Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation

 , ,
, ,
Abstract
:1. Introduction
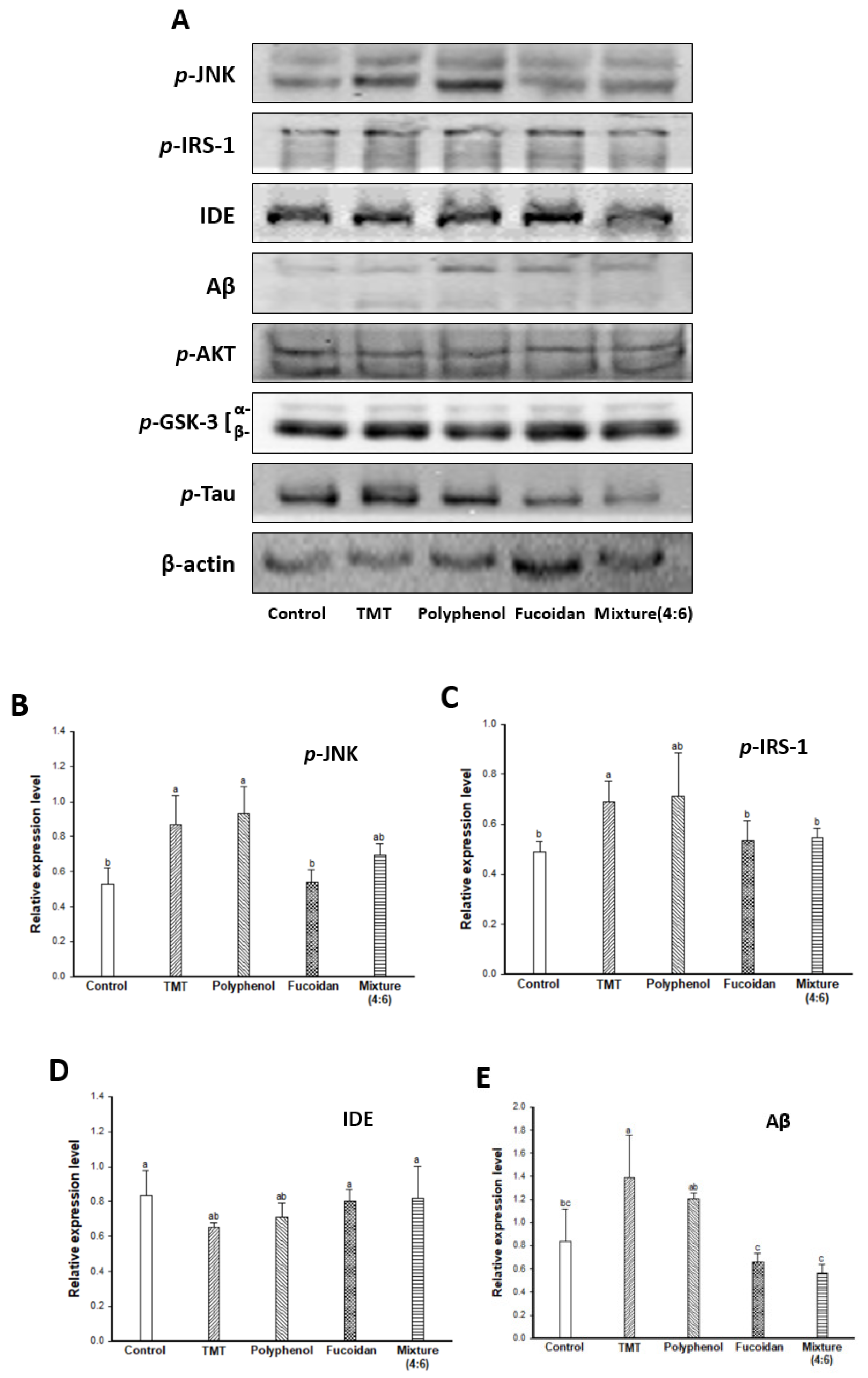
2. Results and Discussion
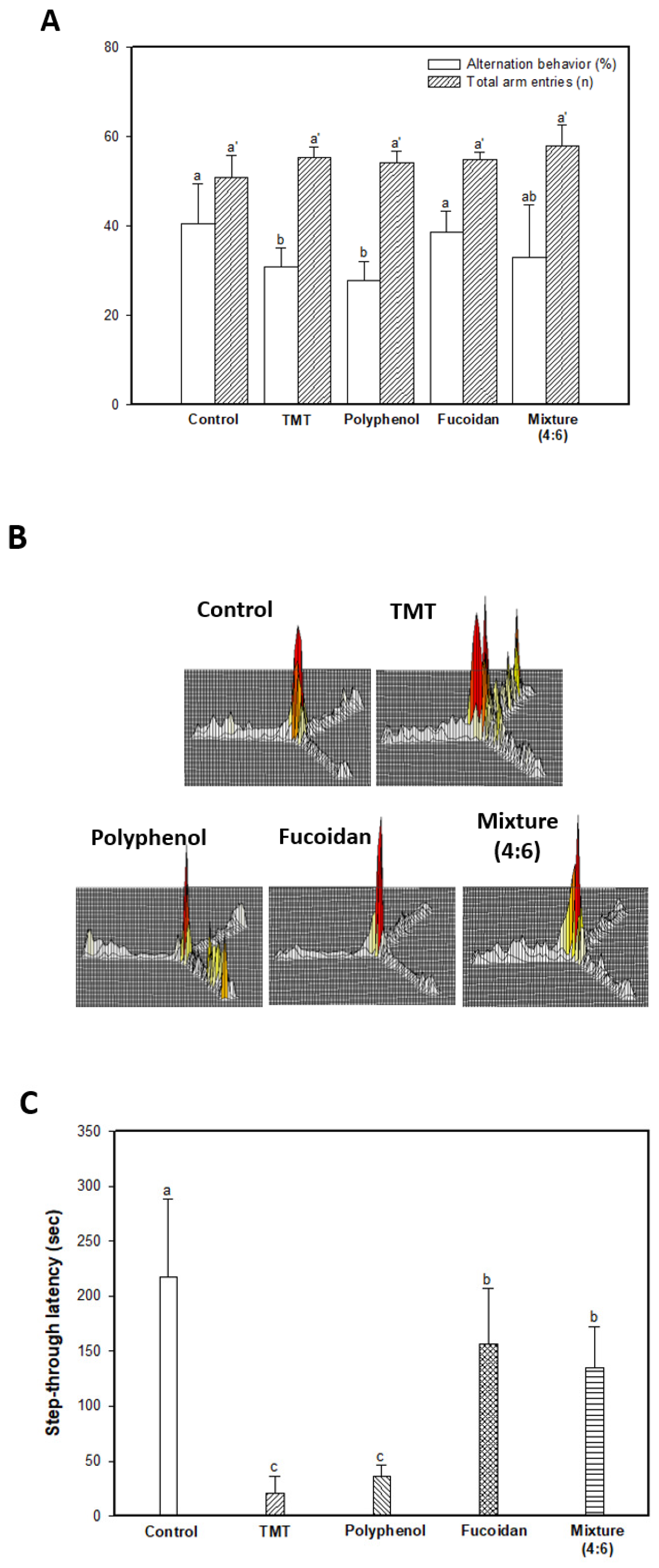
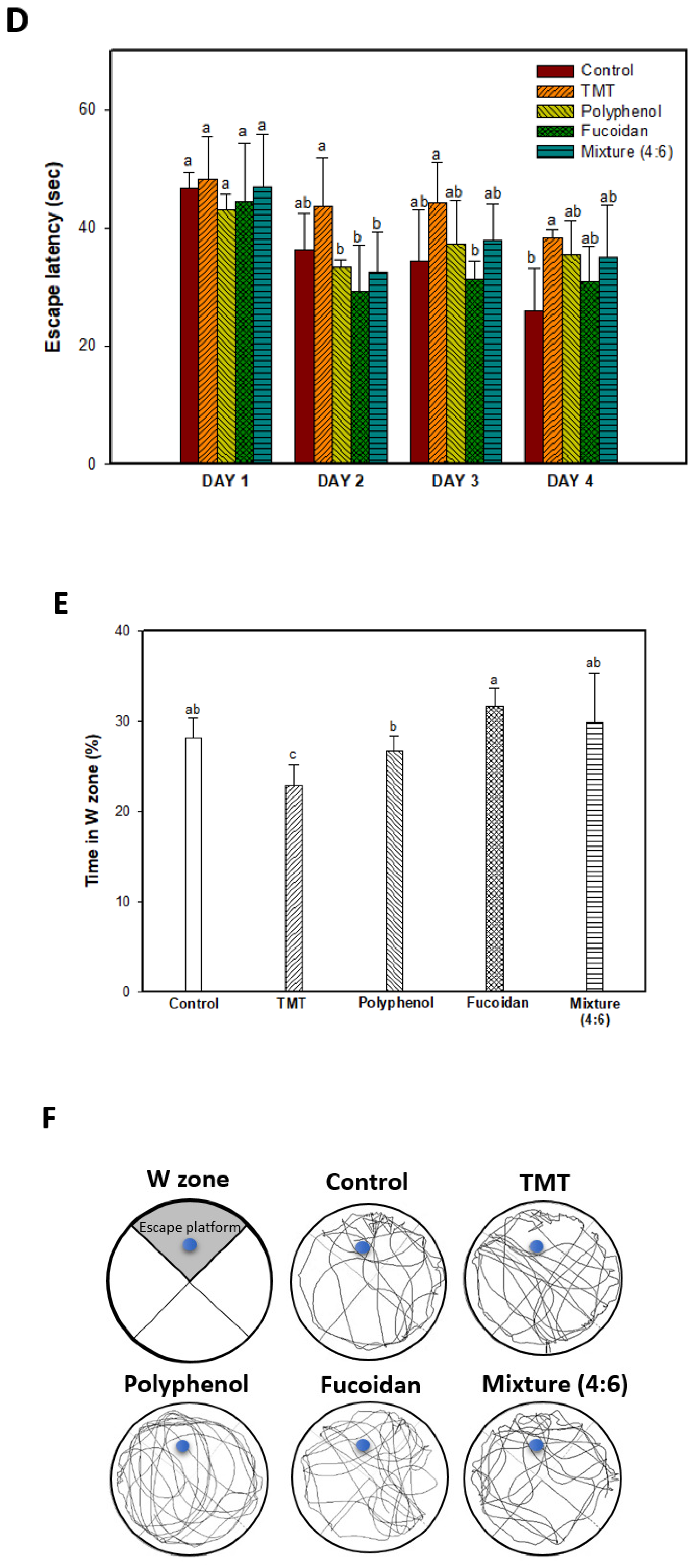
2.1. Behavioral Tests
2.2. Inhibition of Lipid Peroxidation in Mouse Brain Tissue
2.3. Cholinergic System Activity
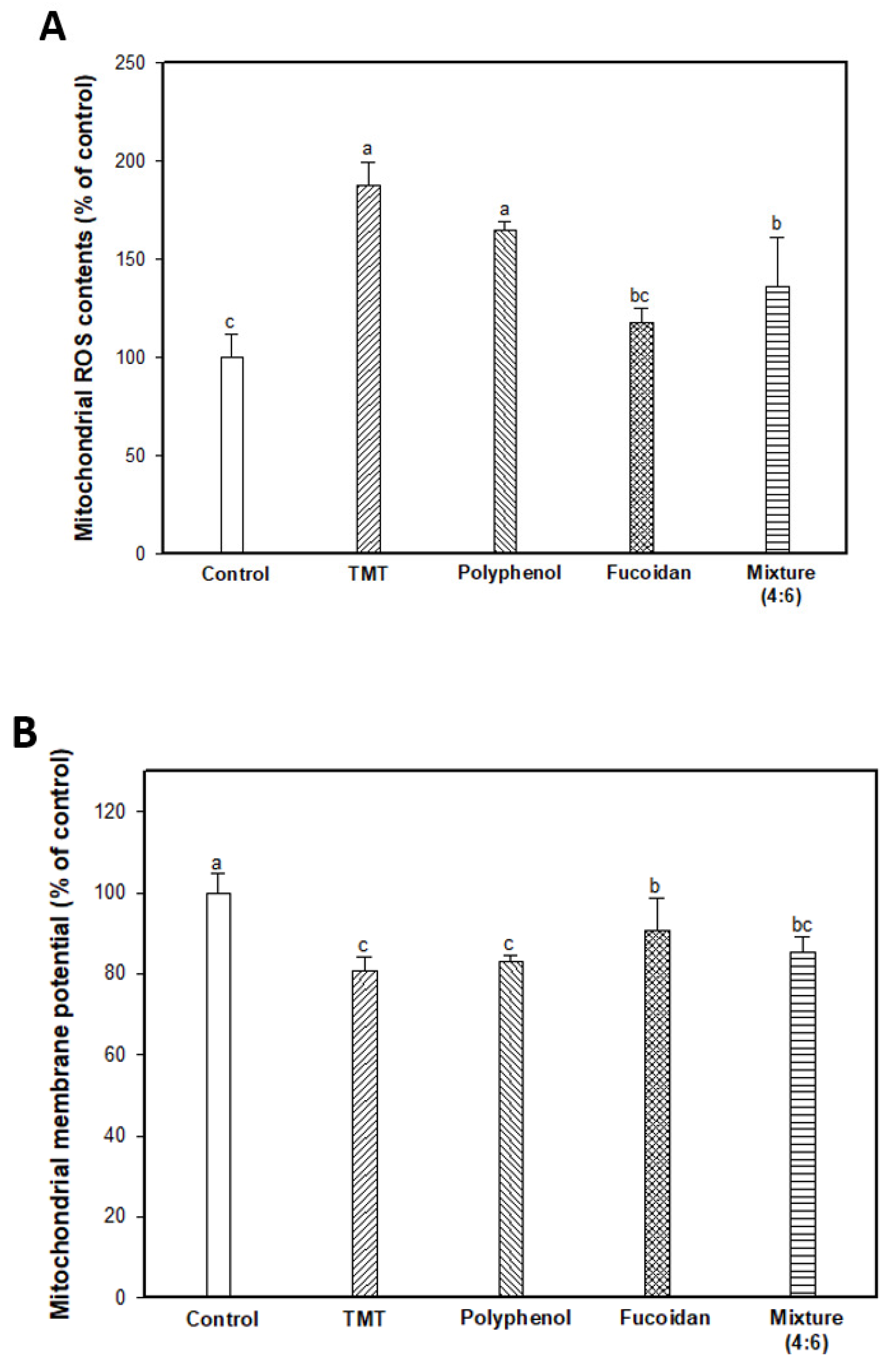
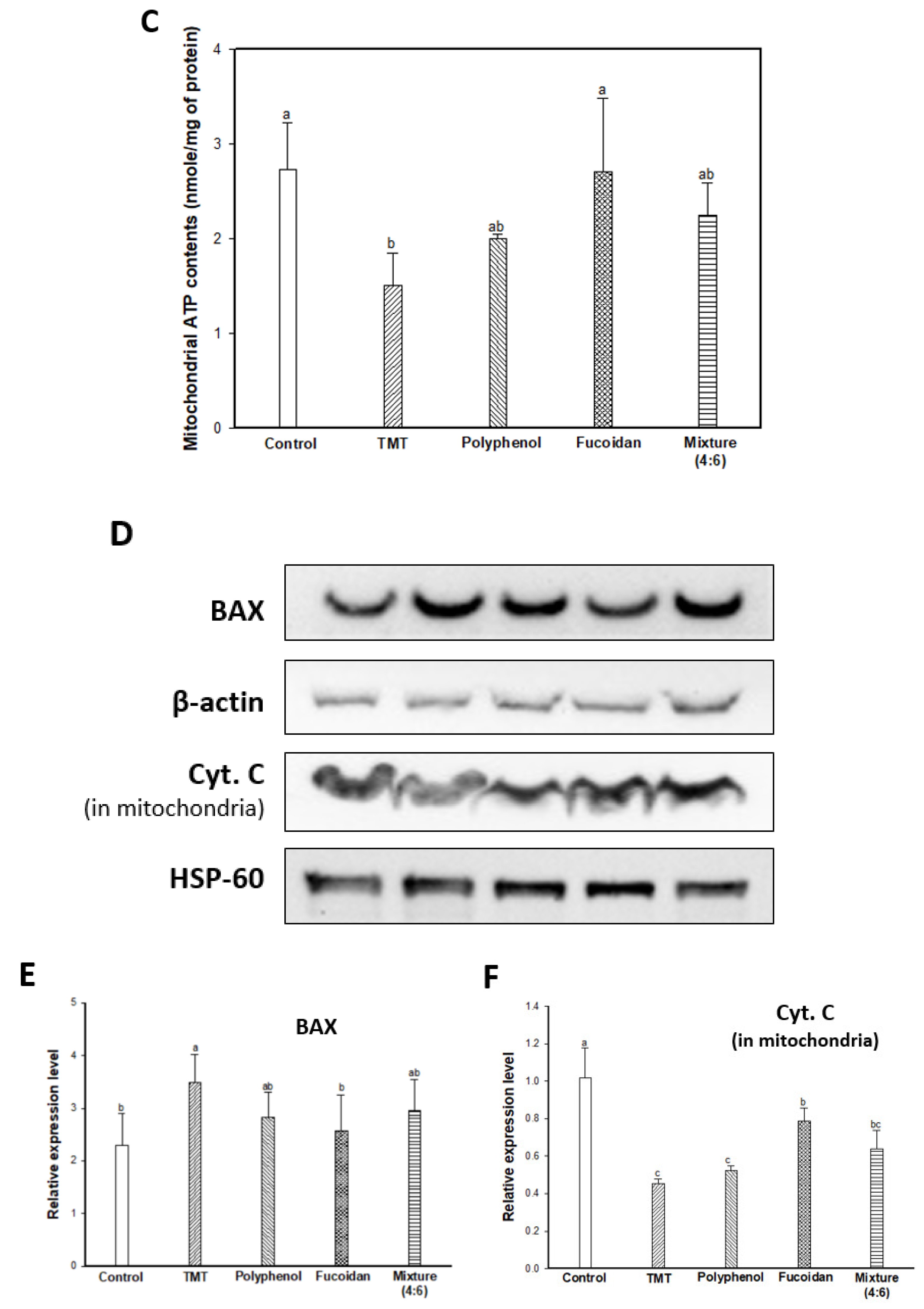
2.4. Mitochondrial Activity
2.5. Cognitive Function-Related Mechanism
2.6. Chemical Composition and Molecular Weight of Fucoidan from E. cava
3. Materials and Methods
3.1. Chemicals
3.2. Sample Preparation
3.2.1. Preparation of Polyphenol Extract from E. cava
3.2.2. Preparation of Fucoidan Extract from E. cava
3.2.3. Preparation of Mixture (Polyphenol:Fucoidan) from E. cava
3.3. In Vivo Experimental Design
3.3.1. Animals
3.3.2. Y-Maze Test
3.3.3. Passive Avoidance Test
3.3.4. Morris Water Maze Test
3.4. Inhibition of Lipid Peroxidation
3.5. Cholinergic System Activity
3.5.1. AChE Activity
3.5.2. ACh Content
3.6. Mitochondrial Activity
3.6.1. Isolation of Mitochondria from Brain Tissue
3.6.2. Mitochondrial ROS Content
3.6.3. Measurement of MMP
3.6.4. ATP Level
3.7. Western Blot Assay for Protein Expression
3.8. Molecular Weight and Composition Analysis of Fucoidan from E. cava
3.8.1. Determination of Sulfate and Monosaccharide Composition
3.8.2. Determination of Average Molecular Weight
3.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pangestuti, R.; Kim, S.K. Neuroprotective Effects of Marine Algae. Mar. Drugs 2011, 9, 803–818. [Google Scholar] [CrossRef] [PubMed]
