Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic


Abstract
:1. Introduction
2. Results
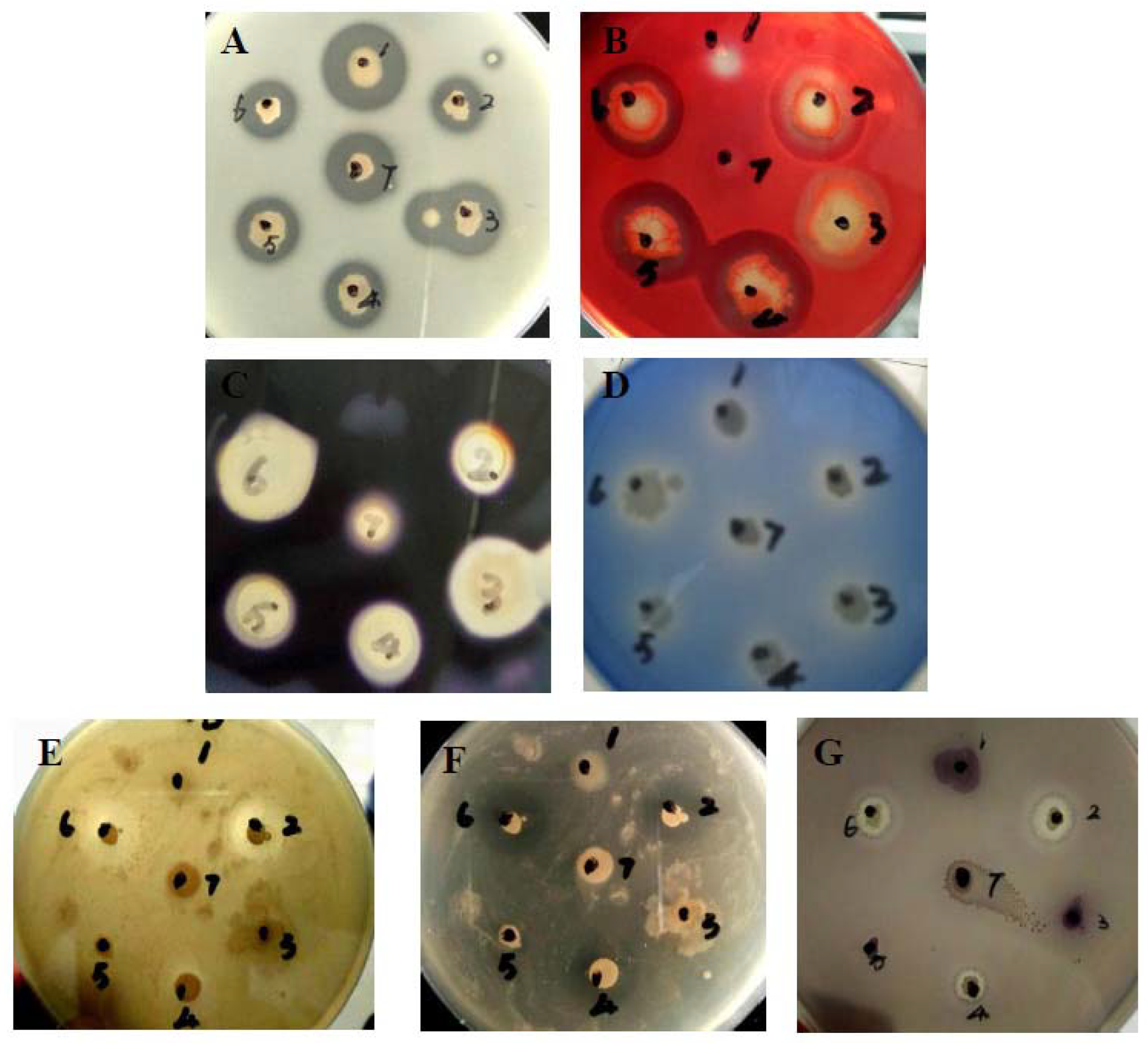
2.1. Isolation of Marine Antagonistic, Quorum-Quenching and Multienzymes Producing Strains
2.2. Characterization and Identification of YB1701
2.3. Acid and Bile Salt Tolerance
2.4. Safety Assessment of YB1701
2.5. Antibiotic Susceptibility
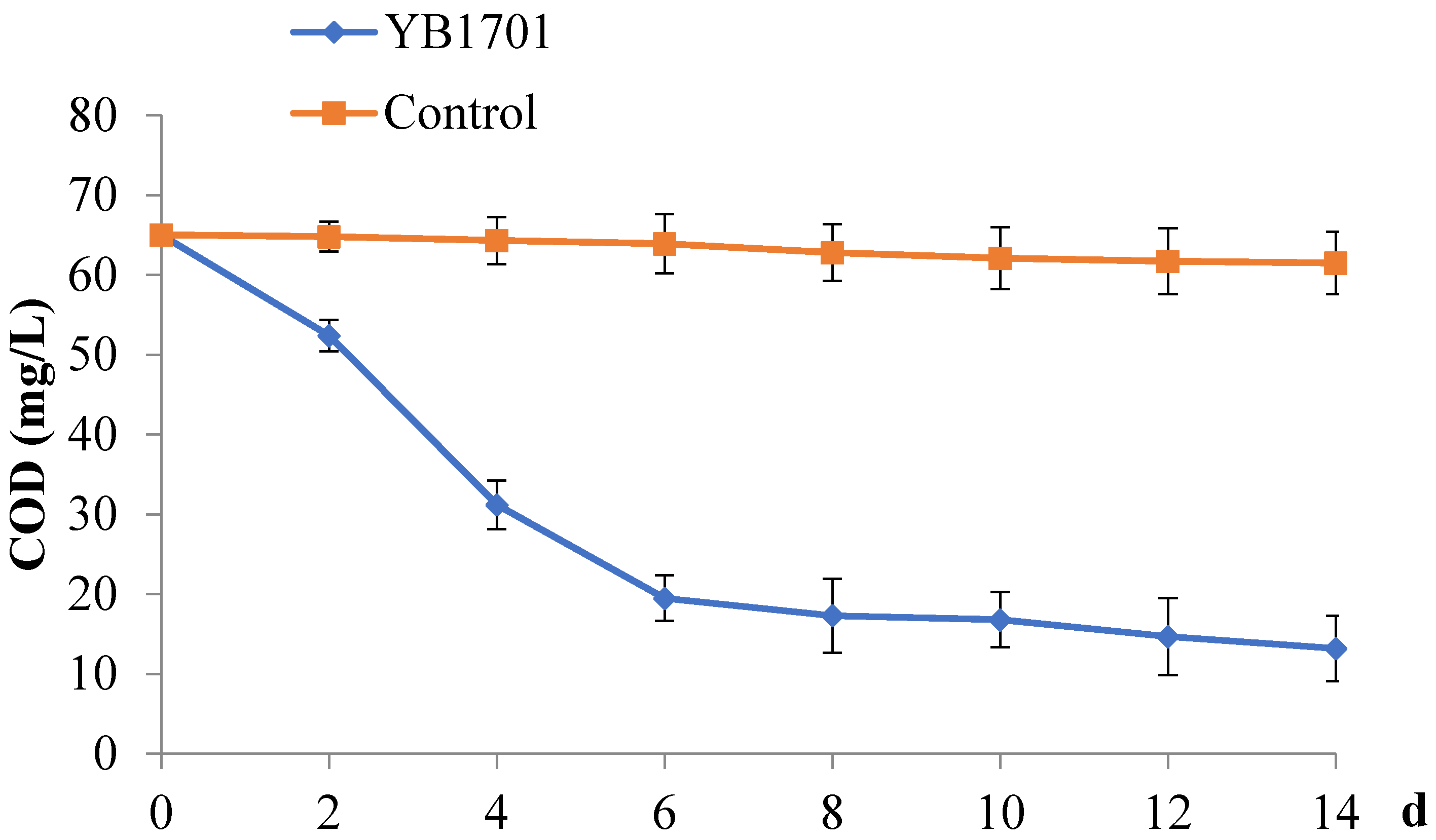
2.6. Ability of YB1701 to Degrade Organic Pollutants
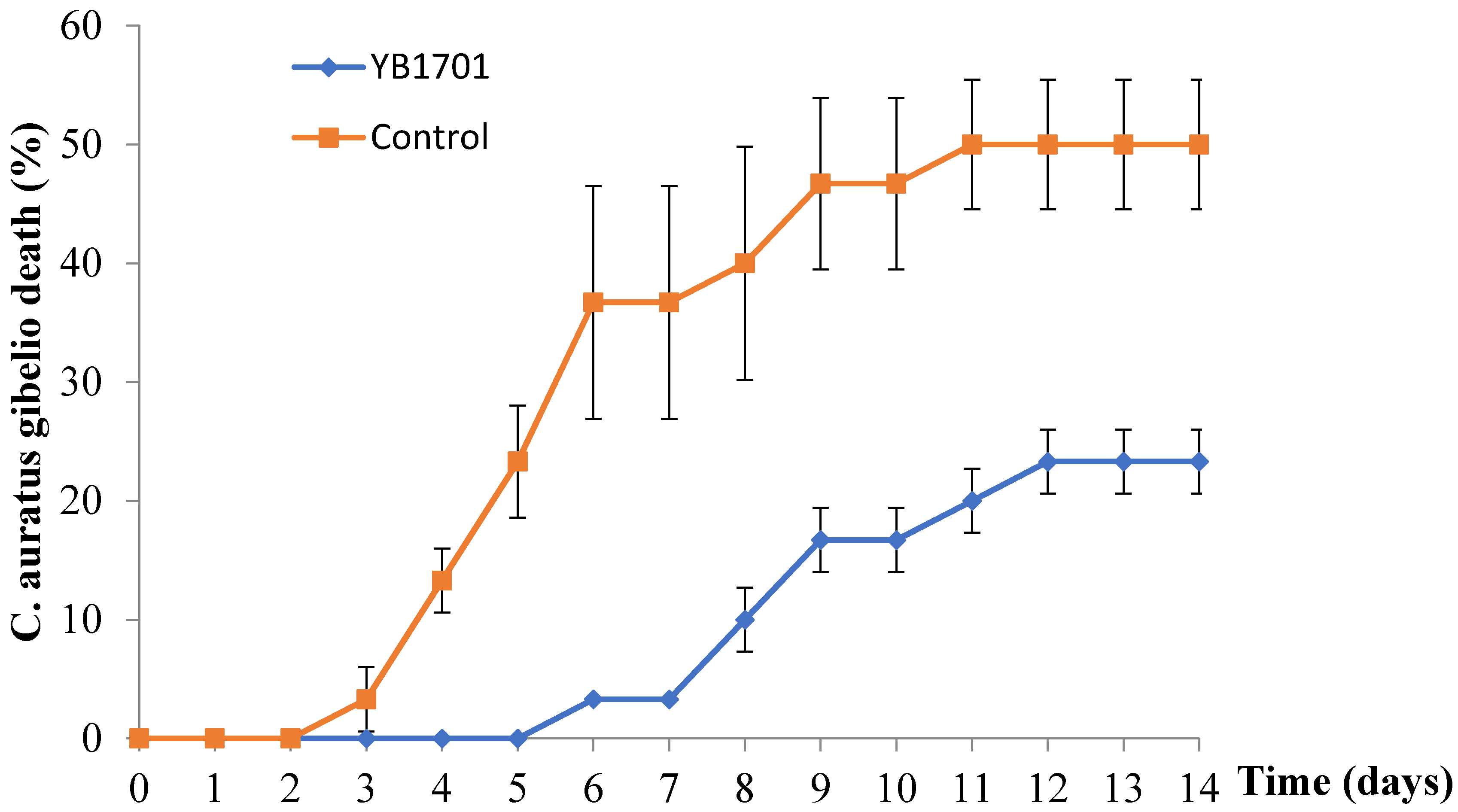
2.7. In Vivo Protective Effect of YB1701 on A. hydrophila Infection
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Isolation of Bacteria
4.2. Antimicrobial, Quorum-Quenching and Extracellular Enzymatic Activity Screening
4.2.1. Antimicrobial Activity Assay
4.2.2. Bioassay of Quorum-Quenching Activity
4.2.3. Extracellular Enzymatic Activities Assay
4.3. Morphological and Physiological Characteristics and 16S rRNA Gene Amplification of Potential Probiotic Strain
4.4. In Vitro Acid and Bile Salt Tolerance Assay
4.5. Safety Evaluation of YB1701
4.6. Antibiotic Susceptibility Assay
4.7. Degradation Rates of YB1701 for Organics in Wastewater of Aquaculture
4.8. Effect of YB1701 Supplemented Fish Diet on Disease Resistance
4.9. Ethics Statement
5. Statistical Analysis
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture, 2010; Food and Agriculture Organization (FAO): Rome, Italy, 2012. [Google Scholar]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The rising tide of antimicrobial resistance in aquaculture: Sources, sinks and solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.I.; Sadekuzzaman, M.; Ha, S.D. Probiotics as potential alternative biocontrol agents in the agriculture and food industries: A review. Food Res. Int. 2017, 100 Pt 1, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO)/World Health Organization (WHO). Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations and World Health Organization Working Group Report; Food and Agriculture Organization: Rome, Italy, 2002. [Google Scholar]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.M.; Ibáñez, A.L.; Hermosillo, O.A.M.; Saad, H.C.R. Use of probiotics in aquaculture. ISRN Microbiol. 2012, 2012, 916845. [Google Scholar] [CrossRef]
- Huynh, T.G.; Shiu, Y.L.; Nguyen, T.P.; Truong, Q.P.; Chen, J.C.; Liu, C.H. Current applications, selection, and possible mechanisms of actions of synbiotics in improving the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 2017, 64, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, G.; Ray, A.K. The advancement of probiotics research and its application in fish farming industries. Res. Vet. Sci. 2017, 115, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.C.; Lin, H.L.; Shiu, Y.L.; Tyan, Y.C.; Liu, C.H. Isolation and characterization of antimicrobial peptides derived from Bacillus subtilis E20-fermented soybean meal and its use for preventing Vibrio infection in shrimp aquaculture. Fish Shellfish Immunol. 2017, 67, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.T.; Kamilya, D.; Kheti, B.; Bordoloi, B.; Parhi, J. Paraprobiotic preparation from Bacillus amyloliquefaciens FPTB16 modulates immune response and immune relevant gene expression in Catla catla (Hamilton, 1822). Fish Shellfish Immunol. 2017, 66, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zorriehzahra, M.J.; Delshad, S.T.; Ade, L.M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Bentzon-Tilia, M.; Sonnenschein, E.C.; Gram, L. Monitoring and managing microbes in aquaculture—Towards a sustainable industry. Microb. Biotechnol. 2016, 9, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, L.; Hernández, R.; Sainz, E.; González Cervantes, R.; Martínez Cruz, P.; Mayorga Reyes, L.; Azaola Espinosa, A. Cambio en la flora intestinal de ratones por la administraci ´on de bifidobacterias y jugo de girasol. Vet. Méx. 1996, 27, 127–131. [Google Scholar]
