Eicosapentaenoic Acid Extraction from Nannochloropsis gaditana Using Carbon Dioxide at Supercritical Conditions

 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
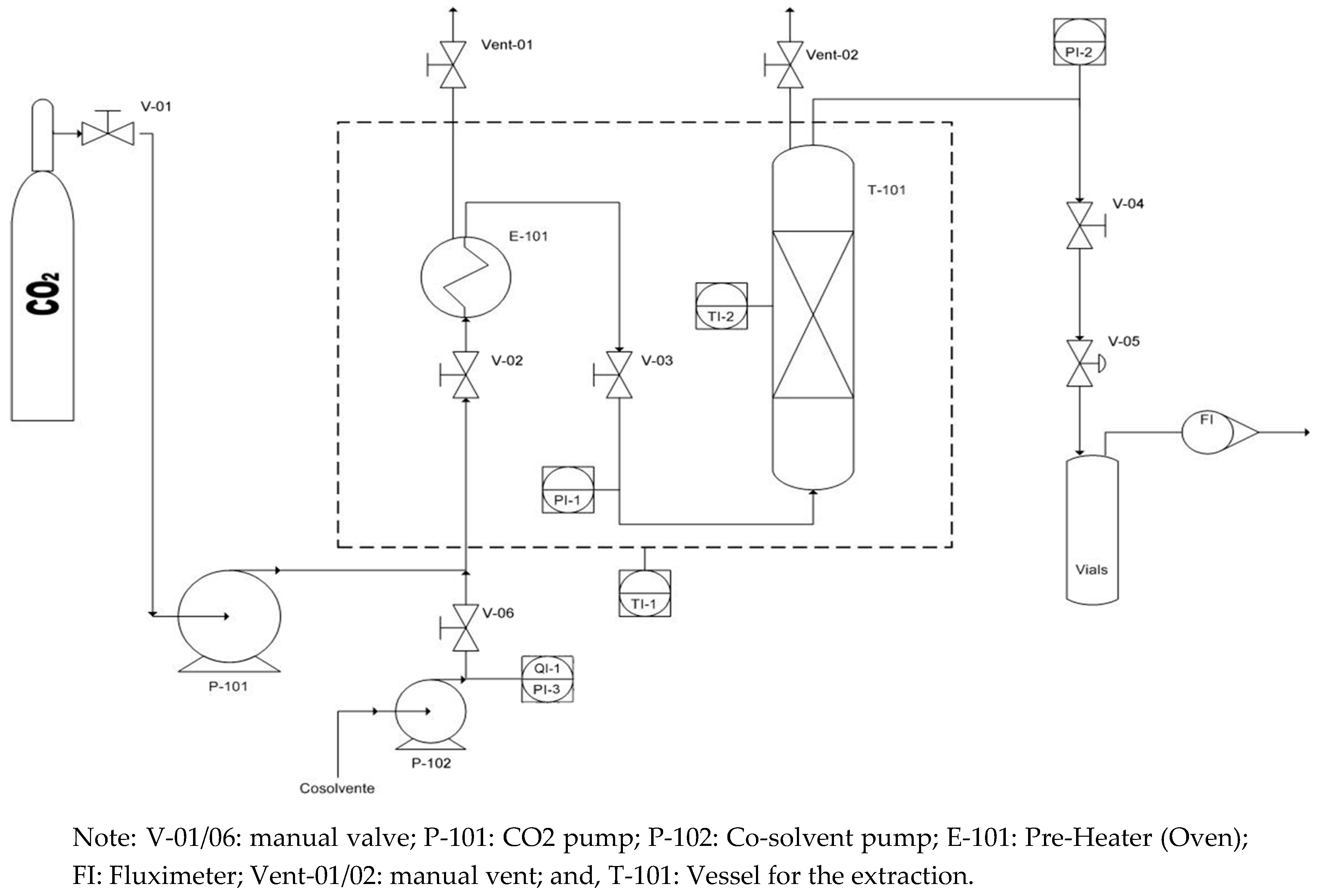
2. Materials and Methods
2.1. Microalgal Biomass and Chemical Composition
2.2. Chemicals
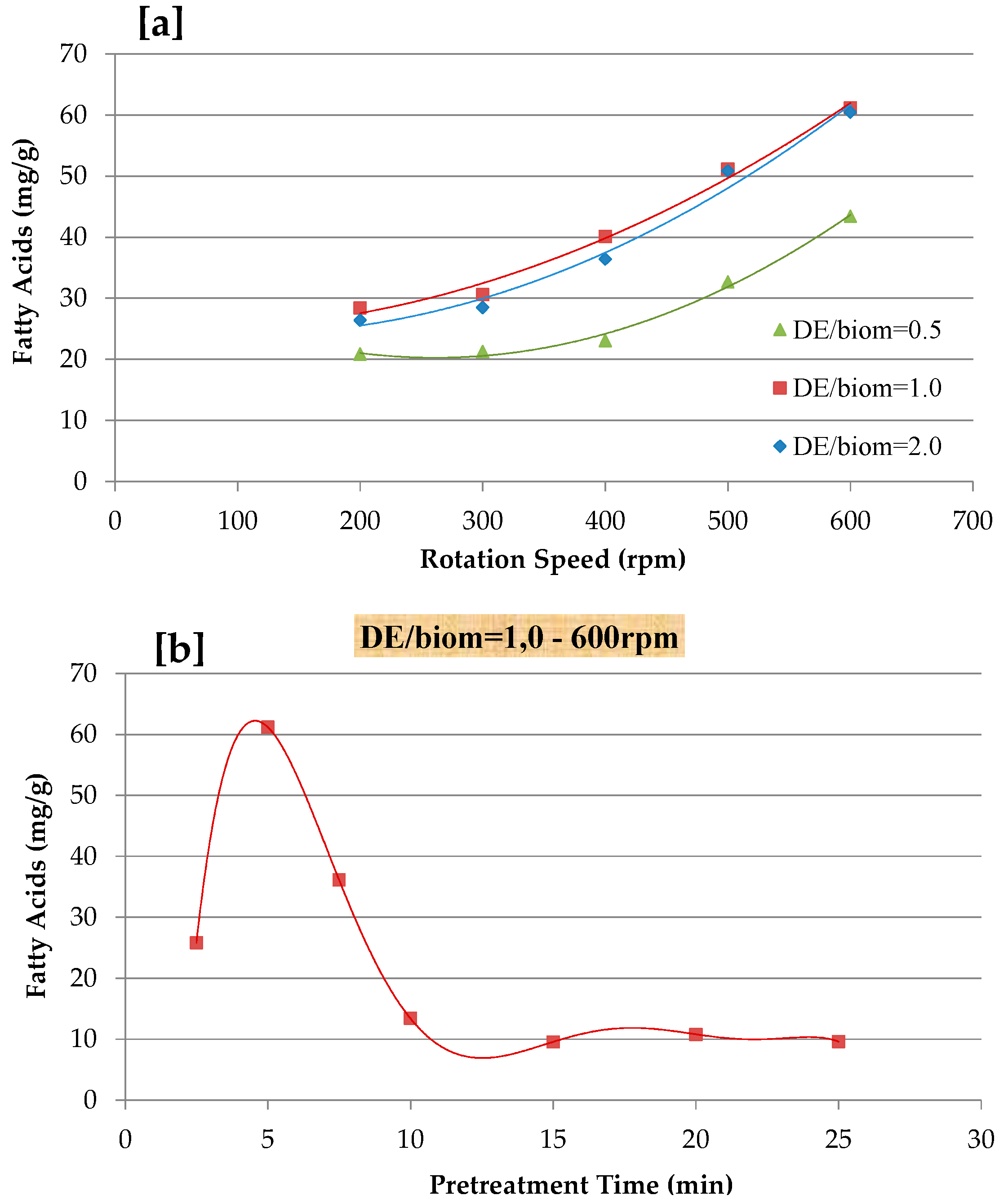
2.3. Mechanical Pre-Treatment of Biomass
2.4. CO2 Supercritical Extraction Experiments
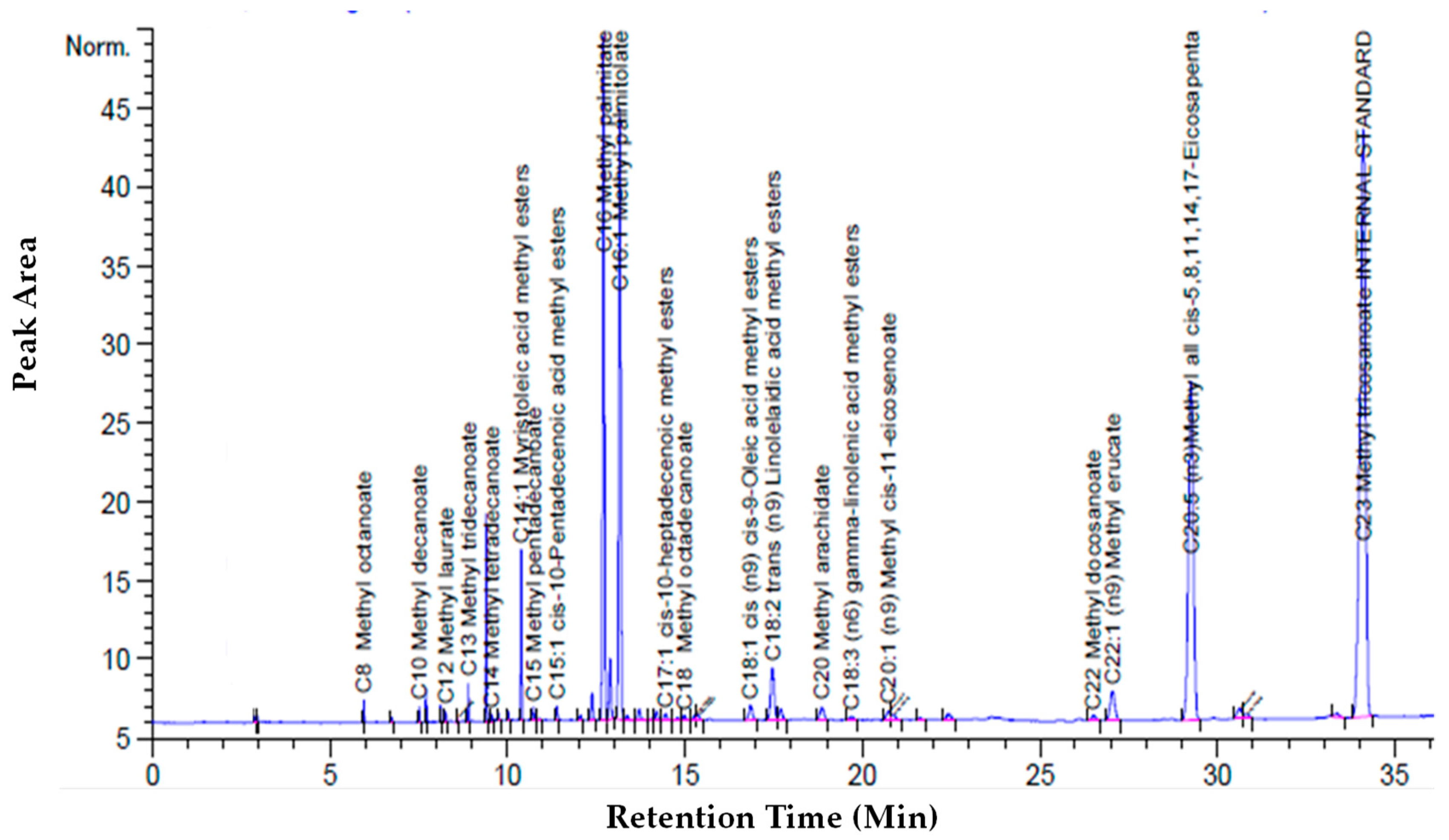
2.5. Analytical Methods
3. Results and Discussion
3.1. Effect of Mechanical Pre-Treatment on Fatty Acid Recovery
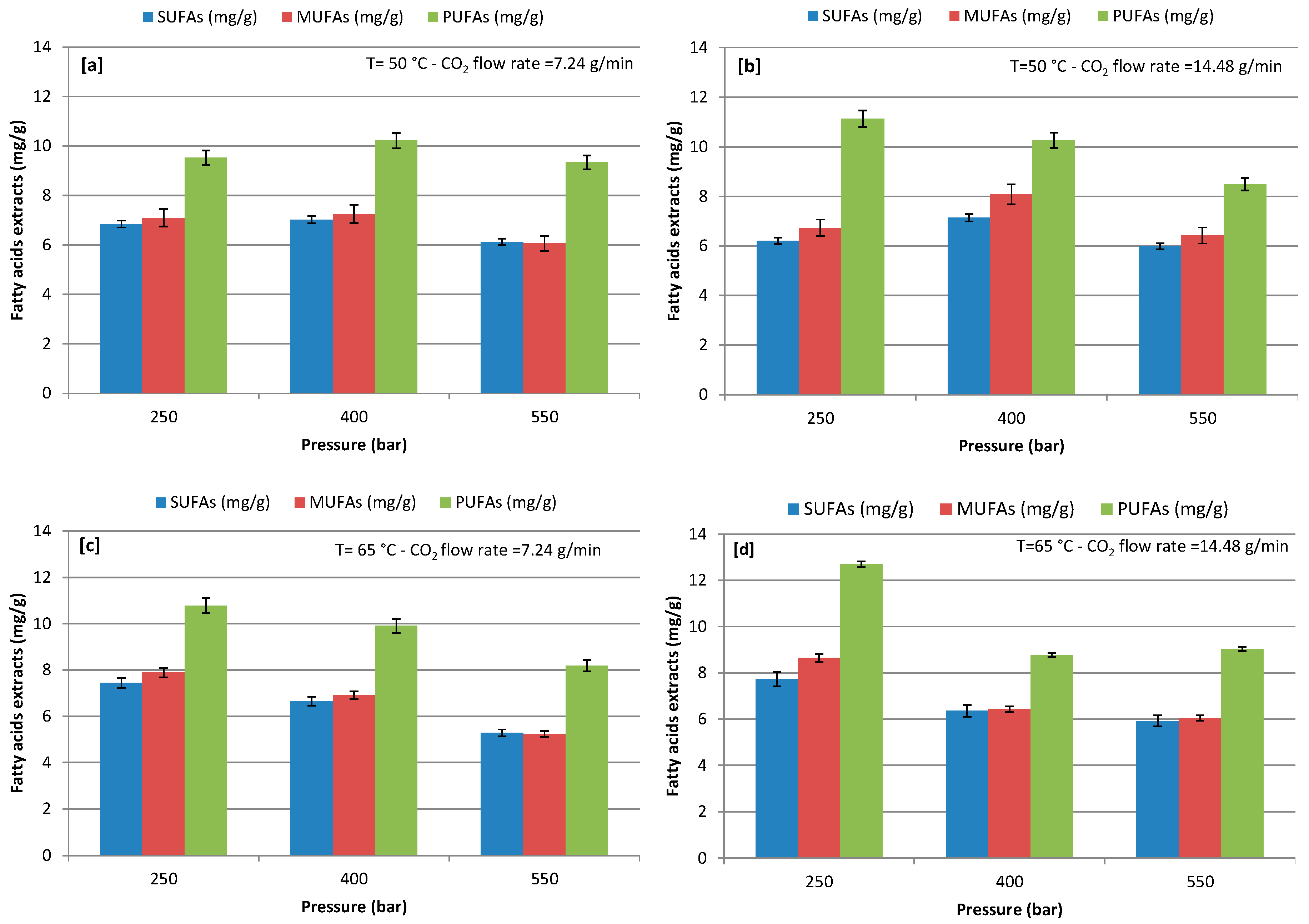
3.2. Effect of Extraction Pressure on Lipids Recovery
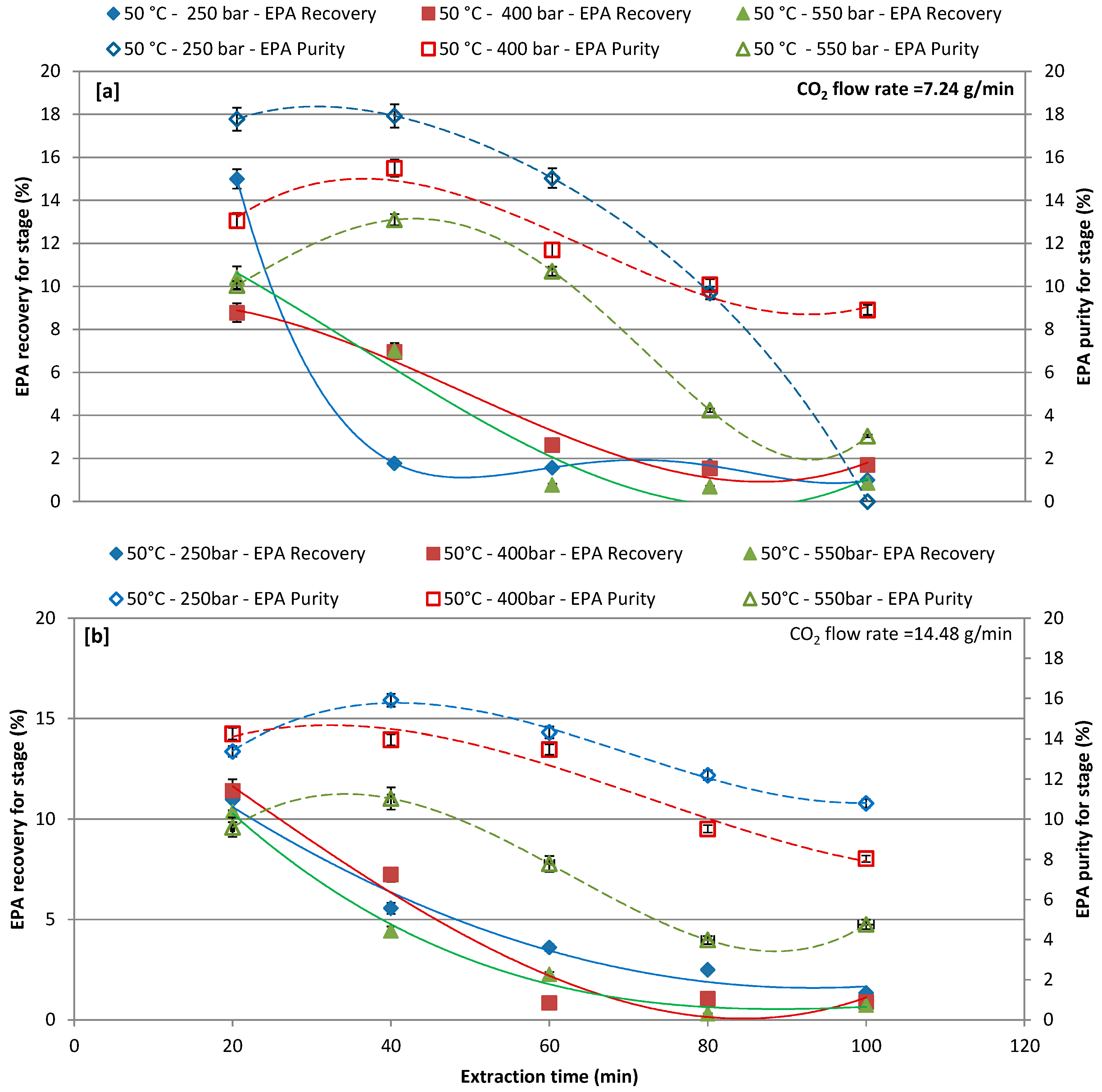
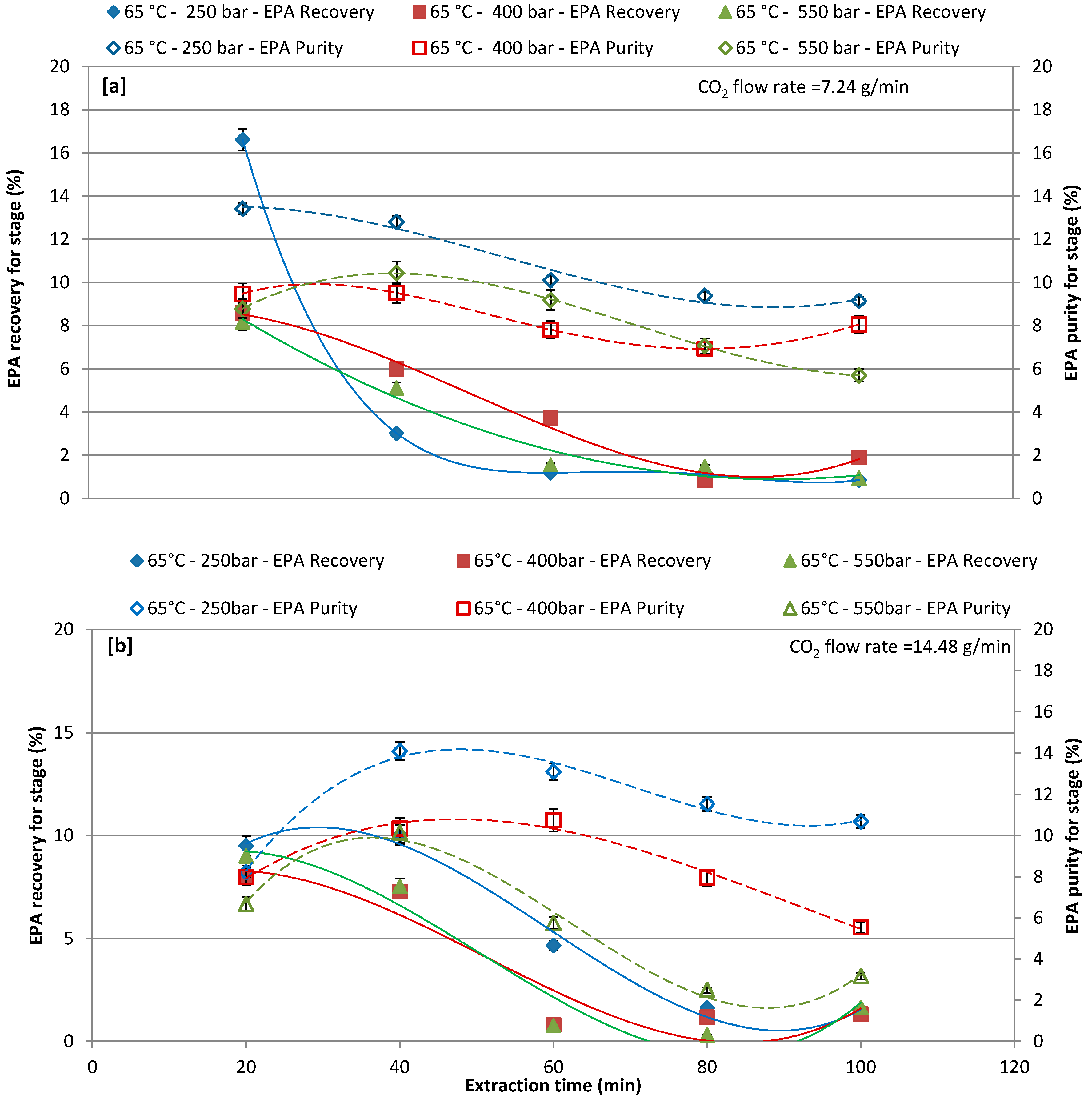
3.3. Effect of Different Pressure with CO2 Flow Rate of 7.24 and 14.48 g/min at 50 °C and 65 °C on EPA Recovery and Purity over Extraction Time
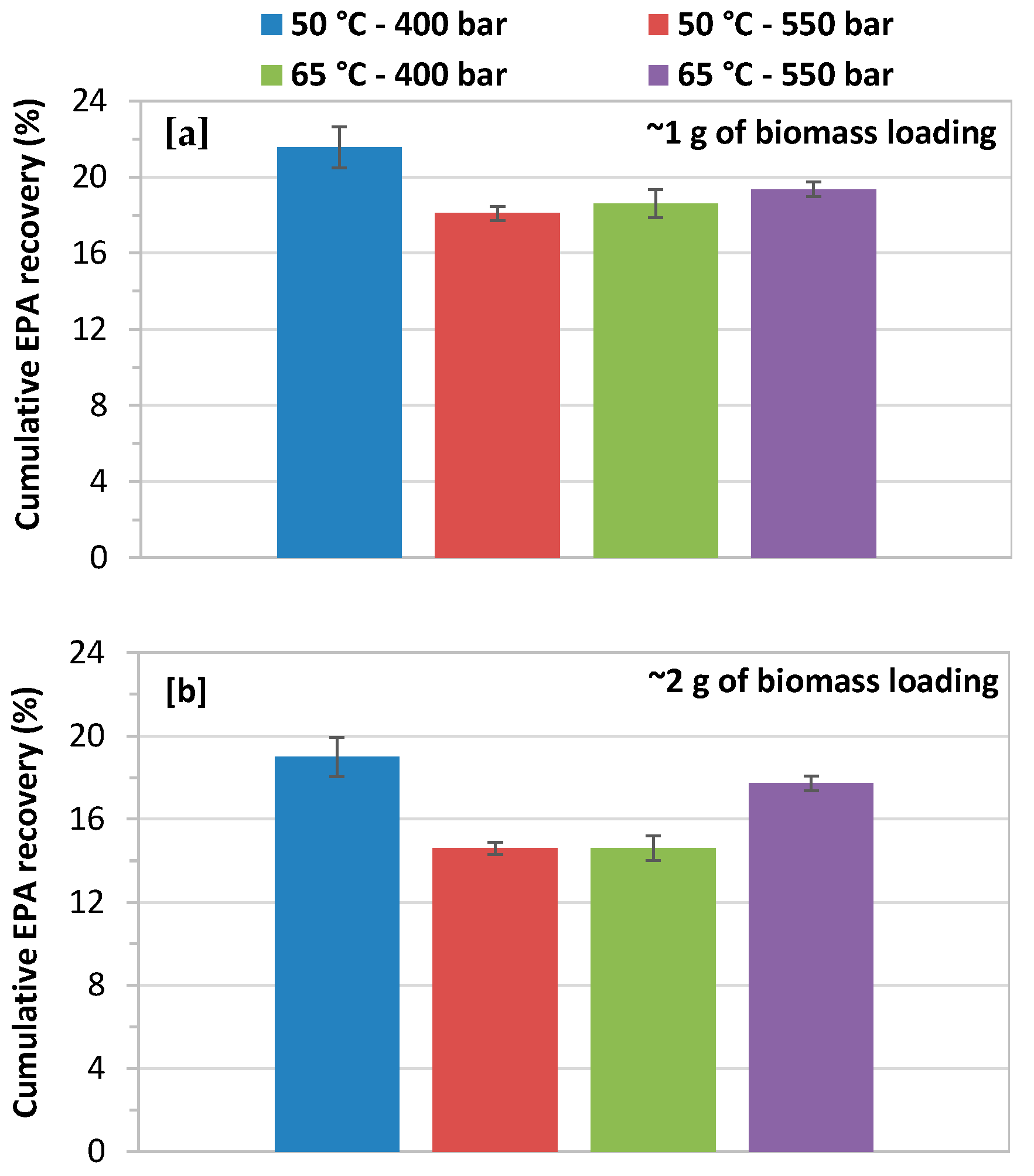
3.4. Effect of Biomass Loading on Cumulative EPA Recovery
3.5. Comparison with Literature for Lipid Recovery Using CO2-SF
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mirón, A.S.; García, M.C.C.; Gómez, A.C.; Camacho, F.G.; Grima, E.M.; Chisti, Y. Shear stress tolerance and biochemical characterization of Phaeodactylum tricornutum in quasi steady-state continuous culture in outdoor photobioreactors. Biochem. Eng. J 2003, 16, 287–297. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Microalgae biofuels: A critical review of issues, problems and the way forward. Biotechnol. Adv. 2012, 30, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, A.; Limonti, C.; Mehariya, S.; Molino, A.; Calabrò, V. Biofuel Production and Phosphorus Recovery through an Integrated Treatment of Agro-Industrial Waste. Sustainability 2018, 11, 52. [Google Scholar] [CrossRef]
- Xu, F.; Hu, H.; Cong, W.; Cai, Z.; Ouyang, F. Growth characteristics and eicosapentaenoic acid production by Nannochloropsis sp. in mixotrophic conditions. Biotechnol. Lett. 2004, 26, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Wei, C.; Zhao-Ling, C.; Fan, O. Effects of organic carbon sources on cell growth and eicosapentaenoic acid content of Nannochloropsis sp. J. Appl. Phycol. 2004, 16, 499–503. [Google Scholar] [CrossRef]
- Ma, X.-N.; Chen, T.-P.; Yang, B.; Liu, J.; Chen, F. Lipid Production from Nannochloropsis. Mar. Drugs 2016, 14, 61. [Google Scholar] [CrossRef]
- Mobin, S.; Alam, F. Some Promising Microalgal Species for Commercial Applications: A review. Energy Procedia 2017, 110, 510–517. [Google Scholar] [CrossRef]
- Scholz, M.J.; Weiss, T.L.; Jinkerson, R.E.; Jing, J.; Roth, R.; Goodenough, U.; Posewitz, M.C.; Gerken, H.G. Ultrastructure and Composition of the Nannochloropsis gaditana Cell Wall. Eukaryot. Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef]