- LaFerla, F.M.; Oddo, S. Alzheimer’s Disease: Aβ, Tau and Synaptic Dysfunction. Trends Mol. Med. 2005, 11, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Shinall, H.; Song, E.S.; Hersh, L.B. Susceptibility of Amyloid Beta Peptide Degrading Enzymes to Oxidative Damage: A Potential Alzheimer’s Disease Spiral. Biochemistry 2005, 44, 15345–15350. [Google Scholar] [CrossRef] [PubMed]
- Bedse, G.; Di Domenico, F.; Serviddio, G.; Cassano, T. Aberrant Insulin Signaling in Alzheimer’s Disease: Current Knowledge. Front. Neurosci. 2015, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Geloso, M.C.; Vercelli, A.; Corvino, V.; Repici, M.; Boca, M.; Haglid, K.; Zelano, G.; Michetti, F. Cyclooxygenase-2 and Caspase 3 Expression in Trimethyltin-induced Apoptosis in the Mouse Hippocampus. Exp. Neurol. 2002, 175, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Geloso, M.C.; Corvino, V.; Michetti, F. Trimethyltin-induced Hippocampal Degeneration as a Tool to Investigate Neurodegenerative Processes. Neurochem. Int. 2011, 58, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Lee, M.; Shim, H.S.; Lee, G.; Chung, S.Y.; Kang, Y.M.; Lee, B.; Seo, Y.B.; Kim, K.S.; Shim, I. Fermented Saccharina Japonica (Phaeophyta) Improves Neuritogenic Activity and TMT-induced Cognitive Deficits in Rats. Algae 2016, 31, 73–84. [Google Scholar] [CrossRef]
- Raja, A.; Vipin, C.; Aiyappan, A. Biological Importance of Marine Algae—An Overview. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 222–227. [Google Scholar]
- Mohamed, S.; Hashim, S.N.; Rahman, H.A. Seaweeds: A Sustainable Functional Food for Complementary and Alternative Therapy. Trends Food Sci. Technol. 2012, 23, 83–96. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Jeon, Y. Biological Activities and Potential Industrial Applications of Fucose Rich Sulfated Polysaccharides and Fucoidans Isolated from Brown Seaweeds: A Review. Carbohydr. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S. Phlorotannins from Ecklonia Cava (Phaeophyceae): Biological Activities and Potential Health Benefits. Biofactors 2010, 36, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ko, C.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.; Jeon, Y. Anti-inflammatory Effect of Fucoidan Extracted from Ecklonia Cava in Zebrafish Model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Athukorala, Y.; Jung, W.; Vasanthan, T.; Jeon, Y. An Anticoagulative Polysaccharide from an Enzymatic Hydrolysate of Ecklonia Cava. Carbohydr. Polym. 2006, 66, 184–191. [Google Scholar] [CrossRef]
- Ogita, K.; Nishiyama, N.; Sugiyama, C.; Higuchi, K.; Yoneyama, M.; Yoneda, Y. Regeneration of Granule Neurons After Lesioning of Hippocampal Dentate Gyrus: Evaluation using Adult Mice Treated with Trimethyltin Chloride as a Model. J. Neurosci. Res. 2005, 82, 609–621. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.; Shim, I.; Lee, H.; Hahm, D.H. Phellodendron amurense and Its Major Alkaloid Compound, Berberine Ameliorates Scopolamine-Induced Neuronal Impairment and Memory Dysfunction in Rats. Korean J. Physiol. Pharmacol. 2012, 16, 79–89. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide Metabolism in Mammalian Organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid Med. Cell. Longev. 2014. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, N.; Zhou, G.; Lu, X.; Xu, Z.; Li, Z. In Vivo Antioxidant Activity of Polysaccharide Fraction from Porphyra Haitanesis (Rhodephyta) in Aging Mice. Pharmacol. Res. 2003, 48, 151–155. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Zhang, Q.; Zhao, T.; Chen, R.; Zhang, H.; Niu, X.; Li, Z. Antioxidant Activity of Different Sulfate Content Derivatives of Polysaccharide Extracted from Ulva Pertusa (Chlorophyta) in Vitro. Int. J. Biol. Macromol. 