- Tan, L.T.; Chan, K.G.; Lee, L.H.; Goh, B.H. Streptomyces bacteria as potential probiotics in aquaculture. Front. Microbiol. 2016, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: Status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, M.; Quan, C.S.; Fan, S.D. Mechanisms of quorum sensing and strategies for quorum sensing disruption in aquaculture pathogens. J. Fish Dis. 2015, 38, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.A.; Duc le, H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.Y.; Liu, Y.; Miao, L.L.; Li, E.W.; Sun, G.X.; Liu, Y.; Liu, Z.P. Characterization and mechanism of anti-Aeromonas salmonicida activity of a marine probiotic strain, Bacillus velezensis V4. Appl. Microbiol. Biotechnol. 2017, 101, 3759–3768. [Google Scholar] [CrossRef] [PubMed]
- Gobi, N.; Vaseeharan, B.; Chen, J.C.; Rekha, R.; Vijayakumar, S.; Anjugam, M.; Iswarya, A. Dietary supplementation of probiotic Bacillus licheniformis Dahb1 improves growth performance, mucus and serum immune parameters, antioxidant enzyme activity as well as resistance against Aeromonas hydrophila in tilapia Oreochromis mossambicus. Fish Shellfish Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ran, C.; Carrias, A.; Williams, M.A.; Capps, N.; Dan, B.C.T.; Newton, J.C.; Kloepper, J.W.; Ooi, E.L.; Browdy, C.L.; Terhune, J.S.; et al. Identification of Bacillus strains for biological control of catfish pathogens. PLoS ONE 2012, 7, e45793. [Google Scholar] [CrossRef] [PubMed]
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of probiotic Bacillus aerius B81e isolated from healthy hybrid catfish on growth, disease resistance and innate immunity of Pla-mong Pangasius bocourti. Fish Shellfish Immunol. 2017, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, D.; Souissi, S.; Ahamed, T.S. Effects of the potential probiotics Bacillus aerophilus KADR3 in inducing immunity and disease resistance in Labeo rohita. Fish Shellfish Immunol. 