Optimum Production Conditions, Purification, Identification, and Antioxidant Activity of Violaxanthin from Microalga Eustigmatos cf. polyphem (Eustigmatophyceae)


Abstract
:1. Introduction
2. Results and Discussion
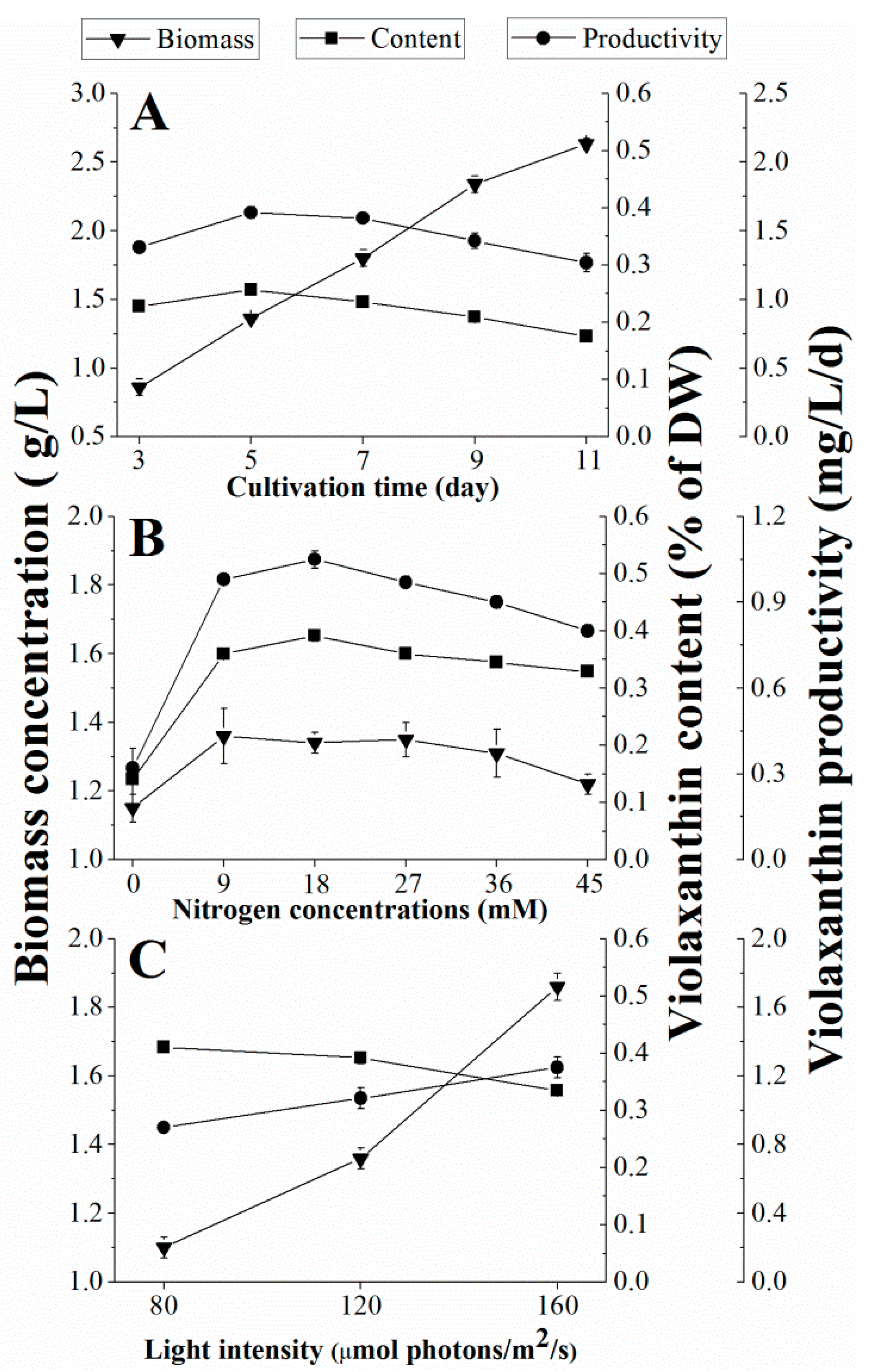
2.1. Effect of Cultivation Time on Violaxanthin Production
2.2. Effect of Nitrogen Concentration on Violaxanthin Production
2.3. Effect of Light Intensity on Violaxanthin Production
2.4. Effect of Culture Mode on Violaxanthin Production
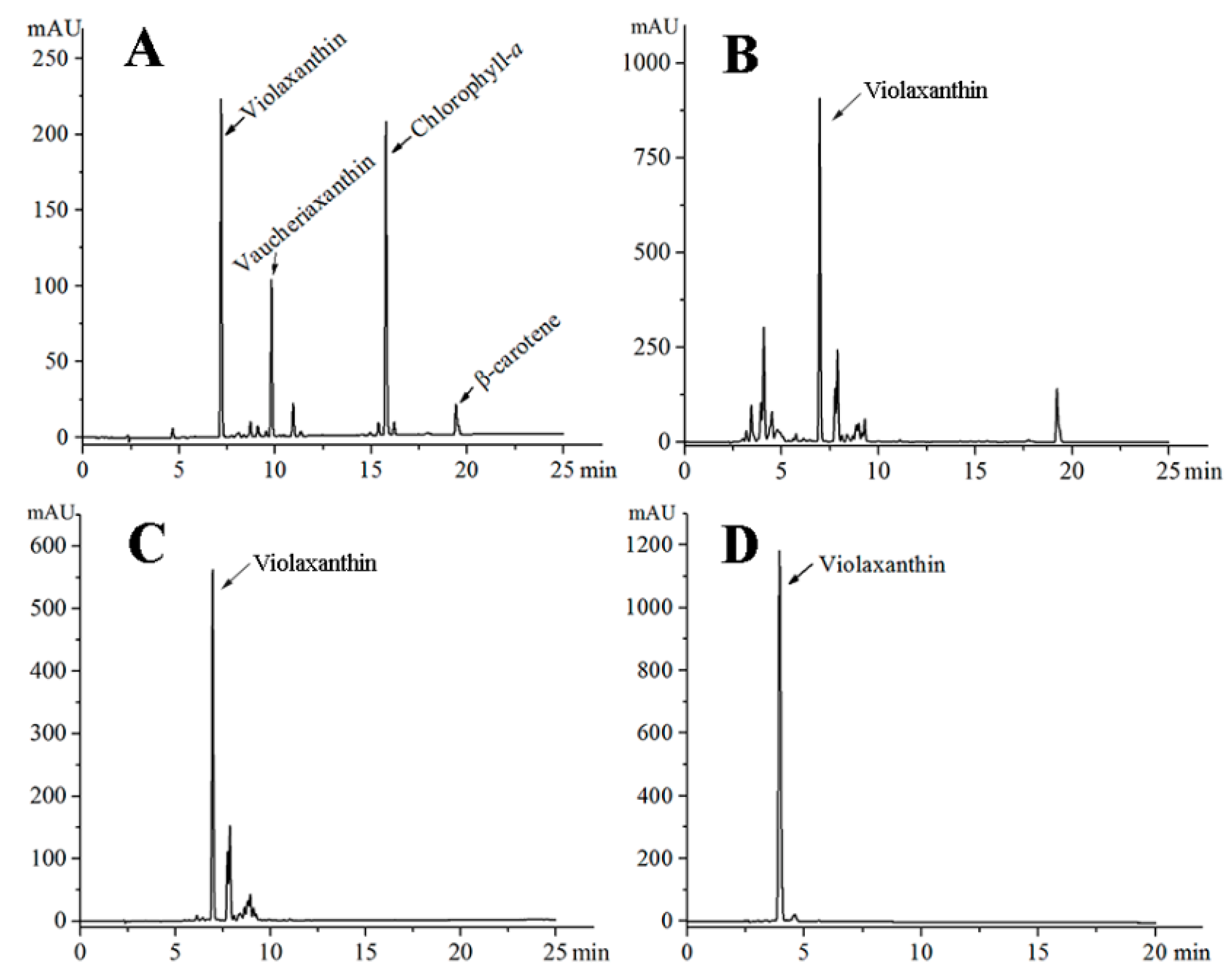
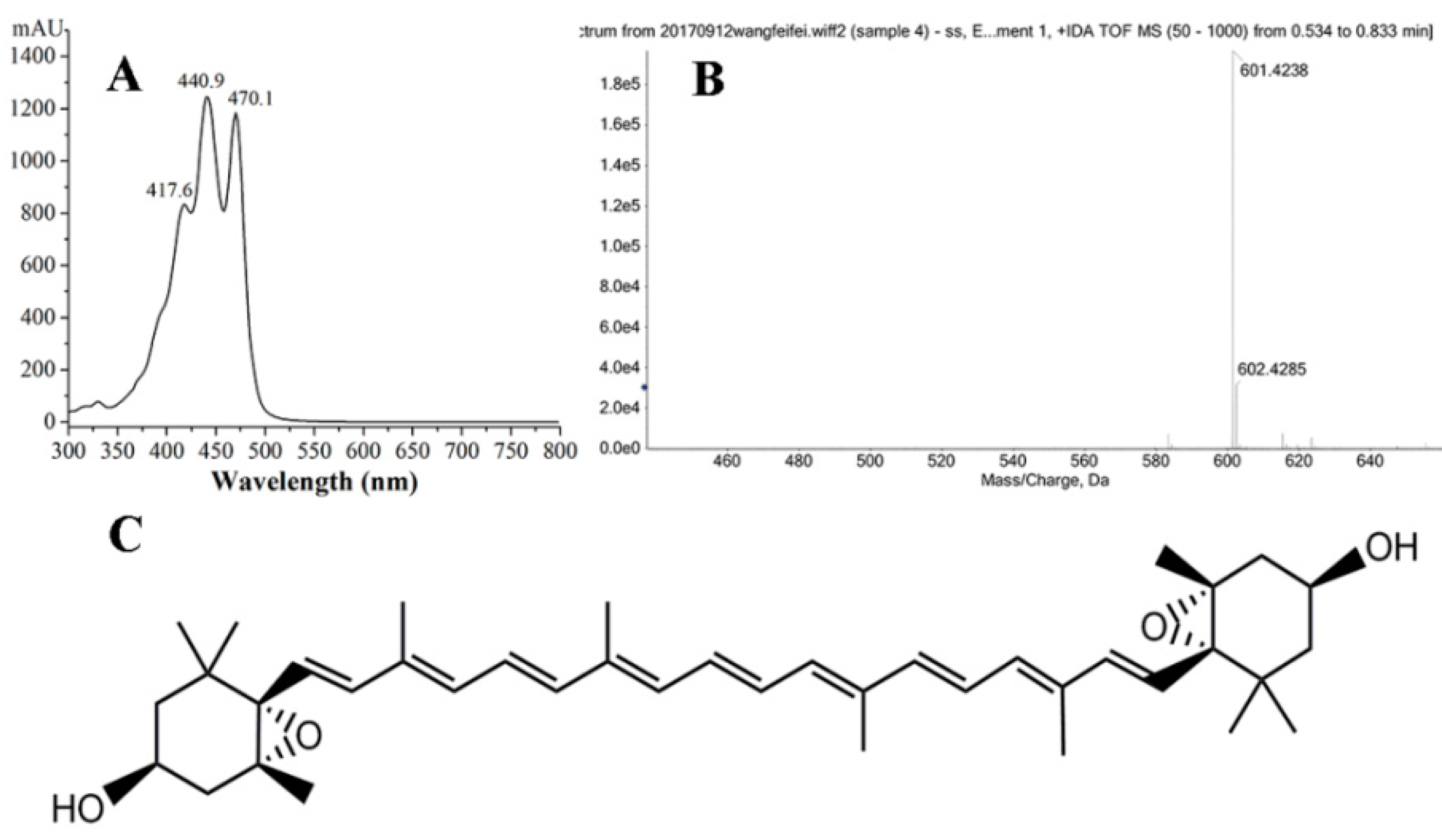
2.5. Purification and Identification of Violaxanthin
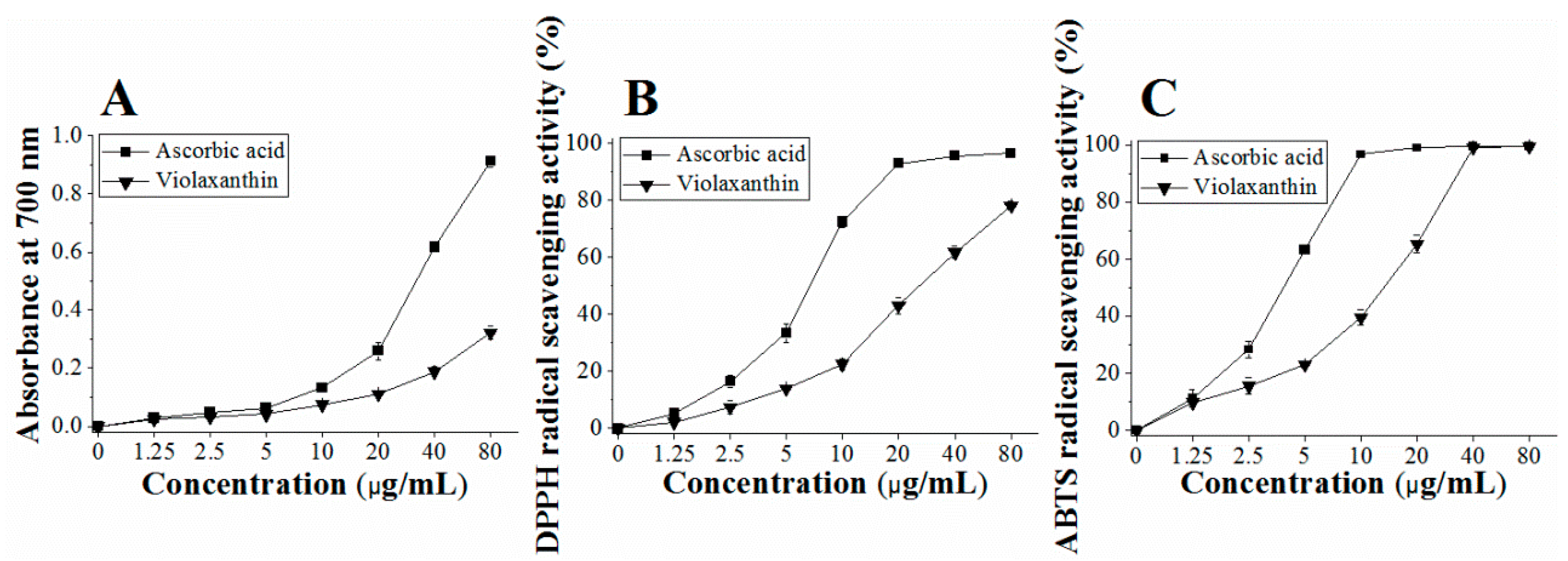
2.6. Reducing Power of Purified Violaxanthin
2.7. DPPH Radical-Scavenging Activity of Purified Violaxanthin
2.8. ABTS Radical Scavenging Activity of the Purified Violaxanthin
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. General Analytical Methods
3.3. Microalgae and Cultivation Conditions
3.4. Extraction, Isolation and Purification of Violaxanthin
3.5. HPLC Analysis and Identification of Purified Violaxanthin
3.6. Antioxidant Activity Assay
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rodriguez-Amaya, D.B. A Guide to Carotenoid Analysis in Foods; ILSI Press: Washington, DC, USA, 2001. [Google Scholar]
- Fu, H.; Xie, B.; Ma, S.; Zhu, X.; Fan, G.; Pan, S. Evaluation of antioxidant activities of principal carotenoids available in water spinach (Ipomoea aquatica). J. Food. Compos. Anal. 2011, 24, 288–297. [Google Scholar] [CrossRef]