2005, 37, 195–199. [Google Scholar] [CrossRef]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal Forebrain Cholinergic Circuits and Signaling in Cognition and Cognitive Decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szutowicz, A.; Bielarczyk, H.; Jankowska-Kulawy, A.; Pawelczyk, T.; Ronowska, A. Acetyl-CoA the Key Factor for Survival or Death of Cholinergic Neurons in Course of Neurodegenerative Diseases. Neurochem. Res. 2013, 38, 1523–1542. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, M.L.; Baisden, R.H. Exposure to Trimethyltin significantly Enhances Acetylcholinesterase Staining in the Rat Dentate Gyrus. Neurotoxicol. Teratol. 1990, 12, 33–39. [Google Scholar] [CrossRef]
- Gao, Y.; Li, C.; Yin, J.; Shen, J.; Wang, H.; Wu, Y.; Jin, H. Fucoidan, a Sulfated Polysaccharide from Brown Algae, Improves Cognitive Impairment Induced by Infusion of Aβ Peptide in Rats. Environ. Toxicol. Pharmacol. 2012, 33, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, S.; Prakash, P.; Essa, M.M.; Qoronfleh, W.M.; Akbar, M.; Song, B.J.; Kumar, S.; Elumalai, P. Marine Derived Bioactive Compounds for Treatment of Alzheimer’s Disease. Front. Biosci. 2018, 10, 537–548. [Google Scholar]
- Yoon, N.Y.; Chung, H.Y.; Kim, H.R.; Choi, J.E. Acetyl-and Butyrylcholinesterase Inhibitory Activities of Sterols and Phlorotannins from Ecklonia Stolonifera. Fish. Sci. 2008, 74, 200–207. [Google Scholar] [CrossRef]
- Giacobini, E. Cholinesterase Inhibitors: New Roles and Therapeutic Alternatives. Pharmacol. Res. 2004, 50, 433–440. [Google Scholar] [CrossRef]
- Davidson, C.E.; Reese, B.E.; Billingsley, M.L.; Yun, J.K. Stannin, a Protein that Localizes to the Mitochondria and Sensitizes NIH-3T3 Cells to Trimethyltin and Dimethyltin Toxicity. Mol. Pharmacol. 2004, 66, 855–863. [Google Scholar] [CrossRef]