- Chua, E.T.; Schenk, P.M. A biorefinery for Nannochloropsis: Induction, harvesting, and extraction of EPA-rich oil and high-value protein. Bioresour. Technol. 2017, 244, 1416–1424. [Google Scholar] [CrossRef]
- Meng, Y.; Jiang, J.; Wang, H.; Cao, X.; Xue, S.; Yang, Q.; Wang, W. The characteristics of TAG and EPA accumulation in Nannochloropsis oceanica IMET1 under different nitrogen supply regimes. Bioresour. Technol. 2015, 179, 483–489. [Google Scholar] [CrossRef]
- Kim, C.W.; Sung, M.-G.; Nam, K.; Moon, M.; Kwon, J.-H.; Yang, J.-W. Effect of monochromatic illumination on lipid accumulation of Nannochloropsis gaditana under continuous cultivation. Bioresour. Technol. 2014, 159, 30–35. [Google Scholar] [CrossRef]
- Ruxton, C.H.S.; Reed, S.C.; Simpson, M.J.A.; Millington, K.J. The health benefits of omega-3 polyunsaturated fatty acids: A review of the evidence. J. Hum. Nutr. Diet. 2004, 17, 449–459. [Google Scholar] [CrossRef]
- Agostoni, C.; Bresson, J.; Fairweather-Tait, S.; Flynn, A.; Golly, I.; Korhonen, H.; Lagiou, P.; Løvik, M.; Marchelli, R.; Martin, A. Scientific opinion on the tolerable upper intake level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA): EFSA panel on dietetic products, nutrition and allergies (NDA). EFSA J. 2012, 10, 1–48. [Google Scholar]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Rodríguez, J.; González-Céspedes, A.M.; Cerón-García, M.C.; Fernández-Sevilla, J.M.; Acién-Fernández, F.G.; Molina-Grima, E. A