(+)-Aeroplysinin-1 Modulates the Redox Balance of Endothelial Cells



Abstract
:1. Introduction
2. Results
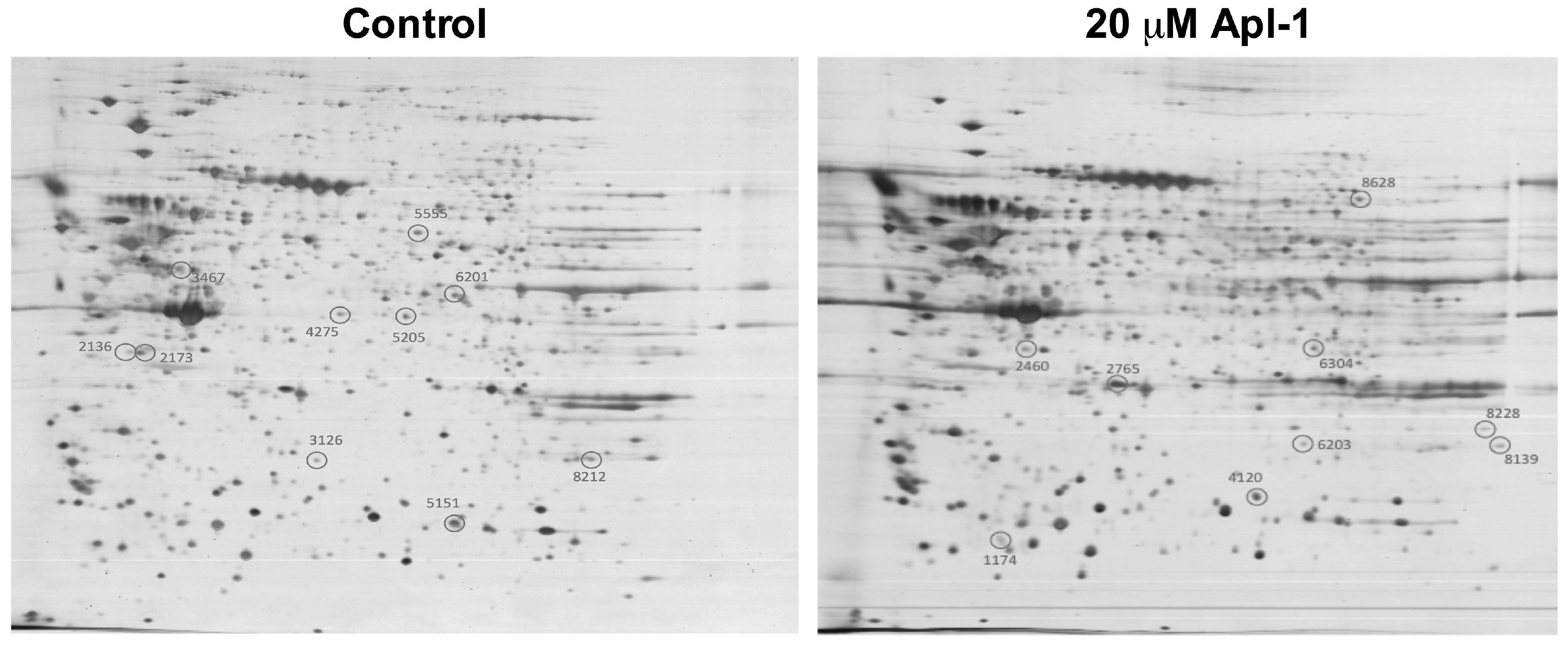
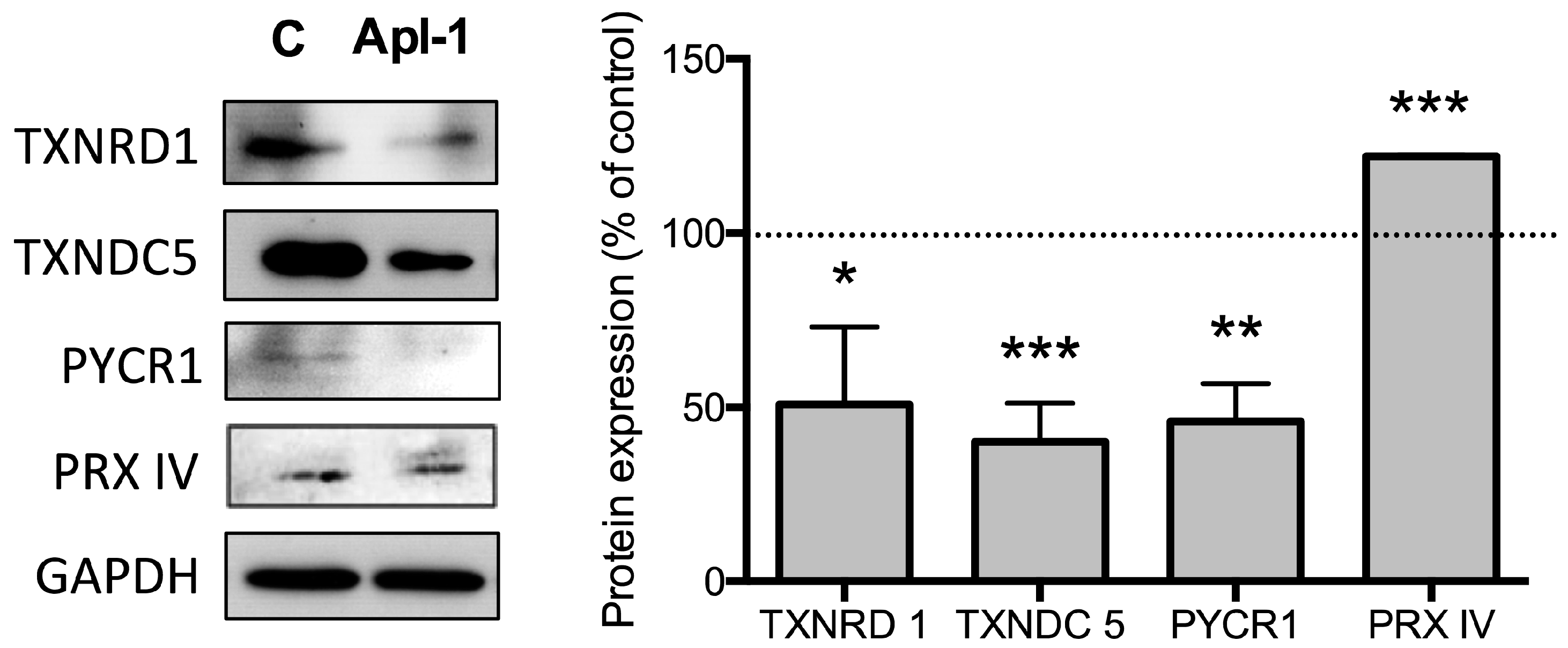
2.1. (+)-Aeroplysinin-1 Affects the Expression Levels of Redox Proteins in RF-24 Endothelial Cells
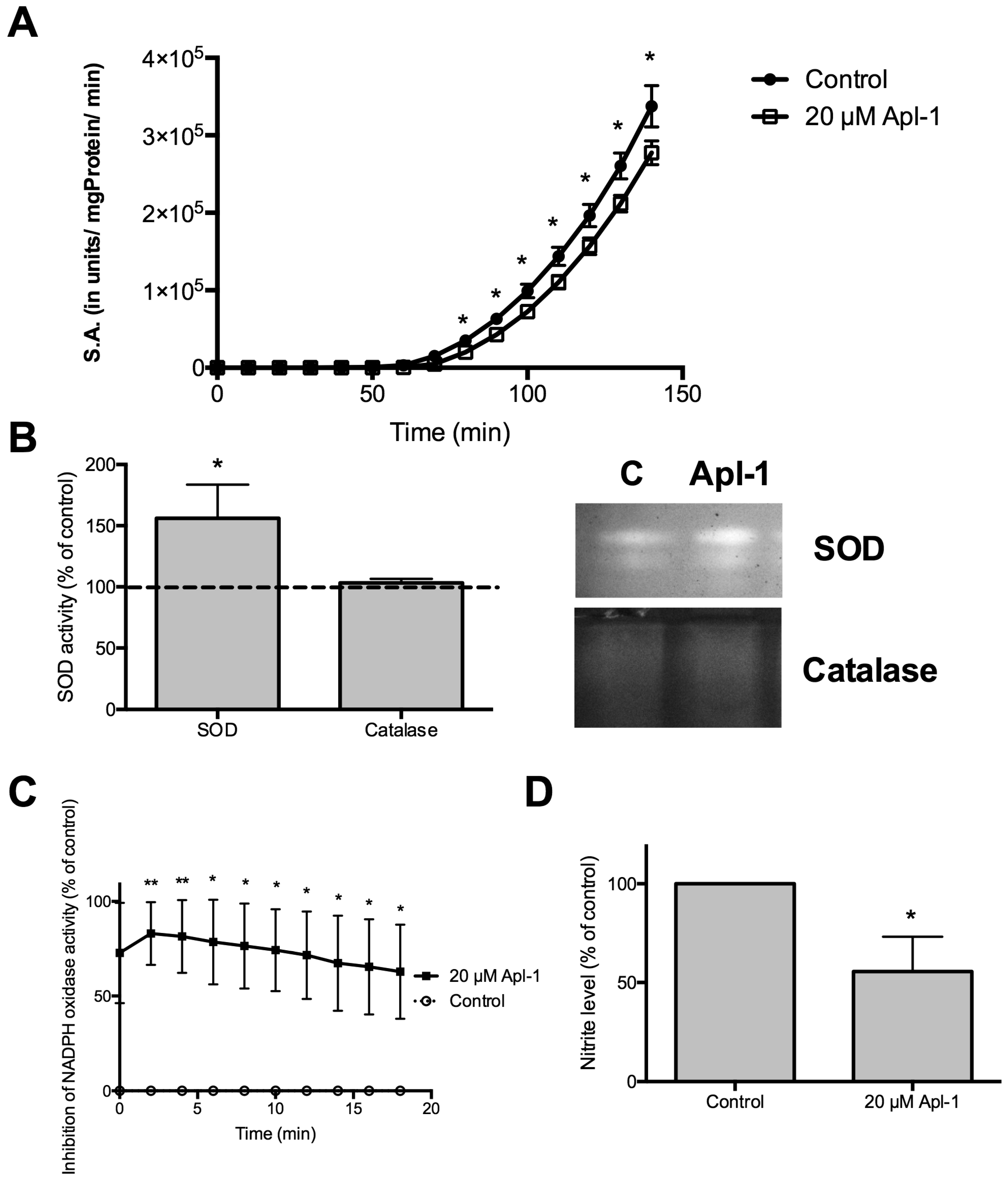
2.2. (+)-Aeroplysinin-1 Affects Redox Enzyme Activities in RF-24 Endothelial Cells
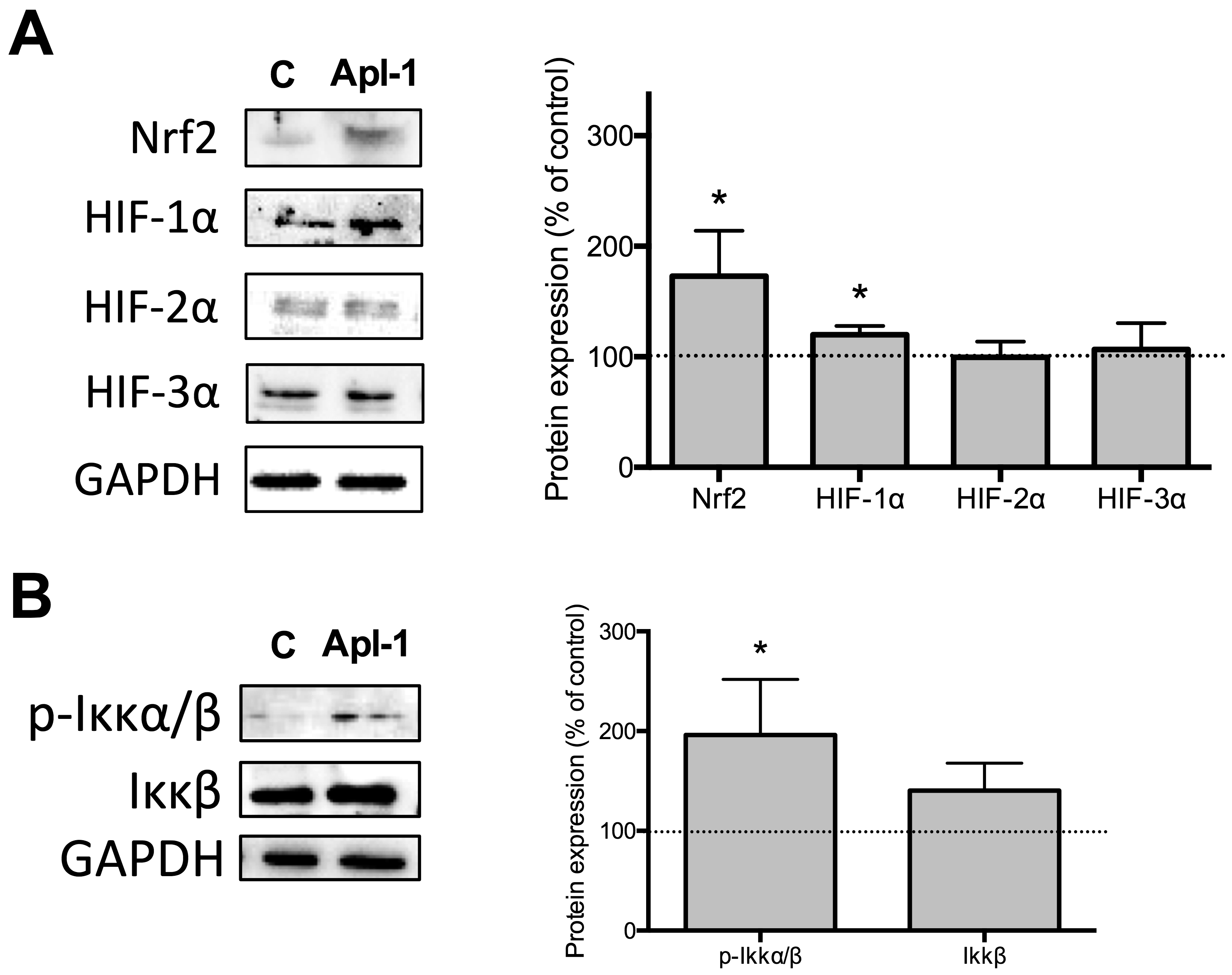
2.3. (+)-Aeroplysinin-1 Modulates Transcription Factors Involved in Redox Homeostasis
2.4. (+)-Aeroplysinin-1 Decreases ROS Levels and Mitochondrial Membrane Potential in RF-24 Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. MTT Cell Survival Assay
4.4. Sample Preparation for Proteomic Analysis, 2-D Electrophoresis and Protein Identification
4.5. Western Blot
4.6. PDI Activity Assay
4.7. SOD and Catalase Zymographies
4.8. Assay of NADPH Oxidase Activity
4.9. Spectrophotometric Measurement of Nitrite as an Indicator of Nitric Oxide Production
4.10. Measurement of Intracellular ROS Levels
4.11. Measurement of Intracellular Mitochondrial Membrane Potential
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- García-Vilas, J.A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.A. Aeroplysinin-1, a sponge-derived multitargeted bioactive marine drug. Mar. Drugs 2016, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Minale, L.S.G. Aeroplysinin-1, an antibacterial bromo-compound from the sponge Verongia aerophoba. J. Chem. Soc. 1972, 16–18. [Google Scholar] [CrossRef]
- Nieto-Ortega, B.; Casado, J.; Blanck, E.W.; López-Navarrete, J.T.; Quesada, A.R.; Ramírez, F.J. Raman optical activity spectra and conformational elucidation of chiral drugs. The case of the antiangiogenic aeroplysinin-1. J. Phys. Chem. 2011, 115, 2752–2755. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.H.; Bernd, A.; Holzmann, H.; Müller-Klieser, W.; Maldhof, A.; Weissmann, N.; Kjajic, Z.; Batel, R.; Schröder, H.C.; Müller, W.E.G. Cytostatic activity of aeroplysinin-1 against lymphoma and epithelioma cells. Z. Naturforsch. C 1989, 44, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.H.; Bernd, A.; Holzmann, H.; Müller-Klieser, W.; Maldhof, A.; Müller, W.E.; Schöder, H.C. Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp. Biochem. Physiol. B 1990, 97, 151–158. [Google Scholar] [CrossRef]
- Rodríguez-Nieto, S.; González-Iriarte, M.; Carmona, R.; Muñoz-Chápuli, R.; Medina, M.A.; Quesada, A.R. Antiangiogenic activity of aeroplysinin-1, a brominated compound isolated from a marine sponge. FASEB J. 2002, 16, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; Rodríguez-Nieto, S.; García-Caballero, M.; Medina, M.A.; Quesada, A.R. The antiangiogenic compound aeroplysinin-1 induces apoptosis in endothelial cells by activating the mitochondrial pathway. Mar. Drugs 2012, 10, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; García-Vilas, J.A.; Cárdenas, C.; Melgarejo, E.; Quesada, A.R.; Medina, M.A. The brominated compound aeroplysinin-1 inhibits proliferation and the expression of key pro- inflammatory molecules in human endothelial and monocyte cells. PLoS ONE 2013, 8, e55203. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; Quesada, A.R.; Medina, M.A. (+)-Aeroplysinin-1 inhibits Akt and Erk pathway signaling selectively in endothelial cells. Integr. Cancer Sci. Ther. 2015, 2, 181–184. [Google Scholar] [CrossRef]
- Brakenhielm, E.; Cao, R.; Cao, Y. Suppression of angiogenesis, tumor growth, and wound healing by resveratrol, a natural compound in red wine and grapes. FASEB J. 2001, 15, 1798–1800. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Fukui, M.; Zhu, B.T. Protein disulfide isomerase mediates glutathione depletion-induced cytotoxicity. Biochem. Biophys. Res. Commun. 2016, 477, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. Biotechniques 2011, 50, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Szabadkai, G.; Duchen, M.R. Mitochondria: The hub of cellular Ca2+ signaling. Physiology 2008, 23, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yang, K.; Wang, X.; Wang, X.; Wang, C.C. A novel reaction of peroxiredoxin 4 towards substrates in oxidative protein folding. PLoS ONE 2014, 9, e105529. [Google Scholar] [CrossRef] [PubMed]