2017, 70, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Meidong, R.; Doolgindachbaporn, S.; Jamjan, W.; Sakai, K.; Tashiro, Y.; Okugawa, Y.; Tongpim, S. A novel probiotic Bacillus siamensis B44v isolated from Thai pickled vegetables (Phak-dong) for potential use as a feed supplement in aquaculture. J. Gen. Appl. Microbiol. 2017, 63, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Srisapoome, P.; Areechon, N. Efficacy of viable Bacillus pumilus isolated from farmed fish on immune responses and increased disease resistance in Nile tilapia (Oreochromis niloticus): Laboratory and on-farm trials. Fish Shellfish Immunol. 2017, 67, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Hao, K.; Wu, Z.Q.; Li, D.L.; Yu, X.B.; Wang, G.X.; Ling, F. Effects of dietary administration of Shewanella xiamenensis A-1, Aeromonas veronii A-7, and Bacillus subtilis, single or combined, on the grass Carp (Ctenopharyngodon idella) intestinal microbiota. Probiotics Antimicrob. Proteins 2017, 9, 386–396. [Google Scholar] [CrossRef] [PubMed]
- De, B.C.; Meena, D.K.; Behera, B.K.; Das, P.; Das Mohapatra, P.K.; Sharma, A.P. Probiotics in fish and shellfish culture: Immunomodulatory and ecophysiological responses. Fish Physiol. Biochem. 2014, 40, 921–971. [Google Scholar] [CrossRef]
- Mondol, M.A.; Shin, H.J.; Islam, M.T. Diversity of secondary metabolites from marine Bacillus species: Chemistry and biological activity. Mar. Drugs 2013, 11, 2846–2872. [Google Scholar] [CrossRef] [PubMed]
- 35Zhang, D.S.; Li, H.; Liu, Y.; Qiao, G.; Chi, S.; Song, J. Screening and identification of organics-degrading bacteria from the sediment of sea cucumber Apostichopus japonicus ponds. Aquacult. Int. 2016, 24, 373–384. [Google Scholar] [CrossRef]