quantitative study of eicosapentaenoic acid (EPA) production by Nannochloropsis gaditana for aquaculture as a function of dilution rate, temperature and average irradiance. Appl. Microbiol. Biotechnol. 2014, 98, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.K.; Lewis, D.M.; Ashman, P.J. Disruption of microalgal cells for the extraction of lipids for biofuels: Processes and specific energy requirements. Biomass Bioenergy 2012, 46, 89–101. [Google Scholar] [CrossRef]
- Molino, A.; Rimauro, J.; Casella, P.; Cerbone, A.; Larocca, V.; Chianese, S.; Karatza, D.; Mehariya, S.; Ferraro, A.; Hristoforou, E.; Musmarra, D. Extraction of astaxanthin from microalga Haematococcus pluvialis in red phase by using Generally Recognized As Safe solvents and accelerated extraction. J. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Lee, S.Y.; Cho, J.M.; Chang, Y.K.; Oh, Y.-K. Cell disruption and lipid extraction for microalgal biorefineries: A review. Bioresour. Technol. 2017, 244, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ghasemi Naghdi, F.; Garg, S.; Adarme-Vega, T.C.; Thurecht, K.J.; Ghafor, W.A.; Tannock, S.; Schenk, P.M. A comparative study: The impact of different lipid extraction methods on current microalgal lipid research. Microb. Cell Fact. 2014, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Pieber, S.; Schober, S.; Mittelbach, M. Pressurized fluid extraction of polyunsaturated fatty acids from the microalga Nannochloropsis oculata. Biomass Bioenergy 2012, 47, 474–482. [Google Scholar] [CrossRef]