- Liu, D.; Shi, J.; Ibarra, A.C.; Kakuda, Y.; Xue, S.J. The scavenging capacity and synergistic effects of lycopene, vitamin E, vitamin C, and β-carotene mixtures on the DPPH free radical. LWT-Food Sci. Technol. 2008, 41, 1344–1349. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 2004, 43, 228–265. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J. Commercial application of microalgae other than as biofuels: A brief review. Rev. Environ. Sci. Bio/Technol. 2011, 10, 31–41. [Google Scholar] [CrossRef]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Characterization of dietary fucoxanthin from Himanthalia elongata brown seaweed. Food Res. Int. 2017, 99, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.; Gómez-Maqueo, A.; García-Cayuela, T.; Welti-Chanes, J. Characterization of carotenoid profile of Spanish Sanguinos and Verdal prickly pear (Opuntia ficus-indica, spp.) tissues. Food Chem. 2017, 237, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.C.; Lin, J.T.; Lu, F.J.; Chou, F.P.; Yang, D.J. Determination of carotenoids in Dunaliella salina cultivated in Taiwan and antioxidant capacity of the algal carotenoid extract. Food Chem. 2008, 109, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-inflammatory effects of violaxanthin isolated from microalga Chlorella ellipsoidea in RAW 264.7 macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Hagen, C.; Braune, W.; Vogel, K.; HÄDER, D.P. Functional aspects of secondary carotenoids in Haematococcus lacustris (Girod) Rostafinski (Volvocales). V. Influences on photomovement. Plant Cell Environ. 1993, 16, 991–995. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Britton, G.; Vicario, I.M.; Heredia, F.J. The complex carotenoid pattern of orange juices from concentrate. Food Chem. 2008, 109, 546–553. [Google Scholar] [CrossRef]