- Okado-Matsumoto, A.; Matsumoto, A.; Fujii, J.; Taniguchi, N. Peroxiredoxin IV is a secretable protein with heparin-binding properties under reduced conditions. J. Biochem. 2000, 127, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Horna-Terrón, E.; Pradilla-Dieste, A.; Sánchez-de-Diego, C.; Osada, J. TXNDC5, a newly discovered disulfide isomerase with a key role in cell physiology and pathology. Int. J. Mol. Sci. 2014, 15, 23501–23518. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, C.; Franckenstein, D.; Schmidt, M.; Gehrmann, M.; Hermes, M.; Geppert, W.S.B.; Maccoux, L.J.; Schug, M.; Schumann, A.; Wilhelm, C.; et al. Role of thioredoxin reductase 1 and thioredoxin interacting protein in prognosis of breast cancer. Breast Cancer Res. 2010, 12, R44. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.L.; Lee, M.B.; Tang, M.; den Besten, W.; Hu, S.; Sweredoski, M.J.; Hess, S.; Chou, C.M.; Changou, C.A.; Su, M.; et al. PYCR1 and PYCR2 interact and collaborate with RRM2b to protect cells from overt oxidative stress. Sci. Rep. 2016, 15, 18846. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.A.; Khalil, S.; Zhaorigetu, S.; Liu, Z.; Tyler, M.; Wan, G.; Valle, D. Human Δ1-pyrroline-5-carboxylate synthase: Function and regulation. Amino Acids 2008, 35, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed]
- Nayernia, Z.; Jaquet, V.; Krause, K.H. New insights on NOX enzymes in the central nervous system. Antioxid. Redox Signal. 2014, 20, 2815–2837. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar] [PubMed]
- Stamler, J.S.; Singel, D.J.; Loscalzo, J. Biochemistry of nitric oxide and its redox-activated forms. Science 1992, 258, 1898–1902. [Google Scholar] [CrossRef] [PubMed]
