Fatty Acid Profiles and Production in Marine Phytoplankton


Abstract
:1. Introduction
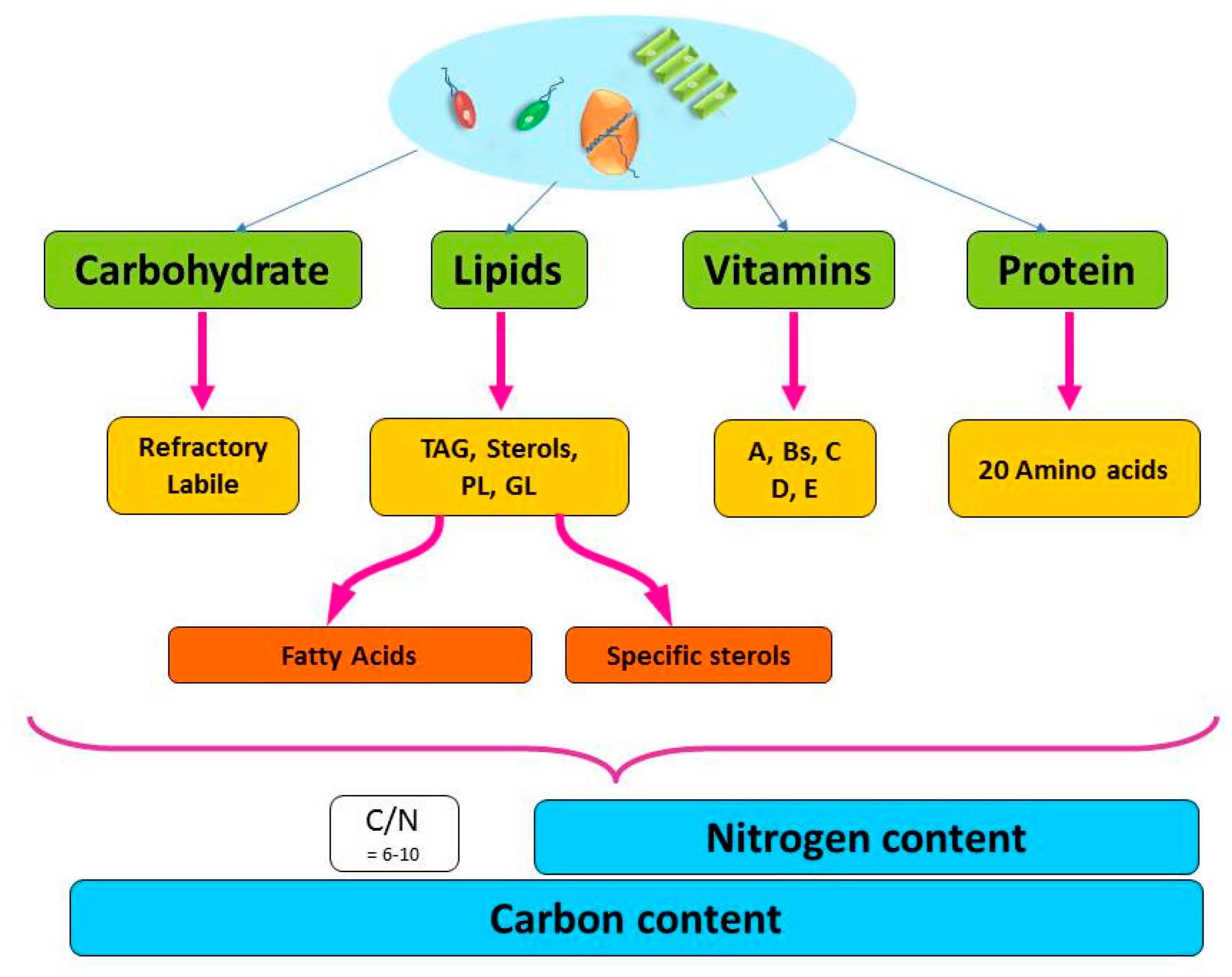
2. The Gross Chemical Composition of Phytoplankton
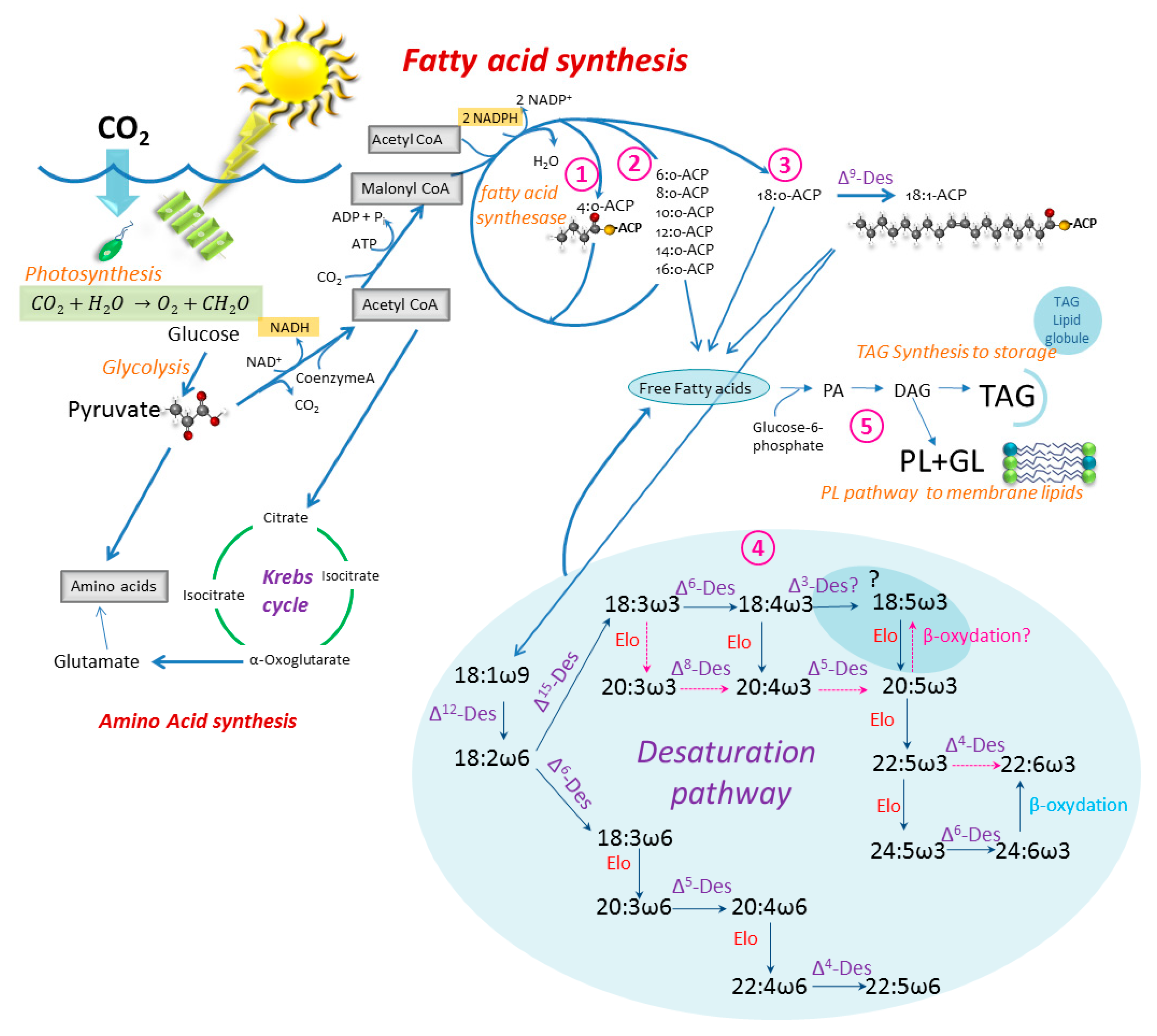
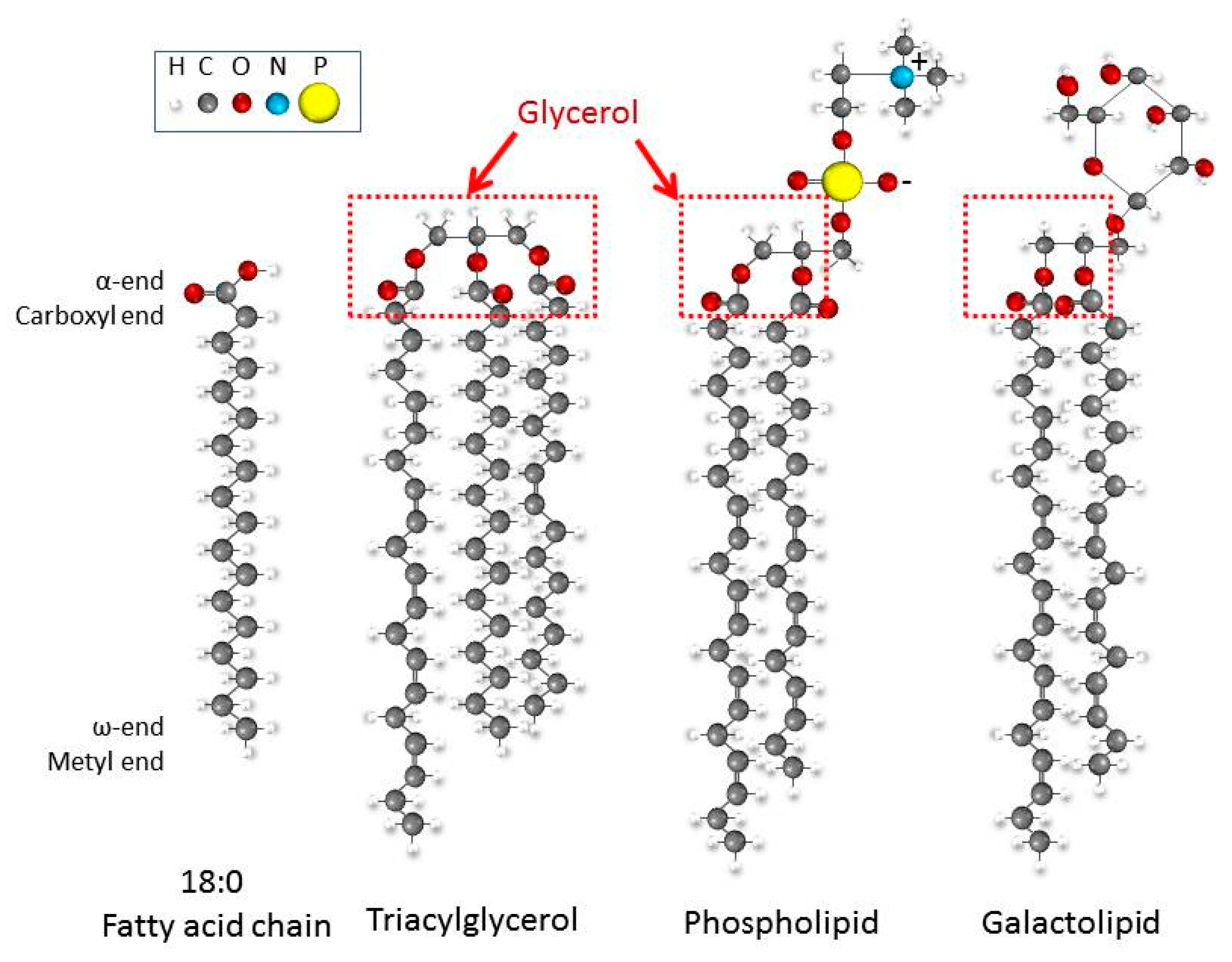
2.1. From Glucose to Fatty Acids
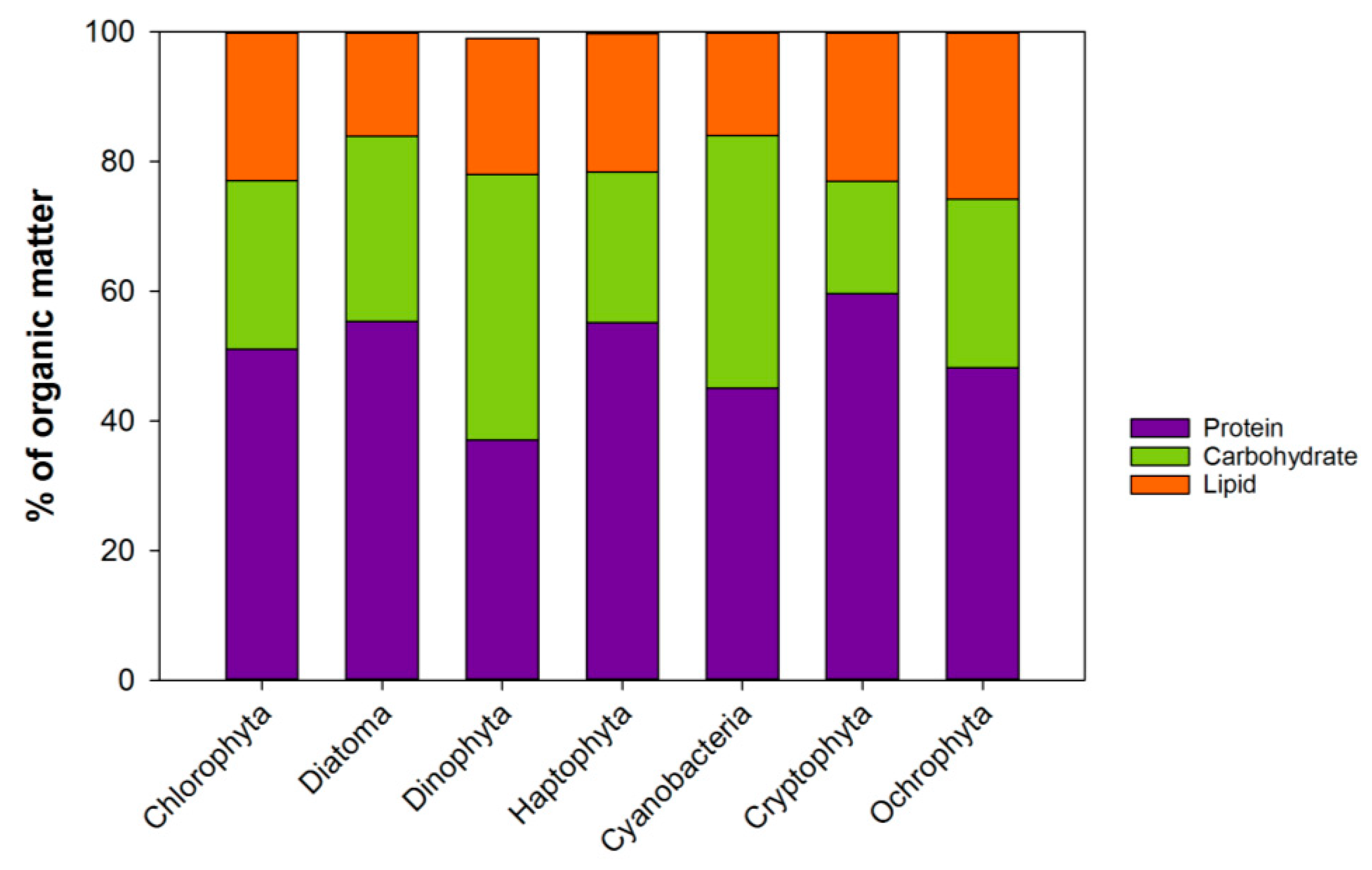
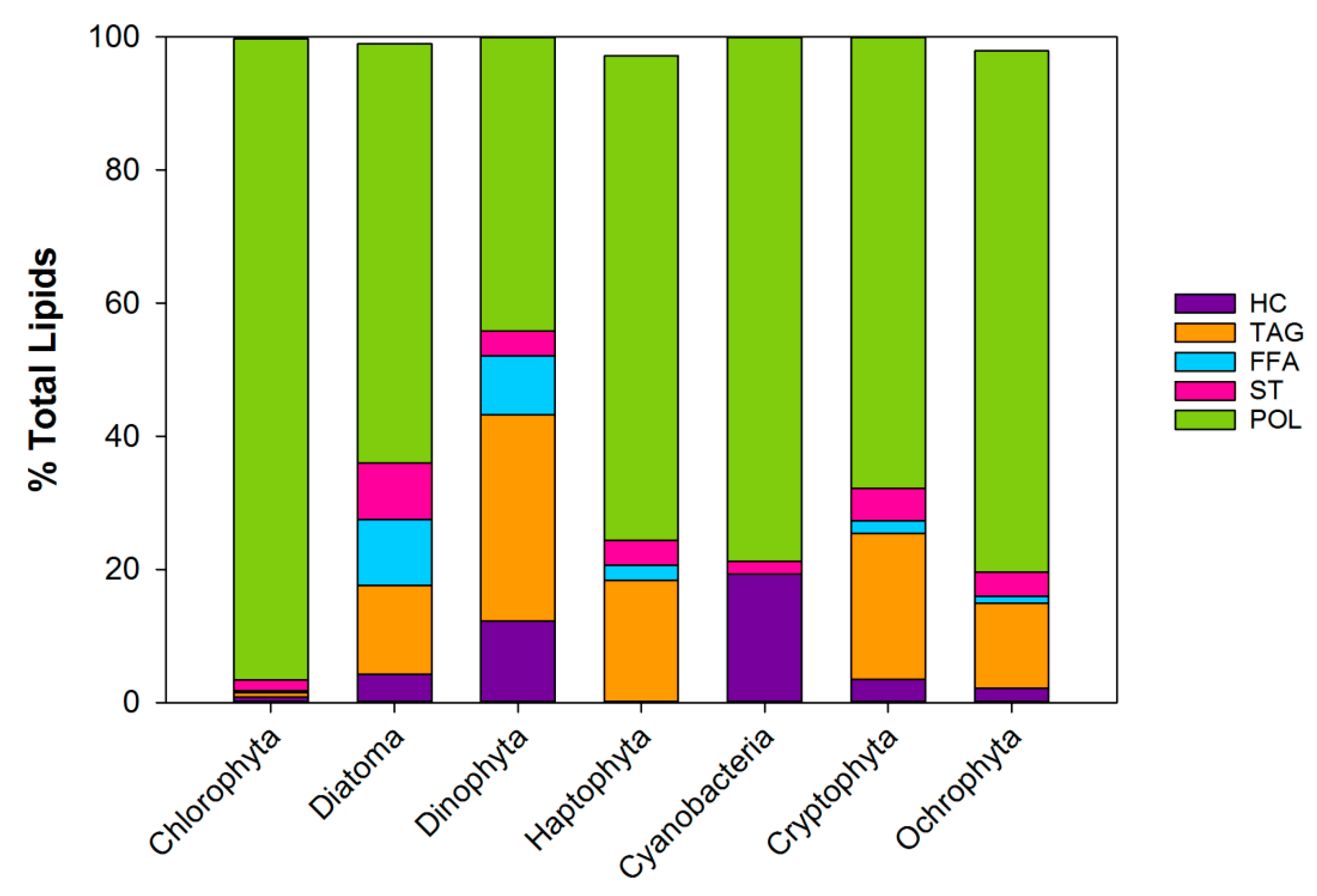
2.2. Specific Lipid Content in the Phytoplankton
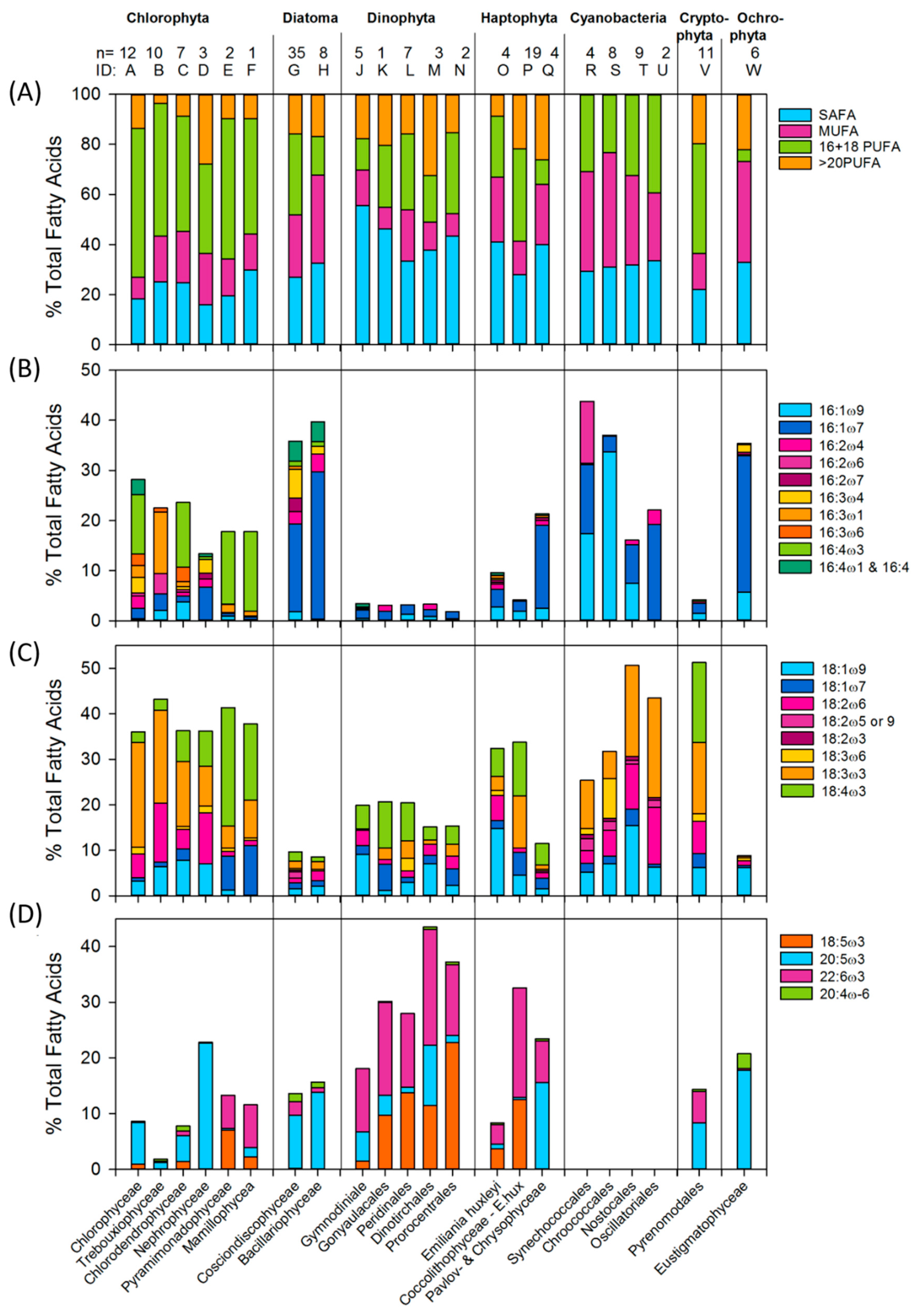
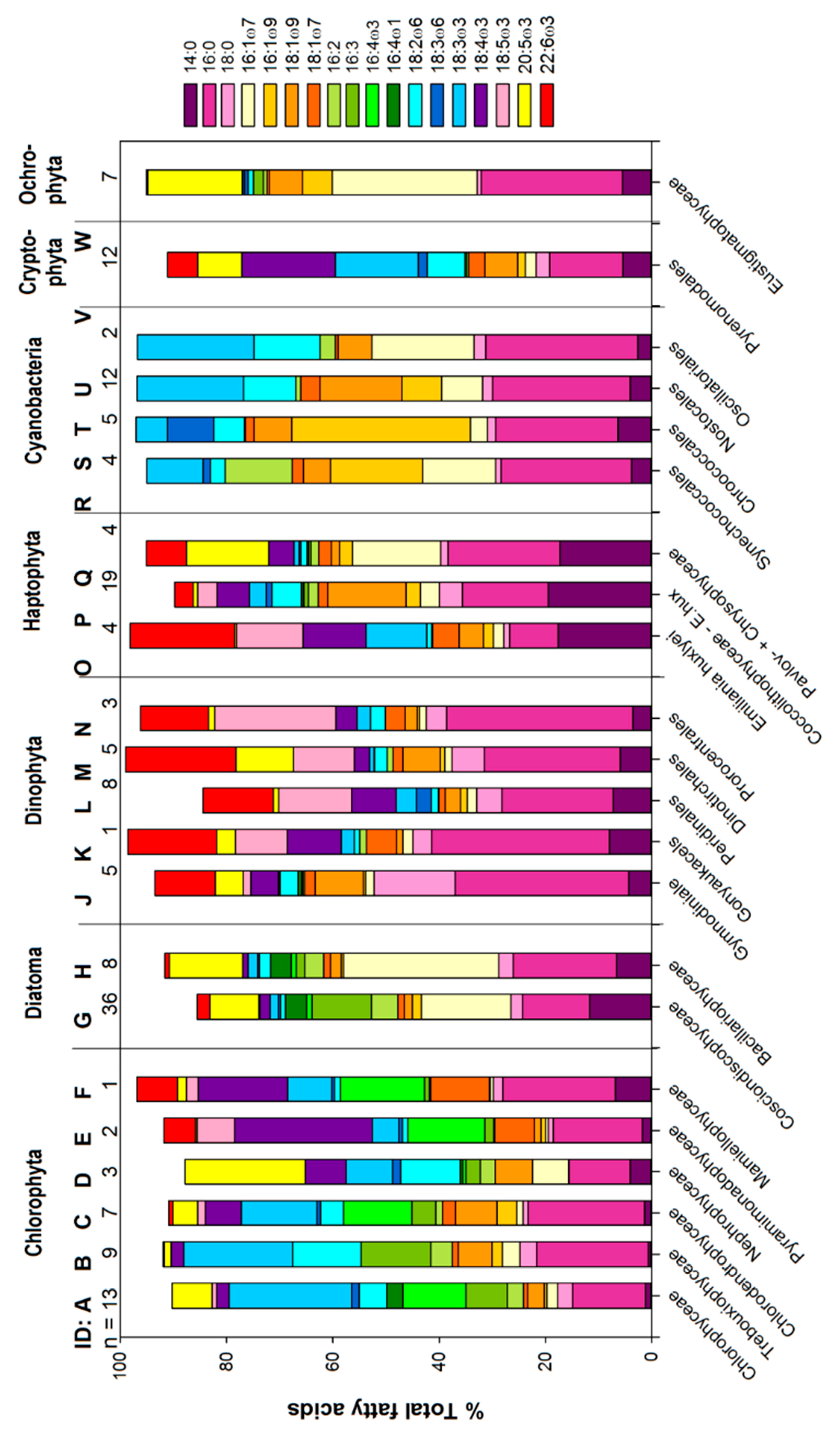
2.2.1. Fatty Acid Profiles
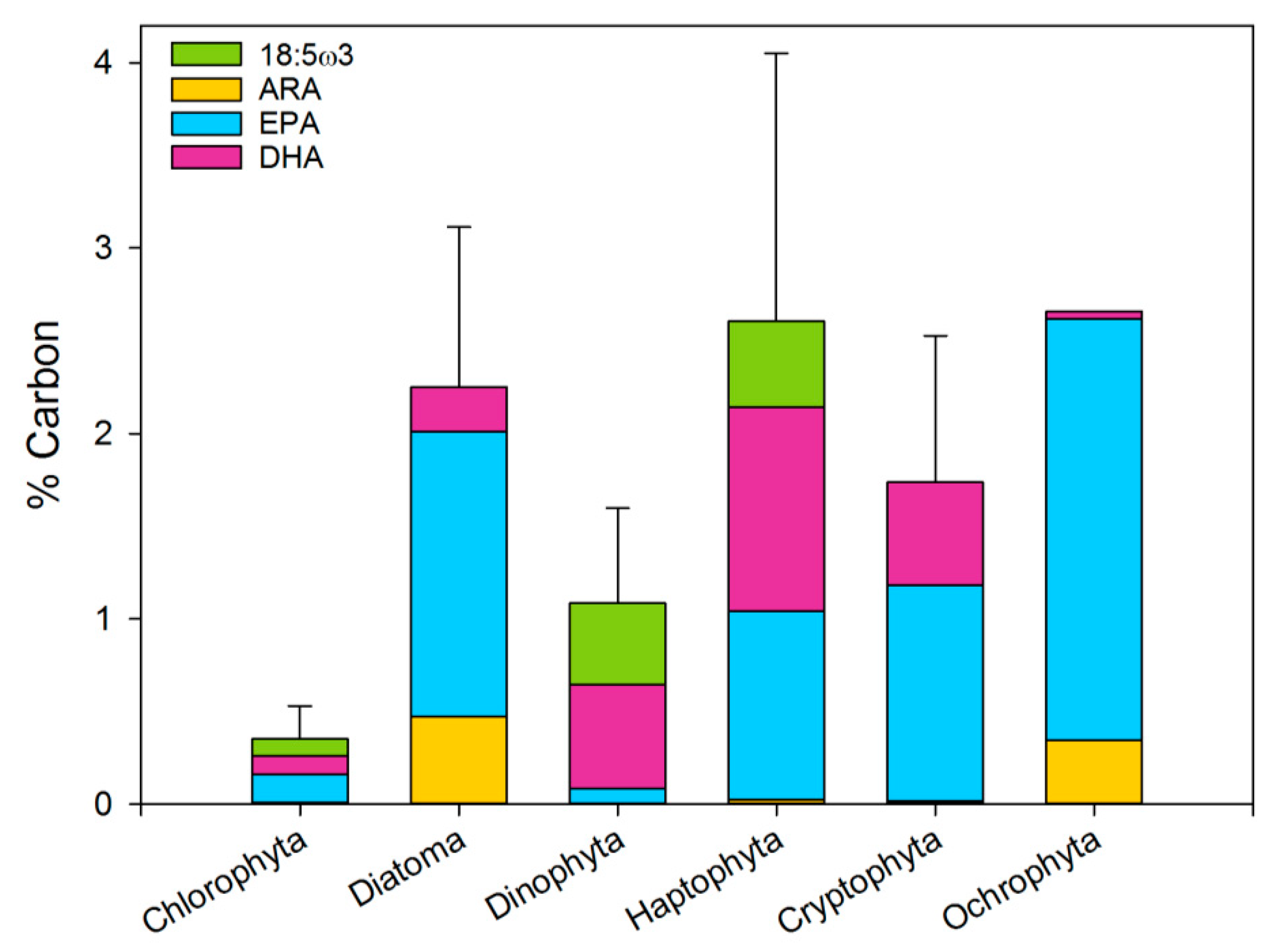
2.2.2. PUFA as a Fraction of Biomass
3. Environmental Effects on Lipid and Fatty Acid Composition
3.1. Physical Environment
3.2. Nutrients
3.3. Trace Metals
4. Discussion
5. Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Lee, R.F.; Nevenzel, J.C.; Paffenhöfer, G.-A. Importance of wax esters and other lipids in the marine food chain: Phytoplankton and copepods. Mar. Biol. 1971, 9, 99–108. [Google Scholar] [CrossRef]
- Graeve, M.; Kattner, G.; Hagen, W. Diet-induced changes in the fatty acid composition of Arctic herbivorous copepods: Experimental evidence of trophic markers. J. Exp. Mar. Biol. Ecol. 1994, 182, 97–110. [Google Scholar] [CrossRef]
- Sargent, J.R.; Falk-Petersen, S. The lipid biochemistry of calanoid copepods. Hydrobiologia 1988, 167–168, 101–114. [Google Scholar] [CrossRef]
- Marshall, S.; Orr, A. On the biology of Calanus finmarchicus. VII. Factors affecting egg production. J. Mar. Biol. Assoc. UK 1952, 30, 527–547. [Google Scholar] [CrossRef]
- Checkley, D.M.D. The egg production of a marine planktonic copepod in relation to its food supply: Laboratory studies. Limnol. Oceanogr. 1980, 25, 430–446. [Google Scholar] [CrossRef] [Green Version]
- Nassogne, A. Influence of food organisms on the development and culture of pelagic copepods. Helgol. Wiss. Meeresunters. 1970, 20, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Arnott, G.; Brand, G.; Kos, L. Effects of food quality and quantity on the survival, development, and egg production of Gladioferens pectinatus (Brady) (Copepoda: Calanoida). Aust. J. Mar. Freshw. Res. 1986, 37, 467–473. [Google Scholar] [CrossRef]
