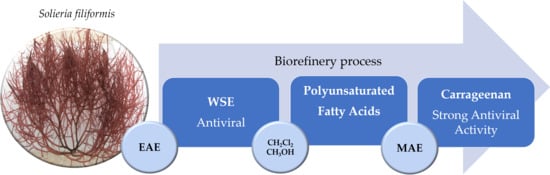
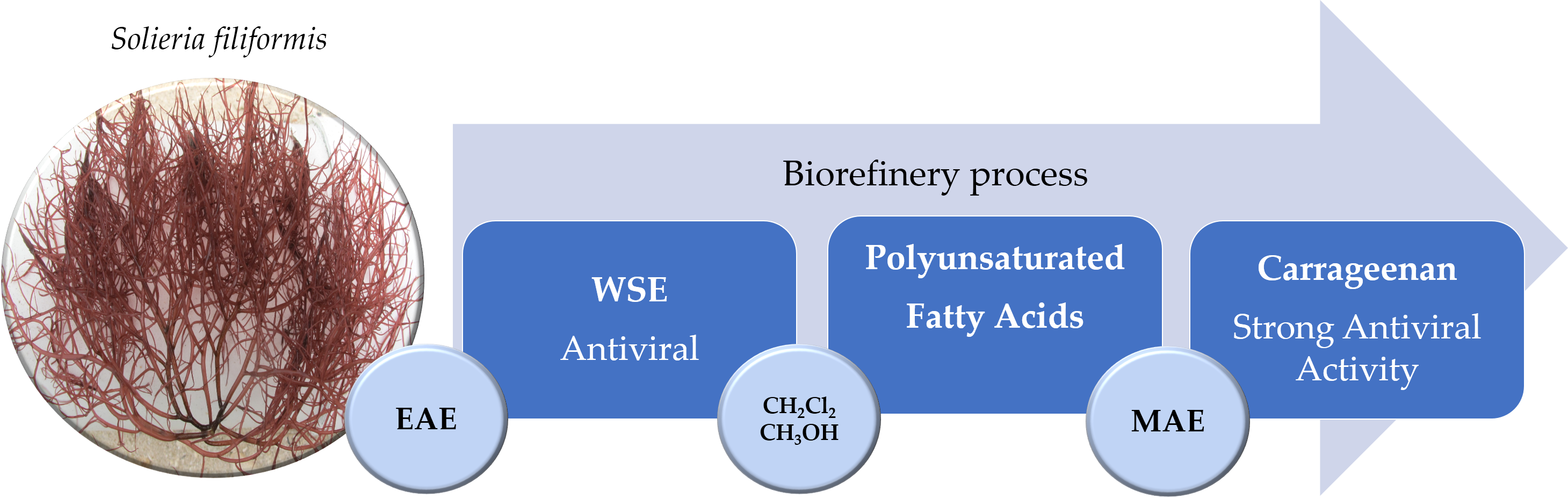
2.3.1. Recovery of Water-Soluble Extract (WSE) by Biorefinery Process ant Its Potential Application (Product 1)
In general, yields obtained using the enzyme (63.5 and 72.4%, conditions a and b respectively) increased in comparison to the blanks (without enzyme) which were 58.6% and 47.6%, showing a 24% gain for
Product 1a and 33% for
Product 1b. The yield and biochemical composition of water-soluble extract WSE obtained by EAE (
Product 1a and
Product 1b) and their corresponding residues (Residue 1
a and Residue 1
b) are presented in
Table 3. For the denatured enzyme extraction (
a), the yield was higher than that under non-denatured enzyme extraction (
b), and it was also higher than that reported for
S. chordalis (55.8%) using the same EAE conditions [
7]. A similar pattern was observed for carbohydrate and sulfate contents, with higher values obtained in
Product 1a when compared to
Product 1b (31.9 and 26.1% higher, carbohydrates and sulfates respectively). No significant differences in protein and lipid content were observed between the two conditions tested. The increase in temperature to 85 °C for 15 min to denature the enzyme allowed a higher yield of the polysaccharide. In relation to this, higher extraction efficiencies of sulfated polysaccharides, such as carrageenan, has been observed and widely reported at high temperatures [
11,
33,
34,
35]. The residues from the EAE extraction step generated under different conditions, residues 1
a and 1
b, rich in carbohydrates (~32%) and sulfate (~23%), were used to obtain
Product 2.