- Billingsley, M.L.; Yun, J.; Reese, B.E.; Davidson, C.E.; Buck-Koehntop, B.A.; Veglia, G. Functional and Structural Properties of Stannin: Roles in Cellular Growth, Selective Toxicity, and Mitochondrial Responses to Injury. J. Cell. Biochem. 2006, 98, 243–250. [Google Scholar] [CrossRef]
- Hroudová, J.; Singh, N.; Fišar, Z.; Ghosh, K.K. Progress in Drug Development for Alzheimer’s Disease: An Overview in Relation to Mitochondrial Energy Metabolism. Eur. J. Med. Chem. 2016, 121, 774–784. [Google Scholar] [CrossRef]
- Gottlieb, E.; Vander Heiden, M.G.; Thompson, C.B. Bcl-X(L) Prevents the Initial Decrease in Mitochondrial Membrane Potential and Subsequent Reactive Oxygen Species Production during Tumor Necrosis Factor Alpha-induced Apoptosis. Mol. Cell. Biol. 2000, 20, 5680–5689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, J.; Nussinov, R.; Ma, B. Release of Cytochrome C from Bax Pores at the Mitochondrial Membrane. Sci. Rep. 2017, 7, 2635. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Reed, J.C. Mitochondria and Apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yankner, B.A. Apoptosis in the Nervous System. Nature 2000, 407, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Mullins, R.J.; Diehl, T.C.; Chia, C.W.; Kapogiannis, D. Insulin Resistance as a Link between Amyloid-Beta and Tau Pathologies in Alzheimer’s Disease. Front. Aging Neurosci. 2017, 9, 118. [Google Scholar] [CrossRef]
- Aguirre, V.; Uchida, T.; Yenush, L.; Davis, R.; White, M.F. The C-Jun NH2-terminal Kinase Promotes Insulin Resistance during Association with Insulin Receptor Substrate-1 and Phosphorylation of Ser307. J. Biol. Chem. 2000, 275, 9047–9054. [Google Scholar] [CrossRef]