- Castell, J. Fatty acid metabolism in crustaceans. In Proceedings of the Second International Conference on Aquaculture Nutrition: Biochemical and Physiological Approaches to Shellfish Nutrition; Pruder, G.D., Langdon, C.J., Conklin, D.E., Eds.; Louisiana State University: Baton Rouge, LA, USA, 1982; pp. 124–145. [Google Scholar]
- Bourdier, G.; Amblard, C. Lipids in Acanthodiaptomus denticornis during starvation and fed on 3 different algae. J. Plankton Res. 1989, 11, 1201–1212. [Google Scholar] [CrossRef]
- Fraser, A.J.; Sargent, J.R.; Gamble, J.C.; Seaton, D.D. Formation and transfer of fatty acids in an enclosed marine food chain comprising phytoplankton, zooplankton and herring (Clupea harengus L.) larvae. Mar. Chem. 1989, 27, 1–18. [Google Scholar] [CrossRef]
- Guisande, C.; Harris, R. Effect of total organic content of eggs on hatching success and naupliar survival in the copepod Calanus helgolandicus. Limnol. Oceanogr. 1995, 40, 476–482. [Google Scholar] [CrossRef]
- Koski, M.; Yebra, L.; Dutz, J.; Jónasdóttir, S.H.; Vidoudez, C.; Jakobsen, H.H.; Pohnert, G.; Nejstgaard, J.C. The effect of egg versus seston quality on hatching success, naupliar metabolism and survival of Calanus finmarchicus in mesocosms dominated by Phaeocystis and diatoms. Mar. Biol. 2012, 159, 643–660. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H.; Visser, A.W.; Jespersen, C. Assessing the role of food quality in the production and hatching of Temora longicornis eggs. Mar. Ecol. Prog. Ser. 2009, 382, 139–150. [Google Scholar] [CrossRef]
- Kanazawa, A.; Teshima, S.; Tokiwa, S. Nutritional requirements of prawn. 7. Effect of dieatry lipids on growth. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 849–856. [Google Scholar] [CrossRef]
- Pond, D.W.; Harris, R.; Head, R.; Harbour, D. Environmental and nutritional factors determining seasonal variability in the fecundity and egg viability of Calanus helgolandicus in coastal waters off Plymouth, UK. Mar. Ecol. Prog. Ser. 1996, 143, 45–63. [Google Scholar] [CrossRef]
- Kleppel, G.; Burkart, C. Egg production and the nutritional environment of Acartia tonsa: The role of food quality in copepod nutrition. ICES J. Mar. Sci. 1995, 52, 297–304. [Google Scholar] [CrossRef]
- Guisande, C.; Riveiro, I.; Maneiro, I. Comparisons among the amino acid composition of females, eggs and food to determine the relative importance of food quantity and food quality to copepod. Mar. Ecol. Prog. Ser. 2000, 202, 135–142. [Google Scholar] [CrossRef]