- Al-Wasify, R.S.; Ali, M.N.; Hamed, S.R. Biodegradation of dairy wastewater using bacterial and fungallocal isolates. Water Sci. Technol. 2017, 76, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.J.; Tian, X.L.; Dong, S.G.; Wu, X.H.; Liu, B.G. Isolation, selection and characterization of low temperature organic-pollutant-degrading bacteria in sea cucumber ponds. J. Fish Sci. China 2012, 19, 626–634. [Google Scholar] [CrossRef]
- Chu, W.H. Studies on the pathology and control of the bacterial hemorrhage disease in a hybrid crussian carp. Reserv. Fish. 2001, 21, 40. [Google Scholar]
- Chu, W.H.; Zhu, W.; Kang, C.T. Isolation, identification of marine Bdellovibrios and its effect on Vibrio parahaemolyticus. Microbiology 2009, 36, 20–24. [Google Scholar]
- Perez, C.; Paul, M.; Bazerque, P. An antibiotic assay by the agar well diffusion method. Acta Biol. Med. Exp. 1990, 15, 113–115. [Google Scholar]
- McLean, R.J.; Pierson, L.S.; Fuqua, C. A simple screening protocol for the identification of quorum signal antagonists. J. Microbiol. Methods 2004, 58, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.V.; Vijayan, K.K.; Chakrabort, K.; Antony, M.L. Diversity and characterization of antagonistic bacteria from tropical estuarine habitats of Cochin, India for fish health management. World J. Microbiol. Biotechnol. 2012, 28, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Dlusskaya, E.A.; McMullen, L.M.; Gänzle, M.G. Characterization of an extremely heat-resistant Escherichia coli obtained from a beef processing facility. J. Appl. Microbiol. 2011, 110, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbour Laboratory: Cold Spring Harbor, NY, USA, 1982. [Google Scholar]
- Duc, L.H.; Hong, H.A.; Barbosa, T.M.; Henriques, A.O.; Cutting, S.M. Characterization of Bacillus probiotics available for human use. Appl. Environ. Microbiol. 2004, 70, 2161–2171. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- NCCLS. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, Approved Standard, 9th ed.; 2012; Available online: http://antimicrobianos.com.ar/ATB/wp-content/uploads/2012/11/03-CLSI-M07-A9-2012.pdf (accessed on 3 November 2012).