- Stamler, J.S. Redox signalling, nitrosylation and related target interactions of nitric oxide. Cell 1994, 78, 931–936. [Google Scholar] [CrossRef]
- Talks, K.L.; Turley, H.; Gatter, K.C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Harris, A.L. The expression and distribution of the hypoxia-inducible factors HIF-1α and HIF-2α in normal human tissues, cancers, and tumor-associated macrophages. Am. J. Pathol. 2000, 157, 411–421. [Google Scholar] [CrossRef]
- Duan, C. Hypoxia-inducible factor 3 biology: Complexities and emerging themes. Am. J. Physiol. Cell Physiol. 2016, 310, C260–C269. [Google Scholar] [CrossRef] [PubMed]
- Branco-Price, C.; Zhang, N.; Schnelle, M.; Evans, C.; Katschinski, D.M.; Liao, D.; Ellies, L.; Johnson, R.S. Endothelial cell HIF-1α and HIF-2α differentially regulate metastatic success. Cancer Cell 2012, 21, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Xia, Y.; Li, J.; Deng, L.; Zhao, L.; Shi, J.; Wang, X.; Sun, B. High expression levels of IKKα and IKKβ are necessary for the malignant properties of liver cancer. Int. J. Cancer 2010, 126, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Page, A.; Cascallana, J.L.; Casanova, M.L.; Navarro, M.; Alameda, J.P.; Pérez, P.; Bravo, A.; Ramírez, A. IKKβ overexpression leads to pathologic lesions in stratified epithelia and exocrine glands and to tumoral transformation of oral epithelia. Mol. Cancer Res. 2011, 9, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; Halliwell, B. Damage to DNA by reactive oxygen and nitrogen species: Role in inflammatory disease and progression to cancer. Biochem. J. 1996, 313, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.K.; Davies, K.J.; Forman, H.J. Reactive oxygen and nitrogen species in neurodegeneration. Free Rad. Biol. Med. 2013, 62, 1–3. [Google Scholar] [CrossRef] [PubMed]