- Grosse, J.; Brussaard, C.; Boschker, H. Nutrient limitation driven dynamics of amino acids and fatty acids in coastal phytoplankton. Limnol. Oceanogr. 2018. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G. Nutritional properties of microaglae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Fishler, R.; Schneller, A. Chemical composition of dietary species of marine unicellular algae and rotifers with emphasis on fatty acids. Mar. Biol. 1987, 95, 31–36. [Google Scholar] [CrossRef]
- Parsons, T.R.; Stephens, K.; Strickland, J.D.H. On the chemical composition of eleven species of marine phytoplankters. J. Fish. Board Can. 1961, 18, 1001–1016. [Google Scholar] [CrossRef]
- Gatenby, C.M.; Orcutt, D.M.; Kreeger, D.A.; Parker, B.C.; Jones, V.A.; Neves, R.J. Biochemical composition of three algal species proposed as food for captive freshwater mussels. J. Appl. Phycol. 2003, 15, 1–11. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. A Journey from Light to Darkness. Fatty Acids in the Marine Ecosystem: From Photosynthesis to Copepod Lipids and Sequestration. Doctoral Thesis, National Institute of Aquatic Resources, Technical University of Denmark, Lyngby, Denmark, 2015; p. 118. [Google Scholar]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mühlroth, A.; Li, K.; Røkke, G.; Winge, P.; Olsen, Y.; Hohmann-Marriott, M.; Vadstein, O.; Bones, A. Pathways of lipid metabolism in marine algae, co-expression network, bottlenecks and candidate genes for enhanced production of EPA and DHA in species of Chromista. Mar. Drugs 2013, 11, 4662–4697. [Google Scholar] [CrossRef] [PubMed]
- Monroig, Ó.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs 2013, 11, 3998–4018. [Google Scholar] [CrossRef] [PubMed]
- Sargent, J.; Henderson, R. Lipids. In The Biological Chemistry of Marine Copepods; Corner, E., O’Hara, S., Eds.; Oxford Science Publications: New York, NY, USA, 1986; pp. 59–108. [Google Scholar]
- Parrish, C.C.; French, V.M.; Whiticar, M.J. Lipid class and fatty acid composition of copepods (Calanus finmarchicus, C. glacialis, Pseudocalanus sp., Tisbe furcata and Nitokra lacustris) fed various combinations of autotrophic and heterotrophic protists. J. Plankton Res. 2012, 34, 356–375. [Google Scholar] [CrossRef]
