Marine Skeletons: Towards Hard Tissue Repair and Regeneration



{kind=link}
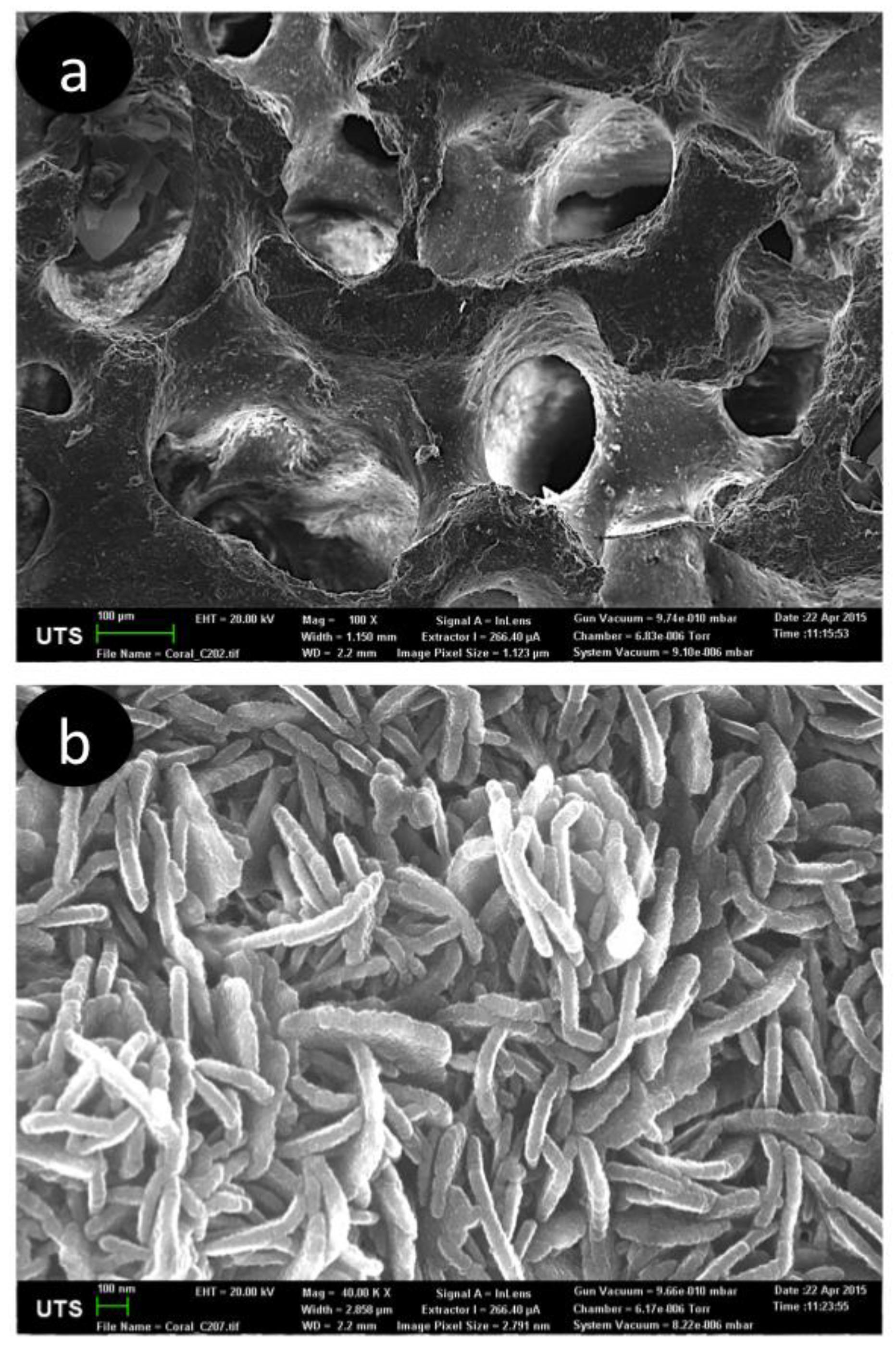{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Marine Skeletons
3. Seashells
4. Corals
4.1. Coral Skeletons in Dentistry
4.2. Coral Skeletons as a Drug Carrier
5. Sea Urchins
6. Cuttlebone
7. Marine Sponges
8. Concluding Remarks and Future Perspectives
Conflicts of Interest
References
- Amini, A.R.; Laurencin, C.T.; Nukavarapu, S.P. Bone Tissue Engineering: Recent Advances and Challenges. Crit. Rev. Biomed. Eng. 2012, 40, 363–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oryan, A.; Alidadi, S.; Moshiri, A.; Maffulli, N. Bone regenerative medicine: Classic options, novel strategies, and future directions. J. Orthop. Surg. Res. 2014, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Libonati, F.; Vergani, L. Cortical Bone as a Biomimetic Model for the Design of New Composites. Procedia Struct. Integr. 2016, 2, 1319–1326. [Google Scholar] [CrossRef]
- Place, E.S.; Evans, N.D.; Stevens, M.M. Complexity in biomaterials for tissue engineering. Nat. Mat. 2009, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, B. Discovery and development of marine biomaterials. In Functional Marine Biomaterials, 1st ed.; Kim, S.-K., Ed.; Elsevier Ltd.: Cambridge, UK, 2015; pp. 3–28. [Google Scholar]
- Webster, T.J.; Ahn, E.S. Nanostructured Biomaterials for Tissue Engineering Bone. In Tissue Engineering II: Basics of Tissue Engineering and Tissue Application; Lee, K., Kaplan, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 275–308. [Google Scholar]
- Keaveny, T.M.; Morgan, E.F.; Niebur, G.L.; Yeh, O.C. Biomechanics of Trabecular Bone. Ann. Rev. Biomed. Eng. 2001, 3, 307–333. [Google Scholar] [CrossRef] [PubMed]
- Barrère, F.; Mahmood, T.A.; de Groot, K.; van Blitterswijk, C.A. Advanced biomaterials for skeletal tissue regeneration: Instructive and smart functions. Mater. Sci. Eng. R Rep. 2008, 59, 38–71. [Google Scholar] [CrossRef]
- Macha, I.J.; Cazalbou, S.; Ben-Nissan, B.; Harvey, K.L.; Milthorpe, B. Marine structure derived calcium phosphate-polymer biocomposites for local antibiotic delivery. Mar. Drugs 2015, 13, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Macha, I.J.; Ozyegin, L.S.; Chou, J.; Samur, R.; Oktar, F.N.; Ben-Nissan, B. An Alternative Synthesis Method for Di Calcium Phosphate (Monetite) Powders from Mediterranean Mussel (Mytilus galloprovincialis) Shells. J. Aust. Ceram. Soc. 2013, 49, 122–128. [Google Scholar]
- Choi, A.H.; Macha, I.J.; Akyol, S.; Cazalbou, S.; Ben-Nissan, B. Nanostructured Calcium Phosphates for Drug, Gene, DNA and Protein Delivery and as Anticancer Chemotherapeutic Devices. In Clinical Applications of Biomaterials: State-of-the-Art Progress, Trends, and Novel Approaches; Kaur, G., Ed.; Springer International Publishing: Cham, The Netherland, 2017; pp. 227–256. [Google Scholar]