- El-Azab, M.F.; Hazem, R.M.; Moustafa, Y.M. Role of simvastatin and/or antioxidant vitamins in therapeutic angiogenesis in experimental diabetic hindlimb ischemia: Effects on capillary density, angiogenesis markers, and oxidative stress. Eur. J. Pharmacol. 2012, 690, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Fuchs-Tarlovsky, V. Role of antioxidants in cancer therapy. Nutrition 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Casserly, I.; Topol, E.J. Convergence of atherosclerosis and Alzheimer’s disease: Inflammation, cholesterol, and misfolded proteins. Lancet 2004, 363, 1139–1146. [Google Scholar] [CrossRef]
- Iannitti, T.; Palmieri, B. Antioxidant therapy effectiveness: An up to date. Eur. Rev. Med. Pharmacol. Sci. 2009, 13, 245–278. [Google Scholar] [PubMed]
- Yang, G.Z.; Wang, Z.J.; Bai, F.; Qin, X.J.; Cao, J.; Lv, J.Y.; Zhang, M.S. Epigallocatechin-3-gallate protects HUVECs from PM2.5-induced oxidative stress injury by activating critical antioxidant pathways. Molecules 2015, 20, 6626–6639. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Hu, X.; Hu, G.; Xu, C.; Jiang, H. Curcumin attenuates hydrogen peroxide-induced premature senescence via the activation of SIRT1 in Human umbilical vein endothelial cells. Biol. Pharm. Bull. 2015, 38, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, H.; Starkov, A.A.; Manfredi, G.; Chinopoulos, C. A kinetic assay of mitochondrial ADP-ATP exchange rate in permeabilized cells. Anal. Biochem. 2010, 407, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Chinopoulos, C.; Tretter, L.; Adam-Vizi, V. Depolarization of in situ mitochondria due to hydrogene peroxide-induced oxidative stress in nerve terminals: Inhibition of alpha-ketoglutarate dehydrogenase. J. Neurochem. 1999, 73, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Heerdt, B.G.; Houston, M.A.; Wilson, A.J.; Augenlicht, L.H. The intrinsic mitochondrial membrane potential (Deltapsim) is associated with steady-state mitochondrial activity and the extent to which colonic epithelial cells under butyrate-mediated growth arrest and apoptosis. Cancer Res. 2003, 63, 6311–6319. [Google Scholar] [PubMed]