The cytotoxicity and in vitro antiviral activity of
Products 1a–
b are presented in
Table 4. No cytotoxic effect on the Vero cells was observed in the range of the concentrations assayed. After 3 days of treatment, no microscopically visible alteration of normal cell morphology was observed and a viability assay showed no destruction of cell layer. Only
Product 1a exhibited antiviral activity against
Herpes simplex virus. In
S. chordalis a similar EC
50 value (86.0 μg/mL) was reported using the same EAE conditions [
7]. The higher carbohydrate and sulfate contents exhibited by
Product 1a in comparison with
Product 1b (
Table 3), its probably related with a higher carrageenan proportion that could be responsible for this activity. It has been previously reported that sulfated polysaccharides, including carrageenans, have antiviral activities against enveloped viruses such as dengue virus, papillomavirus and
H. simplex [
20,
36], where the degree of sulfation is the main factor related to the antiviral activity [
37]. Many biological effects of sulfated polysaccharides are related to their ability to change the surface properties of the cell, which interferes with the early events that impede the entry of the virus since these compete with similar molecular species for initial binding to the target cell [
38].
The selectivity index (SI) was used to compare the antiviral potency of the extracts. Despite the high antiviral activity shown by
Product 1a, weak antiviral potency (<10) was obtained (
Table 4). However, it should be noted that this is a crude extract rich in sulfated polysaccharides, and this activity could be improved by adding a purification step to obtain a pure carrageenan.
2.3.2. Recovery of Fatty Acids (FAs) by Biorefinery Process and its Potential Application (Product 2)
From the EAE Residues 1
a (19.9%) and Residue 1
b (22.5%) the lipids obtained (~0.71%) were used as raw material to obtain FAs (
Product 2). The fatty acids (FAs) profile and nutritional indices (ω6/ω3 and saturated/unsaturated fatty acids (SFAs/UFAs)) of
Products 2a–
b are shown in
Table 5. The FAs profile obtained from the initial
S. filiformis raw biomass by direct extraction is also included for comparison.
The FAs profile obtained for
S. filiformis in this study was similar to those reported for other
Solieria species in previous studies [
39]. A high content of SFAs (69.2–73.6%) were obtained for both
Product 2a and
Product 2b, as well as for FAs obtained directly from raw material, without significant differences among them. Palmitic acid C16:0 was the most abundant SFA (43.9–55.8%) in all conditions. Although from a nutritional point of view dietary SFAs are usually associated with negative consequences for human health, the role of individual SFAs in metabolic disease is still incompletely understood. Individual SFAs such as myristic (C14:0) and palmitic acids, both present in
S. filiformis, are involved in protein fatty acid acetylation, which may have various regulation functions and, as has been recently reviewed, in the future dietary recommendations will probably evolve to differentiate them [
40,
41].
Concerning unsaturated FAs (UFAs), MUFAs content in
Products 2a–
b ranged from 12.47 to 16.57% and no significant differences were found between condition
a and
b. However, there was a significant reduction of MUFAs when compared with those obtained directly from raw material. Palmitoleic acid (C16:1n-7) and oleic acid (C18:1n-9c) were present in the MUFAs fraction. Healthy diets are abundant in MUFAs that are often associated with low prevalence of chronic disease. These include oleic acid that has been reported to reduce cardiovascular risk by reducing blood pressure [
42,
43], and also palmitoleic acid that has been recently reported to have anti-inflammatory and antidiabetic activities [
44].