- Macha, I.J.; Boonyang, U.; Cazalbou, S.; Ben-Nissan, B.; Charvillat, C.; Oktar, F.N.; Grossin, D. Comparative study of Coral Conversion, Part 2: Microstructural evolution of calcium phosphate. J. Aust. Ceram. Soc. 2015, 51, 149–159. [Google Scholar]
- Ben-Nissan, B. Natural bioceramics: From coral to bone and beyond. Curr. Opin. Solid State Mater. Sci. 2003, 7, 283–288. [Google Scholar] [CrossRef]
- Macha, I.J.; Ozyegin, L.S.; Oktar, F.N.; Ben-Nissan, B. Conversion of Ostrich Eggshells (Struthio camelus) to Calcium Phosphates. J. Aust. Ceram. Soc. 2015, 51, 125–133. [Google Scholar]
- Oktar, F.; Tuyel, U.; Demirkol, N.; Gunduz, O.; Samur, R.; Kannan, S.; Agathopoulos, S. A new safe method to produce bioceramic nano-powders from nacre venus verrucosa. Int. J. Artif. Org. 2010, 33, 467. [Google Scholar]
- Atlan, G.; Balmain, N.; Berland, S.; Vidal, B.; Lopez, É. Reconstruction of human maxillary defects with nacre powder: Histological evidence for bone regeneration. C. R. Acad. Sci. 1997, 320, 253–258. [Google Scholar] [CrossRef]
- Shim, K.-S.; Kim, S.E.; Yun, Y.-P.; Jeon, D.I.; Kim, H.-J.; Park, K.; Song, H.-R. Surface immobilization of biphasic calcium phosphate nanoparticles on 3D printed poly(caprolactone) scaffolds enhances osteogenesis and bone tissue regeneration. J. Ind. Eng. Chem. 2017, 55, 101–109. [Google Scholar] [CrossRef]
- Chou, J.; Ben-Nissan, B.; Green David, W.; Valenzuela Stella, M.; Kohan, L. Targeting and Dissolution Characteristics of Bone Forming and Antibacterial Drugs by Harnessing the Structure of Microspherical Shells from Coral Beach Sand. Adv. Eng. Mater. 2011, 13, 93–99. [Google Scholar] [CrossRef]
- Green, D.W.; Ben-Nissan, B.; Yoon, K.S.; Milthorpe, B.; Jung, H.-S. Natural and Synthetic Coral Biomineralization for Human Bone Revitalization. Trends Biotechnol. 2017, 35, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, B.; Choi, A.H. Sol-gel production of bioactive nanocoatings for medical applications. Part 1: An introduction. Nanomedicine 2006, 1, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, B.; Macha, I.; Cazalbou, S.; Choi, A.H. Calcium phosphate nanocoatings and nanocomposites, part 2: Thin films for slow drug delivery and osteomyelitis. Nanomedicine 2016, 11, 531–554. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, B.; Milev, A.; Vago, R. Morphology of sol-gel derived nano-coated coralline hydroxyapatite. Biomaterials 2004, 25, 4971–4975. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-Y.; Lin, A.Y.-M.; McKittrick, J.; Meyers, M.A. Structure and mechanical properties of crab exoskeletons. Acta Biomater. 