation, and anti-inflammatory effect of a water-soluble 1,6-branched 1,3-α-d-galactan from cultured mycelia of Poria cocos. Food Chem. 2010, 118, 349–356. [Google Scholar] [CrossRef]
- Skriptsova, A. Seasonal variations in the fucoidan content of brown algae from Peter the Great Bay, Sea of Japan. Russ. J. Mar. Biol. 2016, 42, 351–356. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Jeon, Y.J. Biological activities and potential industrial applications of fucose rich sulfated polysaccharides and fucoidans isolated from brown seaweeds: A review. Carbohydr. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Mak, W.; Wang, S.K.; Liu, T.; Hamid, N.; Li, Y.; Lu, J.; White, W.L. Anti-proliferation potential and content of fucoidan extracted from sporophyll of New Zealand Undaria pinnatifida. Front. Nutr. 2014, 1, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.L.; Lee, B.Y.; You, S.G. Relationship between oversulfation and conformation of low and high molecular weight fucoidans and evaluation of their in vitro anti-cancer activity. Molecules 2011, 16, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chung, D.; Shin, S.; Lee, H.Y.; Kim, J.C.; Lee, Y.J.; Sang, G.Y. Effects of molecular weight and hydrolysis conditions on anticancer activity of fucoidans from sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, H.; Miki, Y.; Kimura, T.; Tanaka, K.; Nakagawa, T.; Kawamukai, M.; Matsuda, H. Effects of fucoidan from mozuku on human cell lines. Food Sci. Technol. Resour. 2006, 12, 218–222. [Google Scholar] [CrossRef]
- You, S.G.; Yang, C.; Lee, H.Y.; Lee, B.Y. Molecular characteristics of partially hydrolyzed fucoidans from sporophyll of Undaria Pinnatifida and their in vitro anticancer activity. Food Chem. 2010, 119, 554–559. [Google Scholar] [CrossRef]
- Zhang, Z.; Teruya, K.; Eto, H.; Shirahata, S. Induction of apoptosis by low-molecular-weight fucoidan through calcium- and caspase dependent mitochondrial pathways in MDA-MB-231 breast cancer cells. Biosci. Biotechnol. Biochem. 2013, 77, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Lake, A.C.; Vassy, R.; Di Benedetto, M.; Lavigne, D.; Le Visage, C.; Perret, G.Y.; Letourneur, D. Low molecular weight fucoidan increases VEGF165-induced endothelial cell migration by enhancing VEGF165 binding to VEGFR-2 and NRP1. J. Biol. Chem. 2006, 281, 37844–37852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Teruya, K.; Eto, H.; Shirahata, S. Fucoidan extract induces apoptosis in MCF-7 cells via a mechanism involving the ROS-dependent JNK activation and mitochondria-mediated pathways. PLoS ONE 2011, 6, e27441. [Google Scholar] [CrossRef] [PubMed]
- Pádua, D.; Rocha, E.; Gargiulo, D.; Ramos, A. Bioactive compounds from brown seaweeds: Phloroglucinol, fucoxanthin and fucoidan as promising therapeutic agents against breast cancer. Phytochem. Lett. 2015, 14, 91–98. [Google Scholar] [CrossRef]
- Xue, M.; Ge, Y.; Zhang, J.; Liu, Y.; Wang, Q.; Hou, L.; Zheng, Z. Fucoidan inhibited 4T1 mouse breast cancer cell growth in vivo and in vitro via downregulation of Wnt/β-catenin signaling. Nutr. Cancer 2013, 65, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Thinh, P.D.; Menshova, R.V.; Ermakova, S.P.; Anastyuk, S.D.; Ly, B.M.; Zvyagintseva, T.N. Structural characteristics and anticancer activity of fucoidan from the brown alga Sargassum mcclurei. Mar. Drugs 2013, 11, 1456–1476. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sun, J.; Su, X.; Qiu, L.; Yu, Q.; Yu, Q.; Zhang, P. A review about the development of fucoidan in antitumor activity: Progress and challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Song, M.G.; Kim, Y.N.; Park, J.I.; Kwak, J.Y. The mechanism of fucoidan-induced apoptosis in leukemic cells: Involvement of ERK1/2, JNK, glutathione, and nitric oxide. Mol. Carcinog. 2010, 49, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer properties and mechanisms of fucoidan on mouse breast cancer in vitro and in vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, S.; Sokolova, R.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, Eclonia cava, Costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Kim, W.-J.; Kim, S.-M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Park, Y.I. Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; You, S. Determination of physicochemical properties of sulphated fucans from sporophyll of Undaria pinnatifida using light scattering technique. Food Chem. 2008, 111, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.Y.; Robertson, J.; Hamid, N.; Ma, Q.; Lu, J. Changes in total nitrogen and amino acid composition of New Zealand Undaria pinnatifida with growth, location and plant parts. Food Chem. 2015, 186, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Volesky, B.; Mucci, A. A review of the biochemistry of heavy metal biosorption by brown algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2005, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Lin, T.Y.; Wu, Y.C.; Tsao, S.M.; Hwang, P.A.; Shih, Y.W.; Hsu, J. Fucoidan inhibition of lung cancer in vivo and in vitro: Role of the Smurf2-dependent ubiquitin proteasome pathway in TGFbeta receptor degradation. Oncotarget 2014, 5, 7870–7885. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.; Lu, J.; Roscilli, G.; Aurisicchio, L.; Cappelitti, M.; Pavoni, E.; White, W.L.; Bedogni, B. The natural compound fucoidan from New Zealand Undaria pinnatifida synergizes with the ERBB inhibitor lapatinib enhancing melanoma growth inhibition. Oncotarget 2017, 8, 17887–17896. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.L.; Tai, C.J.; Huang, C.W.; Chang, F.R.; Wang, J.Y. Efficacy of low-molecular-weight fucoidan as a supplemental therapy in metastatic colorectal cancer patients: A double-blind randomized controlled trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.; Synytsya, A.; Capek, P.; Lee, C.W.; Choi, J.W.; Cho, S.; Kim, W.J.; Park, Y. Low molecular weight mannogalactofucans derived from Undaria pinnatifida induce apoptotic death of human prostate cancer cells in vitro and in vivo. Mar. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Choi, J. Effect of molecular weight of fucoidan on its anticancer-activity. J. Cancer Sci. Ther. 2018, 10. [Google Scholar] [CrossRef]
- Hwang, P.; Yan, M.; Lin, H.; Li, K.; Lin, Y. Toxicological evaluation of low molecular weight fucoidan in vitro and in vivo. Mar. Drugs 2016, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, Q.; Wang, Q.; He, Y.; Ren, D.; Liu, S.; Wu, L. Structural characterization and antitumor effects of fucoidans from brown algae Kjellmaniella crassifolia farmed in northern China. Int. J. Biol. Macromol. 2018, 119, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, H.; Tamauchi, H.; Iizuka, M.; Nakano, T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta. Med. 2006, 72, 1415–1417. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.F.; Li, X.J.; Zhang, H.Y. Natural products and drug discovery: can thousands of years of ancient medical knowledge lead us to new and powerful drug combinations in the fight against cancer and dementia? EMBO Reports 2009, 10, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, C.; Tyner, A. A new method for determining the status of p53 in tumor cell lines of different origin. Oncol. Res. 2003, 13, 3. [Google Scholar]
- Banafa, A.; Roshan, S.; Liu, Y.; Zhao, S.; Yang, G.; He, G.; Chen, M. Fucoidan induces apoptosis in MDA-MB-231 cells by activating caspase cascade and down-regulating XIAP. J. Pharm. Biol. Sci. 2014, 9, 59–64. [Google Scholar] [CrossRef]
- Martinez, L.; Thames, E.; Kim, J.; Chaudhuri, G.; Singh, R.; Pervin, S. Increased sensitivity of African American triple negative breast cancer cells to nitric oxide-induced mitochondria-mediated apoptosis. BMC Cancer 2016, 16, 559. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.M.; Pae, H.O.; Jang, S.I.; Kim, Y.M.; Chung, H.T. Nitric oxide as a pro-apoptotic as well as anti-apoptotic modulator. BMB Reports 2002, 35, 116–126. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.; Preobrazhenskaya, M.; DIncecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.; Berman, A.; Bilan, M.; et al. A comparative study of the antiinflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.P.; Silva, E.F.; Stern, A. Nitric oxide: A potential inducer of adhesion-related apoptosis—anoikis. Nitric Oxide 2004, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, J.F.; Leon, L.; Cortier, M.; Sassi, N.; Paul, C.; Bettaieb, A. Nitric oxide-induced resistance or sensitization to death in tumor cells. Nitric Oxide 2008, 19, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Fan Yang, F.; Lu, L. Fucoidan extracted from Undaria pinnatifida: Source for nutraceuticals/functional foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef] [PubMed]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Muir, C.P.; Adams, M.A.; Graham, C.H. Nitric oxide attenuates resistance to doxorubicin in three-dimensional aggregates of human breast carcinoma cells. Breast Cancer Res. Tr. 2006, 96, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, S.; Wang, M.; Liu, X.; Zheng, B. Fluoride in chilies from southwestern China. Ann. N.Y. Acad. Sci. 2008, 1140, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Bird, G.M.; Haas, P. On the nature of the cell wall constituents of Laminaria spp. Mannuronic acid. Biochem. J. 1931, 25, 403. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dodgson, K.; Price, R. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106. [Google Scholar] [CrossRef] [PubMed]