- Garrido, J.J.; Simon, D.; Varea, O.; Wandosell, F. GSK3 Alpha and GSK3 Beta are Necessary for Axon Formation. FEBS Lett. 2007, 581, 1579–1586. [Google Scholar] [CrossRef]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25–35 and D-Gal-induced Neurotoxicity in PC12 Cells and D-Gal-induced Cognitive Dysfunction in Mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef]
- Ali, T.; Kim, M.O. Melatonin Ameliorates Amyloid Beta-induced Memory Deficits, Tau Hyperphosphorylation and Neurodegeneration via PI3/Akt/GSK3β Pathway in the Mouse Hippocampus. J. Pineal Res. 2015, 59, 47–59. [Google Scholar] [CrossRef]
- Fitton, J.; Stringer, D.; Karpiniec, S. Therapies from fucoidan: An update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Gerbst, A.G.; Ushakova, N.A.; Tsvetkova, E.A.; Dmitrenok, A.S.; Usov, A.I.; Nifantiev, N.E. Anticoagulant and antithrombotic activities of modified xylofucan sulfate from the brown alga Punctaria plantaginea. Carbohydr. Polym. 2016, 136, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Dong, C.; Yin, J.; Shen, J.; Tian, J.; Li, C. Neuroprotective effect of fucoidan on H2O2-induced apoptosis in PC12 cells via activation of PI3K/Akt pathway. Cell. Mol. Neurobiol. 2012, 32, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Van Noort, J.M.; Bsibsi, M. Toll-like receptors in the CNS: Implications for neurodegeneration and repair. Prog. Brain Res. 2009, 175, 139–148. [Google Scholar]
- Liu, S.; Liu, Y.; Hao, W.; Wolf, L.; Kiliaan, A.J.; Penke, B.; Rübe, C.E.; Walter, J.; Heneka, M.T.; Hartmann, T.; et al. TLR2 is a primary receptor for Alzheimer’s amyloid β peptide to trigger neuroinflammatory activation. J. Immunol. 2012, 188, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between sulfated polysaccharides from sea brown algae and Toll-like receptors on HEK293 eukaryotic cells in vitro. Bull. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef]