2008, 4, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Howard, D.; Yang, X.; Kelly, M.; Oreffo, R.O. Natural marine sponge fiber skeleton: A biomimetic scaffold for human osteoprogenitor cell attachment, growth, and differentiation. Tissue Eng. 2003, 9, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, M.; Robic, A. Structure and Functional Properties of Ulvan, a Polysaccharide from Green Seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, M.J.; Grimes, J.S.; Kennedy, M.P. Coralline hydroxyapatite bone graft substitute in hindfoot surgery. Foot Ankle Int. 2006, 27, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Karacan, I.; Macha, I.J.; Choi, G.; Cazalbou, S.; Ben-Nissan, B. Antibiotic Containing Poly Lactic Acid/Hydroxyapatite Biocomposite Coatings for Dental Implant Applications. Key Eng. Mater. 2017, 758, 120–125. [Google Scholar] [CrossRef]
- Macha, I.J.; Cazalbou, S.; Shimmon, R.; Ben-Nissan, B.; Milthorpe, B. Development and dissolution studies of bisphosphonate (clodronate)-containing hydroxyapatite–polylactic acid biocomposites for slow drug delivery. J. Tissue Eng. Regen. Med. 2017, 11, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.S.; Rao, K.P. Preparation, characterization and in vitro release of gentamicin from coralline hydroxyapatite-gelatin composite microspheres. Biomaterials 2002, 23, 3175–3181. [Google Scholar]
- Kolk, A.; Handschel, J.; Drescher, W.; Rothamel, D.; Kloss, F.; Blessmann, M.; Heiland, M.; Wolff, K.-D.; Smeets, R. Current trends and future perspectives of bone substitute materials—From space holders to innovative biomaterials. J. Cranio-Maxillofac. Surg. 2012, 40, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Alakpa, E.V.; Burgess, K.E.V.; Chung, P.; Riehle, M.O.; Gadegaard, N.; Dalby, M.J.; Cusack, M. Nacre Topography Produces Higher Crystallinity in Bone than Chemically Induced Osteogenesis. ACS Nano 2017, 11, 6717–6727. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhou, H.; Lee, J. Various preparation methods of highly porous hydroxyapatite/polymer nanoscale biocomposites for bone regeneration. Acta Biomater. 2011, 7, 3813–3828. [Google Scholar] [CrossRef] [PubMed]
- Salah, H.; Wasim, K.; Harriet, D.; Henry, P.; Michael, M. Calcium Salts Bone Regeneration Scaffolds: A Review Article. Curr. Stem Cell Res. Ther. 2012, 7, 336–346. [Google Scholar]
- Fabritius, H.; Sachs, C.; Raabe, D.; Nikolov, S.; Friák, M.; Neugebauer, J. Chitin in the Exoskeletons of Arthropoda: From Ancient Design to Novel Materials Science. In Chitin: Formation and Diagenesis; Gupta, N.S., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 35–60. [Google Scholar]
- Doherty, M.J.; Schlag, G.; Schwarz, N.; Mollan, R.A.; Nolan, P.C.; Wilson, D.J. Biocompatibility of xenogeneic bone, commercially available coral, a bioceramic and tissue sealant for human osteoblasts. Biomaterials 1994, 15, 601–608. [Google Scholar] [CrossRef]
- Guillemin, G.; Patat, J.L.; Fournie, J.; Chetail, M. The use of coral as a bone graft substitute. J. Biomed. Mater. Res. 2004, 21, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Marchac, D.; Sandor, G. Use of coral granules in the craniofacial skeleton. J. Craniofac. Surg. 1994, 5, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Manjubala, I.; Sivakumar, M.; Nikkath, S.N. Synthesis and characterisation of hydroxy/fluoroapatite solid solution. J. Mater. Sci. 2001, 36, 5481–5486. [Google Scholar] [CrossRef]