- Kluge, M.A.; Fetterman, J.L.; Vita, J.A. Mitochondria and endothelial function. Circ. Res. 2013, 112, 1171–1188. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Guh, J.H.; Chueh, S.C.; Yu, H.J. Anti-angiogenic effects and mechanism of prazosin. Prostate 2011, 71, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Galougahi, K.K.; Ashley, E.A.; Ali, Z.A. Redox regulation of vascular remodeling. Cell. Mol. Life Sci. 2016, 76, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Van Beijnum, J.R.; van der Linden, E.; Griffioen, A.W. Angiogenic profiling and comparison of immortalized endothelial cells for functional genomics. Exp. Cell Res. 2008, 314, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Gel | Protein | Gen | Function |
|---|---|---|---|
| 2136 | Ubiquitin-conjugating enzyme E2 1 | UBC1 | Catalyzes the covalent attachment of ubiquitin to other proteins. Functions in degradation of misfolded or regulated proteins localized in the endoplasmic reticulum (ER) lumen or membrane via the ubiquitin-proteasome system. |
| 2173 | Lactoylglutathione lyase | GLO1 | Catalyzes the conversion of hemimercaptal, formed from methylglyoxal and glutathione, to S-lactoylglutathione. |
| 3126 | Chloride intracellular channel protein 4 | CLIC4 | Can insert into membranes and form poorly selective ion channels that may also transport chloride ions. Has alternate cellular functions like a potential role in angiogenesis or in maintaining apical-basolateral membrane polarity during mitosis and cytokinesis. Could also promote endothelial cell proliferation and regulate endothelial morphogenesis (tubulogenesis). |
| 3467 | TXNDC5 protein | TXNDC5 | Cell redox homeostasis. TXNCD5 is a protein-disulfide isomerase. Its expression is induced by hypoxia and its role may be to protect hypoxic cells from apoptosis. |
| 4275 | L-lactate dehydrogenase B chain | LDHB | Catalytic activity (S)-lactate + NAD+ = pyruvate + NADH |
| 5151 | Triosephosphate isomerase | TPI1 | d-glyceraldehyde 3-phosphate = glycerone phosphate. |