- Caramujo, M.-J.; Boschker, H.T.S.; Admiraal, W. Fatty acid profiles of algae mark the development and composition of harpacticoid copepods. Freshw. Biol. 2008, 53, 77–90. [Google Scholar] [CrossRef]
- De Troch, M.; Boeckx, P.; Cnudde, C.; Van Gansbeke, D.; Vanreusel, A.; Vincx, M.; Caramujo, M.-J. Bioconversion of fatty acids at the basis of marine food webs: Insights from a compound-specific stable isotope analysis. Mar. Ecol. Prog. Ser. 2012, 465, 53–67. [Google Scholar] [CrossRef]
- Arndt, C.; Sommer, U. Effect of algal species and concentration on development and fatty acid composition of two harpacticoid copepods, Tisbe sp. and Tachidius discipes, and a discussion about their suitability for marine fish larvae. Aquac. Nutr. 2014, 20, 44–59. [Google Scholar] [CrossRef]
- Pelley, J.W. 10-Fatty Acid and Triglyceride Metabolism. In Elsevier’s Integrated Review Biochemistry, 2nd ed.; Pelley, J.W., Ed.; W.B. Saunders: Philadelphia, PA, USA, 2012; pp. 81–88. ISBN 978-0-323-07446-9. [Google Scholar]
- Singh, A.; Ward, O.P. Microbial production of docosahexaenoic acid. Adv. Appl. Microbiol. 1997, 45, 271–312. [Google Scholar] [PubMed]
- Arts, M.; Evans, M.; Robarts, R.D. Seasonal patterns of total and energy reserve lipids of dominant zooplanktonic crustaceans from a hyper-eutrophic lake. Oecologia 1992, 90, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.; Tocher, D. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 211–236. ISBN 978-0-387-89366-2. [Google Scholar]
- Yu, W.-L.; Ansari, W.; Schoepp, N.G.; Hannon, M.J.; Mayfield, S.P.; Burkart, M.D. Modifications of the metabolic pathways of lipid and triacylglycerol production in microalgae. Microb. Cell Factories 2011, 10, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, M.; Shiraiwa, Y. Biosynthesis of lipids and hydrocarbons in algae. In Photosynthesis; Dubinsky, S., Ed.; InTech: Vienna, Austria, 2013; pp. 331–356. ISBN 978-953-51-1161-0. [Google Scholar]
- Heydarizadeh, P.; Poirier, I.; Loizeau, D.; Ulmann, L.; Mimouni, V.; Schoefs, B.; Bertrand, M. Plastids of marine phytoplankton produce bioactive pigments and lipids. Mar. Drugs 2013, 11, 3425–3471. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, J.; St John, M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [PubMed]