- Langer, R. Perspectives and Challenges in Tissue Engineering and Regenerative Medicine. Adv. Mater. 2009, 21, 3235–3236. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Lai, W.-F.; Jung, H.-S. Evolving Marine Biomimetics for Regenerative Dentistry. Mar. Drugs 2014, 12, 2877–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, B.; Lemos, A.; Soundrapandian, C.; Sen, P.S.; Datta, S.; Ferreira, J.M.F.; Basu, D. Development of porous HAp and beta-TCP scaffolds by starch consolidation with foaming method and drug-chitosan bilayered scaffold based drug delivery system. J. Mater. Sci. Mater. Med. 2010, 21, 2955–2969. [Google Scholar] [CrossRef] [PubMed]
- Baradari, H.; Damia, C.; Dutreih-Colas, M.; Champion, E.; Chulia, D.; Viana, M. beta-TCP porous pellets as an orthopaedic drug delivery system: Ibuprofen/carrier physicochemical interactions. Sci. Technol. Adv. Mater. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, H.; Liu, J.; He, H.; Hua, X.; He, Q.; Zhang, L.; Ye, X.; Shi, J. A mesoporous silica nanoparticulate/beta-TCP/BG composite drug delivery system for osteoarticular tuberculosis therapy. Biomaterials 2011, 32, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Seto, J.; Ma, Y.; Davis, S.A.; Meldrum, F.; Gourrier, A.; Kim, Y.Y.; Schilde, U.; Sztucki, M.; Burghammer, M.; Maltsev, S.; et al. Structure-property relationships of a biological mesocrystal in the adult sea urchin spine. Proc. Natl. Acad. Sci. USA 2012, 109, 3699–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, A.; Addadi, L.; Weiner, S. Interactions of sea-urchin skeleton macromolecules with growing calcite crystals—A study of intracrystalline proteins. Nature 1988, 331, 546. [Google Scholar] [CrossRef]
- Cao, L.; Li, X.; Zhou, X.; Li, Y.; Vecchio, K.S.; Yang, L.; Cui, W.; Yang, R.; Zhu, Y.; Guo, Z.; et al. Lightweight Open-Cell Scaffolds from Sea Urchin Spines with Superior Material Properties for Bone Defect Repair. ACS Appl. Mater. Interfaces 2017, 9, 9862–9870. [Google Scholar] [CrossRef] [PubMed]
- Palaveniene, A.; Harkavenko, V.; Kharchenko, V.; Daugela, P.; Pranskunas, M.; Juodzbalys, G.; Babenko, N.; Liesiene, J. Cuttlebone as a Marine-Derived Material for Preparing Bone Grafts. Mar. Biotechnol. 2018, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Ozin, G.A. Synthesis of inorganic materials with complex form. Nature 1996, 382, 313. [Google Scholar] [CrossRef]
- Shapiro, O.H.; Kramarsky-Winter, E.; Gavish, A.R.; Stocker, R.; Vardi, A. A coral-on-a-chip microfluidic platform enabling live-imaging microscopy of reef-building corals. Nat. Commun. 2016, 7, 10860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Hu, H.; Zhao, S.; Liu, Y.-M. Microfluidic Platform with In-Chip Electrophoresis Coupled to Mass Spectrometry for Monitoring Neurochemical Release from Nerve Cells. Anal. Chem. 2016, 88, 5338–5344. [Google Scholar] [CrossRef] [PubMed]