- Cardoso, L.C.; Serrano, C.M.; Rodríguez, M.R.; de la Ossa, E.J.M.; Lubián, L.M. Extraction of carotenoids and fatty acids from microalgae using supercritical technology. Am. J. Anal. Chem. 2012, 3, 877. [Google Scholar] [CrossRef]
- Di Sanzo, G.; Mehariya, S.; Martino, M.; Larocca, V.; Casella, P.; Chianese, S.; Musmarra, D.; Balducchi, R.; Molino, A. Supercritical Carbon Dioxide Extraction of Astaxanthin, Lutein, and Fatty Acids from Haematococcus pluvialis Microalgae. Mar. Drugs 2018, 16, 334. [Google Scholar] [CrossRef] [PubMed]
- Couto, R.M.; Simões, P.C.; Reis, A.; Da Silva, T.L.; Martins, V.H.; Sánchez-Vicente, Y. Supercritical fluid extraction of lipids from the heterotrophic microalga Crypthecodinium cohnii. Eng. Life Sci. 2010, 10, 158–164. [Google Scholar] [CrossRef]
- Yen, H.-W.; Yang, S.-C.; Chen, C.-H.; Jesisca; Chang, J.-S. Supercritical fluid extraction of valuable compounds from microalgal biomass. Bioresour. Technol. 2015, 184, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Rodríguez, N.; Beltrán, S.; Jaime, I.; de Diego, S.M.; Sanz, M.T.; Carballido, J.R. Production of omega-3 polyunsaturated fatty acid concentrates: A review. Innov. Food Sci. Emerg. Technol. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Reyes, F.A.; Mendiola, J.A.; Ibañez, E.; del Valle, J.M. Astaxanthin extraction from Haematococcus pluvialis using CO2-expanded ethanol. J. Supercrit. Fluids 2014, 92, 75–83. [Google Scholar] [CrossRef]