- Harwood, J.L.; Guschina, I.A. The versatility of algae and their lipid metabolism. Biochimie 2009, 91, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Sayanova, O.; Haslam, R.P.; Calerón, M.V.; López, N.R.; Worthy, C.; Rooks, P.; Allen, M.; Napier, J.A. Identification and functional characterisation of genes encoding the omega-3 polyunsaturated fatty acid biosynthetic pathway from the coccolithophore Emiliania huxleyi. Phytochemistry 2011, 72, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.; Leftley, J.W.; Sargent, J.R. Lipid composition and biosynthesis in the marine dinoflagellate Crypthecodinium cohnii. Phytochemistry 1988, 27, 1679–1683. [Google Scholar] [CrossRef]
- Volkman, J.K.; Jeffrey, S.W.; Nichols, P.D.; Rogers, G.I.; Garland, C.D. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1989, 128, 219–240. [Google Scholar] [CrossRef]
- Al-Hasan, R.H.; Ali, A.M.; Radwan, S.S. Lipids, and their constituent fatty acids, of Phaeocystis sp. from the Arabian Gulf. Mar. Biol. 1990, 105, 9–14. [Google Scholar] [CrossRef]
- Dunstan, G.; Volkman, J.K.; Jeffrey, S.W.; Barrett, S. Biochemical composition of microalgae from the green algal classes Chlorophyceae and Prasinohyceae. 2. Lipid classes and fatty acids. J. Exp. Mar. Biol. Ecol. 1992, 161, 115–134. [Google Scholar] [CrossRef]
- Dunstan, G.; Volkman, J.K.; Barrett, S. Essential polyunsaturated fatty acids from 14 species of diatom (Bacillariophyceae). Phytochemistry 1993, 35, 155–161. [Google Scholar] [CrossRef]
- Parrish, C.C.; Bodennec, G.; Gentien, P. Time courses of intracellular and extracellular lipid classes in batch cultures of the toxic dinoflagellate, Gymnodinium cf. nagasakiense. Mar. Chem. 1994, 48, 71–78. [Google Scholar] [CrossRef]
- Lombardi, A.T.; Wangersky, P.J. Particulate lipid class composition of three marine phytoplankters Chaetoceros gracilis, Isochrysis galbana (Tahiti) and Dunaliella tertiolecta grown in batch culture. Hydrobiologia 1995, 306, 1–6. [Google Scholar] [CrossRef]
- Nordbäck, J.; Lundberg, E.; Christie, W. Separation of lipid classes from marine particulate material by HPLC on a polyvinyl alcohol-bonded stationary phase using dual-channel evaporative light-scattering detection. Mar. Chem. 1998, 60, 165–175. [Google Scholar] [CrossRef]
- Galloway, A.W.E.; Winder, M. Partitioning the relative importance of phylogeny and environmental conditions on phytoplankton fatty acids. PLoS ONE 2015, 10, e0130053. [Google Scholar] [CrossRef] [PubMed]