- Shukla, M.; Kumar, S. Algal biorefineries for biofuels and other value-added products. In Biorefining of Biomass to Biofuels; Springer: Cham, Switzerland, 2018; pp. 305–341. [Google Scholar]
- Pasquet, V.; Morisset, P.; Ihammouine, S.; Chepied, A.; Aumailley, L.; Berard, J.-B.; Serive, B.; Kaas, R.; Lanneluc, I.; Thiery, V. Antiproliferative activity of violaxanthin isolated from bioguided fractionation of Dunaliella tertiolecta extracts. Mar. Drugs 2011, 9, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wan, L.; Xia, S.; Li, A.; Zhang, C. Morphological and spectrometric analyses of lipids accumulation in a novel oleaginous microalga, Eustigmatos cf. polyphem (Eustigmatophyceae). Bioprocess Biosyst. Eng. 2013, 36, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ma, X.; Li, A.; Zhang, C. A novel potential source of β-carotene: Eustigmatos cf. polyphem (Eustigmatophyceae) and pilot β-carotene production in bubble column and flat panel photobioreactors. Bioresour. Technol. 2012, 117, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Xia, S.; Lei, X.; Zhang, C. Combined effects of different nitrogen sources and levels and light intensities on growth and fatty acid and lipid production of oleaginous eustigmatophycean microalga Eustigmatos cf. polyphem. J. Appl. Phycol. 2018, 30, 215–229. [Google Scholar] [CrossRef]
- Eliáš, M.; Amaral, R.; Fawley, K.P.; Fawley, M.W.; Němcová, Y.; Neustupa, J.; Přibyl, P.; Santos, L.M.; Ševčíková, T. Handbook of the Protists; Springer International Publishing: Basel, Switzerland, 2017; pp. 1–39. [Google Scholar]