- Zou, T.-B.; Jia, Q.; Li, H.-W.; Wang, C.-X.; Wu, H.-F. Response Surface Methodology for Ultrasound-Assisted Extraction of Astaxanthin from Haematococcus pluvialis. Mar. Drugs 2013, 11, 1644–1655. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Deng, D.-F.; Dominy, W. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 2012, 354–355, 50–55. [Google Scholar] [CrossRef]
- Wang, L.; Weller, C.L. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol. 2006, 17, 300–312. [Google Scholar] [CrossRef]
- Molino, A.; Mehariya, S.; Iovine, A.; Larocca, V.; Di Sanzo, G.; Martino, M.; Casella, P.; Chianese, S.; Musmarra, D. Extraction of Astaxanthin and Lutein from Microalga Haematococcus pluvialis in the Red Phase Using CO2 Supercritical Fluid Extraction Technology with Ethanol as Co-Solvent. Mar. Drugs 2018, 16, 432. [Google Scholar] [CrossRef] [PubMed]
- Đurović, S.; Šorgić, S.; Popov, S.; Radojković, M.; Zeković, Z. Isolation and GC Analysis of Fatty Acids: Study Case of Stinging Nettle Leaves. Available online: https://www.intechopen.com/books/carboxylic-acid-key-role-in-life-sciences/isolation-and-gc-analysis-of-fatty-acids-study-case-of-stinging-nettle-leaves (accessed on 20 February 2019).