- Cañavate, J.P. Advancing assessment of marine phytoplankton community structure and nutritional value from fatty acid profiles of cultured microalgae. Rev. Aquac. 2018. [Google Scholar] [CrossRef]
- Scott, C.C.L.; Kwasniewski, S.; Falk-Petersen, S.; Sargent, J.R.J. Species differences, origins and functions of fatty alcohols and fatty acids in the wax esters and phospholipids of Calanus hyperboreus, C. glacialis and C. finmarchicus from Arctic waters. Mar. Ecol. Prog. Ser. 2002, 235, 127–134. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Hagen, W.; Kattner, G.; Clarke, A.; Sargent, J. Lipids, trophic relationships, and biodiversity in Arctic and Antarctic krill. Can. J. Fish. Aquat. Sci. 2000, 57, 178–191. [Google Scholar] [CrossRef]
- Stübing, D.; Hagen, W. Fatty acid biomarker ratios-suitable trophic indicators in Antarctic euphausiids? Polar Biol. 2003, 26, 774–782. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. Effects of food quality on the reproductive success of Acartia tonsa and Acartia hudsonica: Laboratory observations. Mar. Biol. 1994, 121, 67–81. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Ackman, R.G.; Jangaard, P.; Hoyle, R.R.J.; Brockerhoff, H. Origin of marine fatty acids. I. Analyses of the fatty acids produced by the diatom Skeletonema costatum. J. Fish. Board Can. 1964, 21, 747–756. [Google Scholar] [CrossRef]
- Ackman, R.G.; Tocher, C.S.; McLachlan, J. Marine phytoplankter fatty acids. J. Fish. Board Can. 1968, 25, 1603–1620. [Google Scholar] [CrossRef]
- Chuecas, L.; Riley, J.P. Component fatty acids of the total lipids of some marine phytoplankton. J. Mar. Biol. Assoc. UK 1969, 49, 97–116. [Google Scholar] [CrossRef]
- Pugh, P.R. Changes in the fatty acid composition of Coscinodiscus eccentricus with culture-age and salinity. Mar. Biol. 1971, 124, 118–124. [Google Scholar]
- Whyte, J.N. Fatty acid profiles from direct methanolysis of lipids in tissue of cultured species. Aquaculture 1988, 75, 193–203. [Google Scholar] [CrossRef]
- Claustre, H.A.; Poulet, S.A.; Williams, R.; Marty, J.C.; Coombs, S.; Ben Mlih, F.; Jezequel-Martin, V. A biochemical investigation of a Phaeocystis sp. bloom in the Irish Sea. J. Mar. Biol. Assoc. UK 1990, 70, 197–207. [Google Scholar] [CrossRef]