- Keşan, G.; Litvín, R.; Bína, D.; Durchan, M.; Šlouf, V.; Polívka, T. Efficient light-harvesting using non-carbonyl carotenoids: Energy transfer dynamics in the VCP complex from Nannochloropsis oceanica. BBA-Bioenerg. 2016, 1857, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Llansola-Portoles, M.J.; Litvin, R.; Ilioaia, C.; Pascal, A.A.; Bina, D.; Robert, B. Pigment structure in the violaxanthin–chlorophyll-a-binding protein VCP. Photosynth. Res. 2017, 134, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Bína, D.; Gardian, Z.; Herbstová, M.; Litvín, R. Modular antenna of photosystem I in secondary plastids of red algal origin: A Nannochloropsis oceanica case study. Photosynth. Rese. 2017, 131, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.W. Xanthophyll cycle and light stress in nature: Uniform response to excess direct sunlight among higher plant species. Planta 1996, 198, 460–470. [Google Scholar] [CrossRef]
- Uhrmacher, S.; Hanelt, D.; Nultsch, W. Zeaxanthin content and the degree of photoinhibition are linearly correlated in the brown alga Dictyota dichotoma. Mar. Biol. 1995, 123, 159–165. [Google Scholar] [CrossRef]
- Gao, B.; Yang, J.; Lei, X.; Xia, S.; Li, A.; Zhang, C. Characterization of cell structural change, growth, lipid accumulation, and pigment profile of a novel oleaginous microalga, Vischeria stellata (Eustigmatophyceae), cultured with different initial nitrate supplies. J. Appl. Phycol. 