- Okolie, C.L.; CK Rajendran, S.R.; Udenigwe, C.C.; Aryee, A.N.; Mason, B. Prospects of brown seaweed polysaccharides (BSP) as prebiotics and potential immunomodulators. J. Food Biochem. 2017, 41, e12392. [Google Scholar] [CrossRef]
- Liu, X.; Cao, S.; Zhang, X. Modulation of gut microbiota–brain axis by probiotics, prebiotics, and diet. J. Agric. Food Chem. 2015, 63, 7885–7895. [Google Scholar] [CrossRef]
- Kim, S.; Kang, S.S.; Choi, S.I.; Kim, G.H.; Imm, J.Y. Ecklonia cava extract containing dieckol suppresses RANKL-induced osteoclastogenesis via MAP kinase/NF-κB pathway inhibition and heme oxygenase-1 induction. J. Microbiol. Biotechnol. 2019, 29, 11–20. [Google Scholar] [CrossRef]
- Choi, G.N.; Kim, J.H.; Kwak, J.H.; Jeong, C.; Jeong, H.R.; Lee, U.; Heo, H.J. Effect of Quercetin on Learning and Memory Performance in ICR Mice under Neurotoxic Trimethyltin Exposure. Food Chem. 2012, 132, 1019–1024. [Google Scholar] [CrossRef]
- Ha, J.S.; Jin, D.E.; Park, S.K.; Park, C.H.; Seung, T.W.; Bae, D.W.; Kim, D.O.; Heo, H.J. Antiamnesic effect of Actinidia arguta extract intake in a mouse model of TMT-induced learning and memory dysfunction. Evid. Based Complement. Altern. Med. 2015, 2015, 876484. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a Water-Maze Procedure for Studying Spatial Learning in the Rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Vincent, D.; Segonzac, G.; Vincent, M.C. Colorimetric Determination of Acetylcholine by the Hestrin Hydroxylamine Reaction and Its Application in Pharmacy. Ann. Pharm. Fr. 1958, 16, 179–185. [Google Scholar] [PubMed]