- Mourente, G.; Lubian, L.M.; Odriozola, J.M. Total fatty acid composition as a taxonomic index of some marine microalgae used as food in marine aquaculture. Hydrobiologia 1990, 203, 147–154. [Google Scholar] [CrossRef]
- Ahlgren, G.; Lundstedt, L.; Brett, M.; Forsberg, C. Lipid composition and food quality of some freshwater phytoplankton for cladoceran zooplankter. J. Plankton Res. 1990, 12, 809–818. [Google Scholar] [CrossRef]
- Ahlgren, G.; Gustafsson, I.B.; Bobeg, M. Fatty acid content and chemical composition of freshwater microalgae. J. Phycol. 1992, 28, 37–50. [Google Scholar] [CrossRef]
- Nichols, P.D.; Skerratt, J.H.; Davidson, A.; Burton, H.; Mcmeekin, T.A. Lipids of cultured Phaeocystis pouchetii: Signatures for food-web, biogeochemical and environmental studies in Antarctica and the Southern ocean. Phytochemistry 1991, 30, 3209–3214. [Google Scholar] [CrossRef]
- Virtue, P.; Nichols, P.D.D.; Nicol, S.; McMinn, A.; Sikes, E.L.L. The lipid composition of Euphausia superba Dana in relation to the nutritional value of Phaeocystis pouchetii (Hariot) Lagerheim. Antarct. Sci. 1993, 5, 169–177. [Google Scholar] [CrossRef]
- Viso, A.C.A.C.; Marty, J.C.J.C. Fatty acids from 28 marine microalgae. Phytochemistry 1993, 34, 1521–1533. [Google Scholar] [CrossRef]
- Ederington, M.C.; McManus, G.B.; Harvey, H.R. Trophic transfer of fatty acids, sterols, and a triterpenoid alcohol between bacteria, a ciliate, and the copepod Acartia tonsa. Limnol. Oceanogr. 1995, 40, 860–867. [Google Scholar] [CrossRef]
- Zhukova, N.; Aizdaicher, N. Fatty acid composition of 15 species of marine microalgae. Phytochemistry 1995, 39, 351–356. [Google Scholar] [CrossRef]
- Berge, J.; Gouygou, J.; Dubacq, J.; Durand, P. Reassessment of lipid composition of the diatom, Skeletonema costatum. Phytochemistry 1995, 39, 1017–1021. [Google Scholar] [CrossRef]
- Bell, M.; Pond, D.W. Lipid composition during growth of motile and coccolith forms of Emiliania huxleyi. Phytochemistry 1996, 41, 465–471. [Google Scholar] [CrossRef]
- Klein Breteler, W.; Schogt, N.; Baas, M.; Schouten, S.; Kraay, G.W. Trophic upgrading of food quality by protozoans enhancing copepod growth: Role of essential lipids. Mar. Biol. 1999, 135, 191–198. [Google Scholar] [CrossRef]
- Riebesell, U.; Revill, A.T.; Holdsworth, D.G.; Volkman, J.K. The effects of varying CO2 concentration on lipid composition and carbon isotope fractionation in Emiliania huxleyi. Geochim. Cosmochim. Acta 2000, 64, 4179–4192. [Google Scholar] [CrossRef]