- Campana, V.; Milano, G.; Pagano, E.; Barba, M.; Cicione, C.; Salonna, G.; Lattanzi, W.; Logroscino, G. Bone substitutes in orthopaedic surgery: From basic science to clinical practice. J. Mater. Sci. 2014, 25, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Birchall, J.D.; Thomas, N.L. On the architecture and function of cuttlefish bone. J. Mater. Sci. 1983, 18, 2081–2086. [Google Scholar] [CrossRef]
- Garcia-Enriquez, S.; Guadarrama, H.E.; Reyes-Gonzalez, I.; Mendizabal, E.; Jasso-Gastinel, C.F.; Garcia-Enriquez, B.; Rembao-Bojorquez, D.; Pane-Pianese, C. Mechanical performance and in vivo tests of an acrylic bone cement filled with bioactive sepia officinalis cuttlebone. J. Biomater. Sci. Polym. Ed. 2010, 21, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Bandel, K.; Boletzky, S.V. A comparative study of the structure, development and morphological relationships of chambered cephalopod shells. Veliger 1979, 11, 313–354. [Google Scholar]
- Florek, M.; Fornal, E.; Gómez-Romero, P.; Zieba, E.; Paszkowicz, W.; Lekki, J.; Nowak, J.; Kuczumow, A. Complementary microstructural and chemical analyses of Sepia officinalis endoskeleton. Mater. Sci. Eng. C 2009, 29, 1220–1226. [Google Scholar] [CrossRef]
- Cadman, J.; Zhou, S.; Chen, Y.; Li, W.; Appleyard, R.; Li, Q. Characterization of cuttlebone for a biomimetic design of cellular structures. Acta Mech. Sin. 2010, 26, 27–35. [Google Scholar] [CrossRef]
- Schröder, H.C.; Grebenjuk, V.A.; Wang, X.; Müller, W.E.G. Hierarchical architecture of sponge spicules: Biocatalytic and structure-directing activity of silicatein proteins as model for bioinspired applications. Bioinspir. Biomimet. 2016, 11, 041002. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wiens, M.; Schroder, H.C.; Jochum, K.P.; Schlossmacher, U.; Gotz, H.; Duschner, H.; Muller, W.E. Circumferential spicule growth by pericellular silica deposition in the hexactinellid sponge Monorhaphis chuni. J. Exp. Biol. 2011, 214 Pt 12, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Schroder, H.C.; Boreiko, O.; Krasko, A.; Reiber, A.; Schwertner, H.; Muller, W.E. Mineralization of SaOS-2 cells on enzymatically (silicatein) modified bioactive osteoblast-stimulating surfaces. J. Biomed. Mater. Res. Part B Appl. Biomater. 2005, 75, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.E.; Binder, M.; von Lintig, J.; Guo, Y.W.; Wang, X.; Kaandorp, J.A.; Wiens, M.; Schroder, H.C. Interaction of the retinoic acid signaling pathway with spicule formation in the marine sponge Suberites domuncula through activation of bone morphogenetic protein-1. Biochim. Biophys. Acta 2011, 1810, 1178–1194. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Wang, X.; Cui, F.-Z.; Jochum, K.P.; Tremel, W.; Bill, J.; Schröder, H.C.; Natalio, F.; Schloßmacher, U.; Wiens, M. Sponge spicules as blueprints for the biofabrication of inorganic–organic composites and biomaterials. Appl. Microbiol. Biotechnol. 