| 5205 | Transaldolase | TALDO1 | Transaldolase is important for the balance of metabolites in the pentose-phosphate pathway. Sedoheptulose 7-phosphate + d-glyceraldehyde 3-phosphate = d-erythrose 4-phosphate + d-fructose 6-phosphate. |
| 5555 | Thioredoxin reductase 1, cytoplasmic | TXNRD1 | Isoform 1 may possess glutaredoxin activity as well as thioredoxin reductase activity and induces actin and tubulin polymerization, leading to formation of cell membrane protrusions. Isoform enhances the transcriptional activity of estrogen receptors alpha and beta while isoform enhances the transcriptional activity of the beta receptor only. |
| 6201 | Annexin A1 | ANXA1 | Calcium/phospholipid-binding protein which promotes membrane fusion and is involved in exocytosis. This protein regulates phospholipase A2 activity. |
| 8212 | Pyrroline-5-carboxylate reductase 1, mitocondrial | PYCR1 | Housekeeping enzyme that catalyzes the last step in proline biosynthesis. Can utilize both NAD and NADP, but has higher affinity for NAD. Involved in the cellular response to oxidative stress. l-proline + NAD(P)+ = 1-pyrroline-5-carboxylate + NAD(P)H. |
| ID Gel | Protein | Gen | Function |
|---|---|---|---|
| 1174 | Vimentin | VIM | Vimentins are class-III intermediate filaments found in various non-epithelial cells, especially mesenchymal cells. |
| 2460 | Keratin, type II cytoskeletal 7 | KRT7 SCL | Blocks interferon-dependent interphase and stimulates DNA synthesis in cells. Involved in the translational regulation of the human papillomavirus type 16 E7 mRNA (HPV16 E7). |
| 2765 | Heat shock protein HSP 90-beta | HSP90ß | Molecular chaperone that promotes the maturation, structural maintenance, and proper regulation of specific target proteins involved for instance in cell cycle control and signal transduction. |
| 4120 | Peroxiredoxin-4 | PRDX4 | Probably involved in redox regulation of the cell. Regulates the activation of NF-kappa-B in the cytosol by a modulation of I-kappa-B-alpha phosphorylation. |