2016, 28, 821–830. [Google Scholar] [CrossRef]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; González-del Valle, M.; Parés, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Peccia, J.; Haznedaroglu, B.; Gutierrez, J.; Zimmerman, J.B. Nitrogen supply is an important driver of sustainable microalgae biofuel production. Trends Biotechnol. 2013, 31, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.R.; Cheng, S.J.; Qi, G.H.; Yang, Z.P.; Yin, S.Y.; Chen, G.T. Antimicrobial and antioxidant activities of Flammulina velutipes polysacchrides and polysacchride-iron (III) complex. Carbohydr. Polym. 2017, 161, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Zhang, J.; Hou, X.; Ahmad, H.; Zhang, H.; Zhang, L.; Wang, T. Assessment of free radicals scavenging activity of seven natural pigments and protective effects in AAPH-challenged chicken erythrocytes. Food Chem. 2014, 145, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Xu, W.; Zhang, J.; Kou, T.; Niu, Y.; Wan, X.; Zhang, L.; Wang, C.; Wang, T. Assessment of free radical scavenging activity of dimethylglycine sodium salt and its role in providing protection against lipopolysaccharide-induced oxidative stress in mice. PLoS ONE 2016, 11, e0155393. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lin, J.; Gao, Y.; Han, W.; Chen, D. Antioxidant activity and mechanism of Rhizoma Cimicifugae. Chem. Cent. J. 2012, 6, 140–150. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Mode | Time (Day) | Biomass (g/L) | Violaxanthin Content (% of DW) | Violaxanthin Productivity (mg/L/d) |
|---|---|---|---|---|
| Batch culture | 5th | 1.36 ± 0.03 a | 0.39 ± 0.01 a | 1.07 ± 0.03 a |