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and Efficient Extraction Methods for Marine-Derived Compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [PubMed]
- Jaime, L.; Mendiola, J.A.; Ibáñez, E.; Martin-Álvarez, P.J.; Cifuentes, A.; Reglero, G.; Señoráns, F.J. β-Carotene Isomer Composition of Sub- and Supercritical Carbon Dioxide Extracts. Antioxidant Activity Measurement. J. Agric. Food Chem. 2007, 55, 10585–10590. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.; Bhattacharjee, P. Supercritical carbon dioxide extraction of antioxidant rich fraction from Phormidium valderianum: Optimization of experimental process parameters. Algal Res. 2014, 3, 49–54. [Google Scholar] [CrossRef]
- Chen, K.-T.; Cheng, C.-H.; Wu, Y.-H.; Lu, W.-C.; Lin, Y.-H.; Lee, H.-T. Continuous lipid extraction of microalgae using high-pressure carbon dioxide. Bioresour. Technol. 2013, 146, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Safi, C.; Camy, S.; Frances, C.; Varela, M.M.; Badia, E.C.; Pontalier, P.-Y.; Vaca-Garcia, C. Extraction of lipids and pigments of Chlorella vulgaris by supercritical carbon dioxide: Influence of bead milling on extraction performance. J. Appl. Phycol. 2014, 26, 1711–1718. [Google Scholar] [CrossRef]
- Kim, S.-K.; Chojnacka, K. Marine Algae Extracts: Processes, Products, and Applications, 2 Volume Set; Wiley-VCH Verlag Gmbh & Co. KGaA: Weinheim, Germany, 2015. [Google Scholar]
- Ryckebosch, E.; Bruneel, C.; Muylaert, K.; Foubert, I. Microalgae as an alternative source of omega-3 long chain polyunsaturated fatty acids. Lipid Technol. 2012, 24, 128–130. [Google Scholar] [CrossRef]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae characterization for consolidated and new application in human food, animal feed and nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef] [PubMed]
- UNI ISO 12966-2 Animal and Vegetables Fat and Oils. Gas Chromatography of Fatty Acid Methyl Esters. Part 2: Preparation of Methyl Esters of Fatty Acids; International Organization for Standardization: Geneva, Switzerland, 2011.
- UNI ISO 12966-4 Animal and Vegetables Fat and Oils. Gas Chromatography of Fatty Acid Methyl Esters. Part 4: Determination by Capillary Chromatography; International Organization for Standardization: Geneva, Switzerland, 2015.
- Show, K.-Y.; Lee, D.-J.; Tay, J.-H.; Lee, T.-M.; Chang, J.-S. Microalgal drying and cell disruption―Recent advances. Bioresour. Technol. 2015, 184, 258–266. [Google Scholar] [CrossRef]
- Howlader, M.S.; Rai, N.; Todd French, W. Improving the lipid recovery from wet oleaginous microorganisms using different pretreatment techniques. Bioresour. Technol. 2018, 267, 743–755. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, D.; Wu, C.; Xu, A.; Xia, W.; Wang, Z.; Wen, F.; Yu, D. Influence of a facile pretreatment process on lipid extraction from Nannochloropsis sp. through an enzymatic hydrolysis reaction. RSC Adv. 2017, 7, 53270–53277. [Google Scholar] [CrossRef]
- Elst, K.; Maesen, M.; Jacobs, G.; Bastiaens, L.; Voorspoels, S.; Servaes, K. Supercritical CO2 Extraction of Nannochloropsis sp.: A Lipidomic Study on the Influence of Pretreatment on Yield and Composition. Molecules 2018, 23, 1854. [Google Scholar] [CrossRef] [PubMed]
- Shene, C.; Monsalve, M.T.; Vergara, D.; Lienqueo, M.E.; Rubilar, M. High pressure homogenization of Nannochloropsis oculata for the extraction of intracellular components: Effect of process conditions and culture age. Eur. J. Lipid Sci. Technol. 2016, 118, 631–639. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Du, T.-B.; Pi, H.-C.; Jang, S.-M.; Lin, Y.-H.; Lee, H.-T. Comparative study of lipid extraction from microalgae by organic solvent and supercritical CO2. Bioresour. Technol. 2011, 102, 10151–10153. [Google Scholar] [CrossRef] [PubMed]
- Andrich, G.; Nesti, U.; Venturi, F.; Zinnai, A.; Fiorentini, R. Supercritical fluid extraction of bioactive lipids from the microalga Nannochloropsis sp. Eur. J. Lipid Sci. Technol. 2005, 107, 381–386. [Google Scholar] [CrossRef]
- Mouahid, A.; Crampon, C.; Toudji, S.-A.A.; Badens, E. Supercritical CO2 extraction of neutral lipids from microalgae: Experiments and modelling. J. Supercrit. Fluids 2013, 77, 7–16. [Google Scholar] [CrossRef]
- Crampon, C.; Boutin, O.; Badens, E. Supercritical Carbon Dioxide Extraction of Molecules of Interest from Microalgae and Seaweeds. Ind. Eng. Chem. Res. 2011, 50, 8941–8953. [Google Scholar] [CrossRef]
- Crampon, C.; Nikitine, C.; Zaier, M.; Lépine, O.; Tanzi, C.D.; Vian, M.A.; Chemat, F.; Badens, E. Oil extraction from enriched Spirulina platensis microalgae using supercritical carbon dioxide. J. Supercrit. Fluids 2017, 119, 289–296. [Google Scholar] [CrossRef]
- Cheung, P.C.K. Temperature and pressure effects on supercritical carbon dioxide extraction of n-3 fatty acids from red seaweed. Food Chem. 1999, 65, 399–403. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.P.; Martinez-Correa, H.A.; Paviani, L.C.; Cabral, F.A. Supercritical CO2 extraction of lipids and astaxanthin from Brazilian redspotted shrimp waste (Farfantepenaeus paulensis). J. Supercrit. Fluids 2011, 56, 164–173. [Google Scholar] [CrossRef]