- Souza, B.W.; Cerqueira, M.A.; Bourbon, A.I.; Pinheiro, A.C.; Martins, J.T.; Teixeira, J.A.; Coimbrab, M.A.; Vicente, A.A. Chemical characterization and antioxidant activity of sulfated polysaccharide from the red seaweed Gracilaria birdiae. Food Hydrocoll. 2012, 27, 287–292. [Google Scholar] [CrossRef]
- Cataldi, T.R.; Campa, C.; De Benedetto, G.E. Carbohydrate analysis by high-performance anion-exchange chromatography with pulsed amperometric detection: The potential is still growing. Fresenius J. Anal. Chem. 2000, 368, 739–758. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mw (kDa) | Sulfate (%) | Monosaccharide Composition (Relative Area, %) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Arabinose | Fucose | Galactose | Glucose | Rhamnose | Xylose | Other | |||
| 110.78 | 3.03% | 2.03 | 15.19 | 9.62 | 11.42 | 1.44 | 23.77 | 36.53 | |
| Sample | Dieckol (% (w/w)) |
|---|---|
| Polyphenol extract | 4.8 ± 0.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoo, S.K.; Han, H.J.; Chung, D.H.; Kim, D.-O.; Kim, G.-H.; Heo, H.J. Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation. Mar. Drugs 2019, 17, 591. https://doi.org/10.3390/md17100591
Park SK, Kang JY, Kim JM, Yoo SK, Han HJ, Chung DH, Kim D-O, Kim G-H, Heo HJ. Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation. Marine Drugs. 2019; 17(10):591. https://doi.org/10.3390/md17100591
Chicago/Turabian StylePark, Seon Kyeong, Jin Yong Kang, Jong Min Kim, Seul Ki Yoo, Hye Ju Han, Dong Hwa Chung, Dae-Ok Kim, Gun-Hee Kim, and Ho Jin Heo. 2019. "Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation" Marine Drugs 17, no. 10: 591. https://doi.org/10.3390/md17100591
APA StylePark, S. K., Kang, J. Y., Kim, J. M., Yoo, S. K., Han, H. J., Chung, D. H., Kim, D. -O., Kim, G. -H., & Heo, H. J. (2019). Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation. Marine Drugs, 17(10), 591. https://doi.org/10.3390/md17100591