| (control) | 10th | 2.53 ± 0.04 b | 0.32 ± 0.01 b | 0.82 ± 0.03 b |
| 15th | 3.39 ± 0.06 c | 0.27 ± 0.02 c | 0.62 ± 0.04 c | |
| 20th | 4.37 ± 0.03 d | 0.23 ± 0.01 d | 0.51 ± 0.02 d | |
| Fed-batch | 5th | 1.36 ± 0.03 a | 0.39 ± 0.02 a | 1.07 ± 0.03 a |
| culture | 10th | 2.92 ± 0.01 e | 0.38 ± 0.01 e | 1.10 ± 0.03 a |
| 15th | 4.02 ± 0.05 f | 0.36 ± 0.01 f | 0.96 ± 0.02 e | |
| 20th | 5.15 ± 0.03 g | 0.35 ± 0.02 f | 0.90 ± 0.06 be |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Huang, L.; Gao, B.; Zhang, C. Optimum Production Conditions, Purification, Identification, and Antioxidant Activity of Violaxanthin from Microalga Eustigmatos cf. polyphem (Eustigmatophyceae). Mar. Drugs 2018, 16, 190. https://doi.org/10.3390/md16060190
Wang F, Huang L, Gao B, Zhang C. Optimum Production Conditions, Purification, Identification, and Antioxidant Activity of Violaxanthin from Microalga Eustigmatos cf. polyphem (Eustigmatophyceae). Marine Drugs. 2018; 16(6):190. https://doi.org/10.3390/md16060190
Chicago/Turabian StyleWang, Feifei, Luodong Huang, Baoyan Gao, and Chengwu Zhang. 2018. "Optimum Production Conditions, Purification, Identification, and Antioxidant Activity of Violaxanthin from Microalga Eustigmatos cf. polyphem (Eustigmatophyceae)" Marine Drugs 16, no. 6: 190. https://doi.org/10.3390/md16060190
APA StyleWang, F., Huang, L., Gao, B., & Zhang, C. (2018). Optimum Production Conditions, Purification, Identification, and Antioxidant Activity of Violaxanthin from Microalga Eustigmatos cf. polyphem (Eustigmatophyceae). Marine Drugs, 16(6), 190. https://doi.org/10.3390/md16060190