- Ho, B.C.H.; Kamal, S.M.M.; Harun, M.R. Extraction of eicosapentaenoic acid from Nannochloropsis gaditana using sub-critical water extraction. Malays. J. Anal. Sci. 2018, 22, 619–625. [Google Scholar] [CrossRef]
- Awaluddin, S.A.; Thiruvenkadam, S.; Izhar, S.; Hiroyuki, Y.; Danquah, M.K.; Harun, R. Subcritical water technology for enhanced extraction of biochemical compounds from Chlorella vulgaris. Biomed Res. Int. 2016, 5816974, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Machmudah, S.; Shotipruk, A.; Goto, M.; Sasaki, M.; Hirose, T. Extraction of Astaxanthin from Haematococcus pluvialis Using Supercritical CO2 and Ethanol as Entrainer. Ind. Eng. Chem. Res. 2006, 45, 3652–3657. [Google Scholar] [CrossRef]
- Taher, H.; Al-Zuhair, S.; Al-Marzouqi, A.H.; Haik, Y.; Farid, M.; Tariq, S. Supercritical carbon dioxide extraction of microalgae lipid: Process optimization and laboratory scale-up. J. Supercrit. Fluids 2014, 86, 57–66. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition of N. gaditana | Concentration (mg/g) |
|---|---|
| Humidity | 47.2 |
| Ash | 100.5 |
| Total dietary fiber (TDF) | 40.2 |
| carbohydrates | 217.4 |
| proteins | 470.4 |
| Total lipids | 164.5 |
| FAs FAME (mg/g of total lipids) | 114.88 |
| mg/g of FAMEs | |
| Σ SFAs | 32.80 |
| Σ MUFAs | 29.96 |
| Σ PUFAs | 52.12 |
| of which: | |
| EPA | 42.28 |
| Operative Conditions | Total Extraction Yield (mg/g) | Total Lipid Yield (mg/g) | |||||
|---|---|---|---|---|---|---|---|
| N. gaditana Biomass Loading (g) | Bed Height (cm) | Porosity [−] | T (°C) | P (Bar) | CO2 Flow Rates (g/min) | ||
| 1.01 | 18.6 | 0.51 | 50 | 250 | 14.48 | 74.94 | 31.83 |
| 1.01 | 18.6 | 0.51 | 50 | 400 | 14.48 | 68.52 | 34.46 |
| 1.01 | 18.6 | 0.51 | 50 | 550 | 14.48 | 85.21 | 28.92 |
| 1.02 | 18.6 | 0.51 | 50 | 250 | 7.24 | 53.40 | 31.14 |
| 1.01 | 18.6 | 0.51 | 50 | 400 | 7.24 | 72.28 | 32.24 |
| 1.01 | 18.6 | 0.51 | 50 | 550 | 7.24 | 89.60 | 29.99 |
| 1.02 | 18.6 | 0.51 | 65 | 250 | 14.48 | 107.56 | 38.15 |
| 1.01 | 18.6 | 0.51 | 65 | 400 | 14.48 | 92.46 | 29.64 |
| 1.01 | 18.6 | 0.51 | 65 | 550 | 14.48 | 122.96 | 29.65 |
| 1.02 | 18.6 | 0.51 | 65 | 250 | 7.24 | 77.68 | 34.61 |
| 1.01 | 18.6 | 0.51 | 65 | 400 | 7.24 | 101.14 | 31.79 |
| 1.02 | 18.6 | 0.51 | 65 | 550 | 7.24 | 84.78 | 26.70 |
| 2.01 | 19.6 | 0.50 | 50 | 400 | 14.48 | 86.59 | 28.79 |
| 2.01 | 19.6 | 0.50 | 65 | 400 | 14.48 | 66.88 | 14.93 |
| 2.01 | 19.6 | 0.50 | 50 | 550 | 14.48 | 41.38 | 22.76 |
| 2.02 | 19.6 | 0.50 | 65 | 550 | 14.48 | 107.35 | 26.19 |
| Operative Temperature (°C) | Biomass Loading of 1.01 g | Biomass Loading of 2.01 g | ||||||
| CO2 Flow Rate (g/min) | ||||||||
| 7.24 | 14.48 | |||||||
| Operative Pressure (bar) | ||||||||
| 250 | 400 | 550 | 250 | 400 | 550 | 400 | 550 | |
| 50 | 8.46 | 9.12 | 8.35 | 10.14 | 9.06 | 7.6 | 7.97 | 6.13 |
| 65 | 9.63 | 8.92 | 7.3 | 11.5 | 7.82 | 8.13 | 6.13 | 7.44 |
| Species | Biomass Loading (g) | CO2 Flow Rate (g/min) | P (Bar) | T (°C) | Extraction Time (min) | Recovery # (%) | Ref. |
|---|---|---|---|---|---|---|---|
| C. vulgaris | 6 | 30 | 600 | 60 | 180 | 59.2 | [34] |
| P. valderianum | 10 | ~3.6 | 350 | 40 | 90 | 26.6 | [32] |
| Nannochloropsis sp. | 1.25 | 0.62 | 300 | 40 | 160 | 65.8 | [57] |
| Synechococccus sp. | 4.6 | 0.8 | 400 | 60 | 180 | 6.0 | [21] |
| C. vulgaris | 150 | 166.6 | 280 | 40 | 540 | 31.9 | [58] |
| Pavlova sp. | 10 | na | 306 | 60 | 360 | 40.0 | [47] |
| Tetraselmis sp. | 0.2 | 0.009 | 150 | 40 | 720 | 72.3 | [19] |
| N. gaditana | 1 | 7.24 | 250 | 65 | 100 | 23.2 | This study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molino, A.; Martino, M.; Larocca, V.; Di Sanzo, G.; Spagnoletta, A.; Marino, T.; Karatza, D.; Iovine, A.; Mehariya, S.; Musmarra, D. Eicosapentaenoic Acid Extraction from Nannochloropsis gaditana Using Carbon Dioxide at Supercritical Conditions. Mar. Drugs 2019, 17, 132. https://doi.org/10.3390/md17020132
Molino A, Martino M, Larocca V, Di Sanzo G, Spagnoletta A, Marino T, Karatza D, Iovine A, Mehariya S, Musmarra D. Eicosapentaenoic Acid Extraction from Nannochloropsis gaditana Using Carbon Dioxide at Supercritical Conditions. Marine Drugs. 2019; 17(2):132. https://doi.org/10.3390/md17020132
Chicago/Turabian StyleMolino, Antonio, Maria Martino, Vincenzo Larocca, Giuseppe Di Sanzo, Anna Spagnoletta, Tiziana Marino, Despina Karatza, Angela Iovine, Sanjeet Mehariya, and Dino Musmarra. 2019. "Eicosapentaenoic Acid Extraction from Nannochloropsis gaditana Using Carbon Dioxide at Supercritical Conditions" Marine Drugs 17, no. 2: 132. https://doi.org/10.3390/md17020132
APA StyleMolino, A., Martino, M., Larocca, V., Di Sanzo, G., Spagnoletta, A., Marino, T., Karatza, D., Iovine, A., Mehariya, S., & Musmarra, D. (2019). Eicosapentaenoic Acid Extraction from Nannochloropsis gaditana Using Carbon Dioxide at Supercritical Conditions. Marine Drugs, 17(2), 132. https://doi.org/10.3390/md17020132