- Bitter, T.; Muir, H.M. A modified uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Association of Official Agricultural Chemists. AOAC Official Methods of Analysis, 18th ed.; Association of Official Agricultural Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an analytical procedure for extraction of lipids from microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Xie, J.H.; Shen, M.-Y.; Nie, S.-P.; Liu, X.; Zhang, H.; Xie, M.-Y. Analysis of monosaccharide composition of Cyclocarya paliurus polysaccharide with anion exchange chromatography. Carbohydr. Polym. 2013, 98, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, Y.; Wan, Y. Determination of the trace elements in Shengmai San decoction by ICP-AES. Guang Pu Xue Yu Guang Pu Fen Xi 2008, 28, 436–440. [Google Scholar] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar (%) | Uronic Acid (%) | Protein (%) | Sulphate (%) | Ash (%) | Crude Fat (%) | Monosaccharide Composition | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Fuc (%) | Gal (%) | Glc (%) | GlcA (%) | |||||||
| S | 96.83 ± 1.71 | 1.61 ± 0.06 | tr | 25.59 ± 0.63 | 16.61 ± 2.13 | nd | 27.44 ± 0.45 | 25.34 ± 0.70 | nd | nd |
| S1 | 88.16 ± 1.85 | 1.76 ± 0.02 | 1.28 ± 0.02 | 19.68 ± 1.60 | 32.96 ± 0.04 | 1.60 ± 0.06 | 19.50 ± 0.35 | 21.20 ± 0.67 | nd | tr |
| S2 | 89.94 ± 0.32 | 3.80 ± 0.37 | 0.25 ± 0.10 | nd | 4.93 ± 0.03 | 0.62 ± 0.09 | nd | nd | 96.71 ± 0.74 | nd |
| S3 | 44.67 ± 0.15 | 2.39 ± 0.01 | 2.09 ± 0.05 | 15.48 ± 0.50 | 25.42 ± 0.05 | 0.29 ± 0.11 | 13.83 ± 0.51 | 13.24 ± 0.45 | tr | 2.46 ± 0.10 |
| S4 | 80.00 ± 1.39 | 2.65 ± 0.03 | 2.41 ± 0.10 | 20.96 ± 0.85 | 19.49 c | 0.89 ± 0.27 | 20.35 ± 2.62 | 19.26 ± 2.79 | tr | 2.02 ± 0.11 |
| S5 | 78.75 ± 0.70 | 4.15 ± 0.12 | 3.08 ± 0.0058 | 20.60 ± 0.23 | 24.33 ± 0.23 | 2.3 c | 19.23 ± 1.26 | 21.00 ± 0.20 | 6.38 | nd |
| Fraction | Total Sugar (%) | Uronic Acid (%) | Protein (%) | Sulphate (%) | Ash (%) |
|---|---|---|---|---|---|
| F<10k | 2.76 ± 0.31 | 1.42 ± 0.03 | 7.04 ± 0.47 | 16.62 ± 1.31 | 74.39 ± 0.33 |
| F10k–300k | 11.20 ± 0.39 | 2.44 ± 0.05 | 9.57 ± 0.43 | 7.17 ± 1.96 | 53.61 ± 0.35 |
| F>300k | 53.14 ± 0.13 | 2.53 ± 0.08 | 7.53 ± 1.03 | 30.75 ± 1.83 | 19.47 ± 0.71 |
| Elements and Pollutants | Sigma Sample (S) | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|---|
| S | 102,750 | 83,700 | 4200 | 60,400 | 86,600 | 76,650 |
| K | 2700 | 53,700 | 14,800 | 61,300 | 29,900 | 26,400 |
| Ca | 1100 | 46,400 | 310 | 4240 | 13,500 | 44,100 |
| Na | 80,200 | 10,800 | 6800 | 27,230 | 24,700 | 2200 |
| Mg | 1900 | 1200 | 6800 | 4500 | 7100 | 2900 |
| Cr | tr | 1.0 | 0.28 | 0.51 | 0.71 b | 7.4 |
| Pb | tr | 4.3 | 15 | 6.9 | 0.11 b | 9.2 |
| Cd | tr | 0.50 | <0.10 | 1.1 | 1.7 b | 0.84 |
| Hg | tr | 0.076 | 0.29 | 0.11 | 0.14 b | 0.56 |
| As | tr | 1.8 | 1.3 | 7.3 | 5.5 b | 0.76 |
| Cl | - a | 27,800 | 23,800 | 33,500 | - a | 210 |
| Fluoride | - a | 226,600 | 92,000 | 59,000 | - a | 393,900 |
| PCB | - a | nd | nd | nd | - a | nd |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Shi, K.K.; Chen, S.; Wang, J.; Hassouna, A.; White, L.N.; Merien, F.; Xie, M.; Kong, Q.; Li, J.; et al. Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines. Mar. Drugs 2018, 16, 461. https://doi.org/10.3390/md16120461
Lu J, Shi KK, Chen S, Wang J, Hassouna A, White LN, Merien F, Xie M, Kong Q, Li J, et al. Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines. Marine Drugs. 2018; 16(12):461. https://doi.org/10.3390/md16120461
Chicago/Turabian StyleLu, Jun, Keyu Kally Shi, Shuping Chen, Junqiao Wang, Amira Hassouna, Loretta Nicole White, Fabrice Merien, Mingyong Xie, Qingjun Kong, Jinyao Li, and et al. 2018. "Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines" Marine Drugs 16, no. 12: 461. https://doi.org/10.3390/md16120461
APA StyleLu, J., Shi, K. K., Chen, S., Wang, J., Hassouna, A., White, L. N., Merien, F., Xie, M., Kong, Q., Li, J., Ying, T., White, W. L., & Nie, S. (2018). Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines. Marine Drugs, 16(12), 461. https://doi.org/10.3390/md16120461