- EFSA. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Latif-Eugenín, F.; Beaz-Hidalgo, R.; Figueras, M.J. Evaluation of different conditions and culture media for the recovery of Aeromonas spp. from water and shellfish samples. J. Appl. Microbiol. 2016, 121, 883–891. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Bacillus sp. YB1701 |
|---|---|
| Morphological | |
| Shape | Rods |
| Gram stain | G+ |
| Motility | Motile |
| Spore formation | + |
| Growth | |
| Growth temperature | 15 °C–50 °C |
| Growth pH | 5–8 |
| Growth in 10% NaCl | + |
| Aerobic growth | + |
| Anaerobic growth | + |
| Biochemical tests | |
| Catalase | + |
| Oxidase | + |
| Voges-Proskauer | + |
| Indole production | - |
| Methyl red test | - |
| Citrate | + |
| Catalase reaction | + |
| Glucosamine | + |
| Nitrate reduction | + |
| H2S production | + |
| Crystalline dextrin production test | - |
| Fermentative | |
| Glucose | + |
| Fructose | + |
| Lactose | - |
| Mannose | - |
| Raffinose | + |
| Hydrolysis of | |
| Casein | + |
| Gelatin | + |
| Starch | + |
| - | Log10 CFU of Viable Bacteria per mL | - | Log10 CFU of Viable Bacteria per mL | ||||
|---|---|---|---|---|---|---|---|
| Time/h | pH 6.5 | pH 3.0 | pH 2.0 | Time/h | 0.0% bile | 1.0% bile | 2.0% bile |
| 0 | 7.98 | 7.98 | 7.98 | 0 | 8.76 | 8.76 | 8.76 |
| 1 | 7.89 | 7.63 | 7.25 | 1 | 8.15 | 8.13 | 7.94 |
| 2 | 8.14 | 7.18 | 7.18 | 3 | 7.96 | 7.95 | 7.32 |
| 3 | 8.27 | 7.03 | 6.84 | 12 | 7.82 | 7.87 | 7.47 |
| Antibiotic Discs * | Inhibition Zone Diameter (mm) # | Susceptibility |
|---|---|---|
| Gentamicin | 23.4 ± 0.4 | S |
| Kanamycin | 27.0 ± 0.2 | S |
| Clindamycin | 25.4 ± 1.4 | S |
| Chloramphenicol | 22.6 ± 0.6 | S |
| Erythromycin | 27.5 ± 0.5 | S |
| Streptomycin | 21.7 ± 0.7 | S |
| Tetracycline | 13.4 ± 0.6 | S |
| Vancomycin | 20.2 ± 1.1 | S |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Xia, Y.; Zhu, C.; Chu, W. Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic. Mar. Drugs 2018, 16, 196. https://doi.org/10.3390/md16060196
Zhou S, Xia Y, Zhu C, Chu W. Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic. Marine Drugs. 2018; 16(6):196. https://doi.org/10.3390/md16060196
Chicago/Turabian StyleZhou, Shuxin, Yu Xia, Chongmiao Zhu, and Weihua Chu. 2018. "Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic" Marine Drugs 16, no. 6: 196. https://doi.org/10.3390/md16060196
APA StyleZhou, S., Xia, Y., Zhu, C., & Chu, W. (2018). Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic. Marine Drugs, 16(6), 196. https://doi.org/10.3390/md16060196