| 6203 | Estrogen sulfotransferase | SULT1E1 STE | May control the level of the estrogen receptor by sulfurylating free estradiol. 3′-phosphoadenylyl sulfate + estrone = adenosine 3′,5′-bisphosphate + estrone 3-sulfate. |
| 6304 | GDP-L-fucose synthase | TSTA3 SDR4E1 | GDP-l-fucose + NADP+ = GDP-4-dehydro-6-deoxy-d-mannose + NADPH. |
| 8139 | Guanine nucleotide-binding protein subunit beta-2-like 1 | GNB2L1 HLC7 PIG21 | Involved in PKC-dependent translocation of ADAM12 to the cell membrane. Promotes the ubiquitination and proteasome-mediated degradation of proteins such as CLEC1B and HIF1A. |
| 8228 | Voltage-dependent anion-selective channel protein 2 | VDAC2 | Forms a channel through the mitochondrial outer membrane that allows diffusion of small hydrophilic molecules. The channel adopts an open conformation at low or zero membrane potential and a closed conformation at potentials above 30–40 mV. The open state has a weak anion selectivity whereas the closed state is cation-selective. |
| 8628 | Transketolase | TKT | Sedoheptulose 7-phosphate + d-glyceraldehyde 3-phosphate = d-ribose 5-phosphate + d-xylulose 5-phosphate. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Vilas, J.A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.Á. (+)-Aeroplysinin-1 Modulates the Redox Balance of Endothelial Cells. Mar. Drugs 2018, 16, 316. https://doi.org/10.3390/md16090316
García-Vilas JA, Martínez-Poveda B, Quesada AR, Medina MÁ. (+)-Aeroplysinin-1 Modulates the Redox Balance of Endothelial Cells. Marine Drugs. 2018; 16(9):316. https://doi.org/10.3390/md16090316
Chicago/Turabian StyleGarcía-Vilas, Javier A., Beatriz Martínez-Poveda, Ana R. Quesada, and Miguel Ángel Medina. 2018. "(+)-Aeroplysinin-1 Modulates the Redox Balance of Endothelial Cells" Marine Drugs 16, no. 9: 316. https://doi.org/10.3390/md16090316
APA StyleGarcía-Vilas, J. A., Martínez-Poveda, B., Quesada, A. R., & Medina, M. Á. (2018). (+)-Aeroplysinin-1 Modulates the Redox Balance of Endothelial Cells. Marine Drugs, 16(9), 316. https://doi.org/10.3390/md16090316