PUFAs corresponded to ~14% in
Products 2a–
b and, contrary to MUFAs, they almost doubled their content when compared with those obtained directly from raw material. Eicosapentaenoic acid (EPA, C20:5 ω3) and arachidonic acid (AA, C20:4 ω6) were detected in
Product 2b as well as in FAs obtained directly from
S. filiformis, whereas the presence of γ-Linolenic acid and Linolenic acid (C18:3 ω6 and C18:2 ω6, respectively) were only observed in FAs obtained directly from
S. filiformis. PUFAs have a great importance for human metabolism. They are the major components of cell membrane phospholipids, and may also be present in cellular storage oils [
45,
46]. Both AA and EPA are two crucial fatty acids of marine origin. AA is the principal ω6 fatty acid in the brain development of infants, and EPA has a beneficial effect on the cardiovascular system. They are also precursors of prostaglandins, thromboxane and other eicosanoids involved in immune reactions. Because their beneficial properties such as antibacterial, anti-inflammatory, antioxidant, antitumoral properties and the prevention of cardiac diseases, they are important nutraceutical and pharmaceutical targets [
47]. The absence of AA and EPA in
Product 2a could be related to the increase of temperature (85 °C for 15 min) to denature the enzyme in that condition. Although the applications of enzymes in bioprocessing are especially advantageous because they act under mild reaction conditions, it has been reported that PUFAs are highly labile to pH or elevated temperatures that can produce an oxidation or double bond migration [
48].
As for the ω6/ω3 ratio, based on the FA results it was calculated at 0.91 for
Product 2b and 5.01 for the control. The World Health Organization (WHO) currently recommends that the ω6/ω3 ratio should not exceed 10 to have protective effects against atherosclerotic heart disease, and in the prevention of chronic diseases [
49].
In the same regard, the SFAs/UFAs ratio is also an indicator used to evaluate the quality of lipids in a nutritional sense, since UFAs are essential for the proper functioning of the human body. The balance of SFAs with UFAs, especially PUFAs, has been associated with a lower risk of heart diseases. Although Products 2a–b displayed high amounts of SFA, the high content of MUFAs and PUFAs indicates a potential nutritional value. Moreover, the increase in the relative abundance of PUFAs observed after the enzymatic extraction raises the potential value of this product once it enters into the biorefinery process.
2.3.3. Recovery of Carrageenan by Biorefinery Process and Its Potential Application (Product 3)
In the third stage of the sequential extraction process, residual biomass (Residues 2
a–
b) resulted from lipid extraction (~96%) was further processed for the recovery of
S. filiformis carrageenan by MAE. Yield and chemical characterization of carrageenan (
Products 3a–
b) are shown in
Table 6.
Carrageenan yields obtained at the end of the biorefinery process of
S. filiformis ranged between 17.1 to 29.7%, whereas that obtained by direct extraction was 25.5%, all of them were in accordance with yields reported for
Solieria chordalis by a direct extraction using MAE [
8]. Our results were also comparable, or even higher, to those reported previously by Fernandes de Araujo et al. (19.1%) [
19], and Caamal-Fuentes et al. (17.8%) [
22] for
S. filiformis using direct carrageenan extraction by conventional methods, where higher carrageenan yields should be expected because no losses related to a sequential extraction process. Moreover, in our case MAE favored the extraction of carrageenan due to the high temperature reached using closed vessels, increasing the mass transfer of carrageenan from the sample matrix. The efficiency of this method over conventional extraction methods has already been previously reported [
8,
11].
It is noteworthy that carrageenan yields and their chemical properties were significantly influenced by the extraction conditions of the first step during the sequential extraction process of
S. filiformis in relation with the denatured and non-denatured enzyme. The lowest carrageenan yield, as well as lowest sulfate content, was obtained in
Product 3a (
Table 6). During EAE extraction conditions
a and
b (corresponding to denatured and non-denaturated enzyme respectively), the high temperatures applied to denature the enzyme together with elevated temperatures reached by microwave heating could explain these differences. Clearly, cell wall maceration during enzymatic hydrolysis treatments could potentially increase the effect of MAE in the subsequent extraction processes. However, in the present study, the use of this pre-treatment before carrageenan extraction did not significantly improve the yield. This behavior can be partly explained due to a degradation of polysaccharides by an excessive thermal treatment and the release of oligosaccharides resulting in lower carrageenan yield in
Product 3a. Additionally, variability using different enzymes in
Solieria chordalis extraction treatments has been previously reported [
7,
50]. This effect, due not only to the thermal treatment itself but also to the specific aggressiveness of the enzyme, could also be related to the lower yields obtained in the present study under extraction condition
a. In this way, to better understand the effect of the different parameters involved in a sequential carrageenan extraction process, further studies on the efficacy of other enzymes to optimize the extraction capacity should be performed. In this study, sulfate and 3,6 anhydro-galactose content were in the range to those previously reported in
S. filiformis [
20,
22]. The Fourier transform–infrared (FT–IR) spectra are shown in
Figure 1.