2009, 83, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledger, P.W.; Jones, W.C. Spicule formation in the calcareous sponge Sycon ciliatum. Cell Tissue Res. 1977, 181, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Schröder, H.C.; Wiens, M.; Ushijima, H.; Müller, W.E.G. Bio-silica and bio-polyphosphate: Applications in biomedicine (bone formation). Curr. Opin. Biotechnol. 2012, 23, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Boute, N.; Exposito, J.-Y.; Boury-Esnault, N.; Vacelet, J.; Noro, N.; Miyazaki, K.; Yoshizato, K.; Garrone, R. Type IV collagen in sponges, the missing link in basement membrane ubiquity. Biol. Cell 1996, 88, 37–44. [Google Scholar] [CrossRef]
- Exposito, J.Y.; Cluzel, C.; Garrone, R.; Lethias, C. Evolution of collagens. Anatom. Record 2002, 268, 302–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swatschek, D.; Schatton, W.; Kellermann, J.; Müller, W.E.G.; Kreuter, J. Marine sponge collagen: Isolation, characterization and effects on the skin parameters surface-pH, moisture and sebum. Eur. J. Pharm. Biopharm. 2002, 53, 107–113. [Google Scholar] [CrossRef]
- Nicklas, M.; Schatton, W.; Heinemann, S.; Hanke, T.; Kreuter, J. Preparation and characterization of marine sponge collagen nanoparticles and employment for the transdermal delivery of 17beta-estradiol-hemihydrate. Drug Dev. Ind. Pharm. 2009, 35, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Weaver, J.C.; Thanawala, M.S.; Sundar, V.C.; Morse, D.E.; Fratzl, P. Skeleton of Euplectella sp.: Structural Hierarchy from the Nanoscale to the Macroscale. Science 2005, 309, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Miserez, A.; Weaver James, C.; Thurner Philipp, J.; Aizenberg, J.; Dauphin, Y.; Fratzl, P.; Morse Daniel, E.; Zok Frank, W. Effects of Laminate Architecture on Fracture Resistance of Sponge Biosilica: Lessons from Nature. Adv. Funct. Mater. 2008, 18, 1241–1248. [Google Scholar] [CrossRef]
- Abramovitch-Gottlib, L.; Geresh, S.; Vago, R. Biofabricated marine hydrozoan: A bioactive crystalline material promoting ossification of mesenchymal stem cells. Tissue Eng. 2006, 12, 729–739. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macha, I.J.; Ben-Nissan, B. Marine Skeletons: Towards Hard Tissue Repair and Regeneration. Mar. Drugs 2018, 16, 225. https://doi.org/10.3390/md16070225
Macha IJ, Ben-Nissan B. Marine Skeletons: Towards Hard Tissue Repair and Regeneration. Marine Drugs. 2018; 16(7):225. https://doi.org/10.3390/md16070225
Chicago/Turabian StyleMacha, Innocent J., and Besim Ben-Nissan. 2018. "Marine Skeletons: Towards Hard Tissue Repair and Regeneration" Marine Drugs 16, no. 7: 225. https://doi.org/10.3390/md16070225
APA StyleMacha, I. J., & Ben-Nissan, B. (2018). Marine Skeletons: Towards Hard Tissue Repair and Regeneration. Marine Drugs, 16(7), 225. https://doi.org/10.3390/md16070225