The presence of ι-carrageenan in
Products 3a–
b and in carrageenan obtained by direct extraction was detected with absorption bands observed at 800–805 and 840–850 cm
−1 characteristic for DA2S and G4S respectively (
Figure 1). Typical absorption bands at 1210–1260 cm
−1 common to all types of compounds containing sulfate, and one band at 930 cm
−1 specific to 3,6-anhydro-
d-galactose were also present [
51]. Bands at 825 and 867 cm
−1 specific of the precursor μ- and ν-linked to C6 (D6S) were not observed.
The ι-carrageenan structure in
Products 3a–
b and in carrageenan obtained by
S. filiformis direct extraction was confirmed by
13C nuclear magnetic resonance (NMR) spectra (
Figure 2). Carbohydrates signals were noted at 63.69 and 104.49 ppm. Typical signals of ι-carrageenan for anomeric carbon (C1) of G4S at 104.49 ppm and anomeric carbon (C1) of DA2S at 94.37 ppm [
52] were observed. Other typical signals of G4S at 63.69, 71.43 and 74.38 ppm attributed to carbon C6, C2 and C4 were observed. Peaks at 72.12, 74.37 and 79.3 ppm of DA2S attributed to C2, C4 and C5 were also detected [
8,
21,
22,
52]. Characteristic signals of anomeric carbons for other polysaccharides or for the μ- and ν-precursors were not detected, indicating the presence of a fairly pure ι-carrageenan.
The cytotoxicity and in vitro antiviral activity of
Products 3a–
b and those obtained from carrageenan by direct extraction are presented in
Table 7. Similar to the results obtained from
Products 1a–
b (water-soluble extracts), no cytotoxic effect on the Vero cells was observed in the range of the concentrations assayed. After 3 days of treatment, no microscopically visible alteration of normal cell morphology was observed and viability assay showed no destruction of the cell layer. All carrageenans (
Products 3a–
b and that obtained from direct extraction) exhibited a powerful antiviral activity against
Herpes simplex virus, higher to that showed by
Product 1a, and very close to that showed by acyclovir.
It is noteworthy that the strongest antiviral activity was obtained by
Product 3b, which showed the highest sulfate content and lowest 3,6-anhydro-galactose (34.9% and 7.43% for sulfate and 3,6-anhydro-galactose, respectively). As mentioned above, previous studies have shown that degree of sulfation has a major impact on the antiviral activity of sulfated polysaccharides because of its ability to interfere with the initial attachment of the virus to the target cell [
53,
54]. Recently a synergistic antiviral effect of sulfated polysaccharides extracted from the brown seaweed Eisenia arborea and
S. filiformis on the enveloped measles virus has been reported [
20]. Authors reported that the most efficient inhibition was obtained in the early phases of infection (0 and 15 min after infection). Furthermore, it has been suggested that the conformational flexibility of the binding region plays an important role in the antiviral activity of sulfated polysaccharides, and the chain flexibility of random coil carrageenans increases with the percentage of 3,6-anhydro-galactose [
55,
56]. According to our results, the lower 3,6-anhydro-galactose content found in
Product 3b would indicate that a decrease in chain flexibility could be related to an increase in the antiviral activity. However, besides the degree of sulfation other factors seems to be involved in the formation of the polysaccharide-virus complex such as the molecular weight and the distribution of sulfate groups [
54]. In this way, more chemical and structural analyzes of this promising carrageenan should be carried out, as well as studies on its mechanism of action as an antiviral agent.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}