- Cotonnec, G.; Brunet, C.; Sautour, B.; Thoumelin, G. Nutritive value and selection of food particles by copepods during a spring bloom of Phaeocystis sp. in the English Channel, as determined by pigment and fatty acid analyses. J. Plankton Res. 2001, 23, 693–703. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Shkrob, I.; Go, J.V. Dicarboxylic and fatty acid compositions of cyanobacteria of the genus Aphanizomenon. Biochem. Mosc. 2001, 66, 72–76. [Google Scholar] [CrossRef]
- Tang, K.W.; Jakobsen, H.H.; Visser, A.W. Phaeocystis globosa (Prymnesiophyceae) and the planktonic food web: Feeding, growth, and trophic interactions among grazers. Limnol. Oceanogr. 2001, 46, 1860–1870. [Google Scholar] [CrossRef]
- Gugger, M.; Lyra, C.; Suominen, I.; Tsitko, I.; Humbert, J.-F.; Salkinoja-Salonen, M.S.; Sivonen, K. Cellular fatty acids as chemotaxonomic markers of the genera Anabaena, Aphanizomenon, Microcystis, Nostoc and Planktothrix (cyanobacteria). Int. J. Syst. Evol. Microbiol. 2002, 52, 1007–1015. [Google Scholar] [PubMed]
- Broglio, E.; Jónasdóttir, S.H.; Calbet, A.; Jakobsen, H.H.; Saiz, E. Effect of heterotrophic versus autotrophic food on feeding and reproduction of the calanoid copepod Acartia tonsa: Relationship with prey fatty acid composition. Aquat. Microb. Ecol. 2003, 31, 267–278. [Google Scholar] [CrossRef]
- Ahlgren, G.; Hyenstrand, P. Nitrogen limitation effects of different nitrogen sources on nutritional quality of two freshwater organisms, Scenedesmus quadricauda (Chlorophyceae) and Synechococcus sp. (Cyanophyceae). J. Phycol. 2003, 39, 906–917. [Google Scholar] [CrossRef]
- Řezanka, T.; Dor, I.; Prell, A.; Dembitsky, V.M.; Řezanka, T.; Dor, I.; Prell, A.; Dembitsky, V.M. Fatty acid composition of six freshwater wild cyanobacterial species. Folia Microbiol. 2003, 48, 71–75. [Google Scholar] [CrossRef]
- Shin, K.; Jang, M.C.; Jang, P.K.; Ju, S.J.; Lee, T.K.; Chang, M. Influence of food quality on egg production and viability of the marine planktonic copepod Acartia omorii. Prog. Oceanogr. 2003, 57, 265–277. [Google Scholar] [CrossRef]
- Arendt, K.E.; Jónasdóttir, S.H.; Hansen, P.J.; Gärtner, S. Effects of dietary fatty acids on the reproductive success of the calanoid copepod Temora longicornis. Mar. Biol. 2005, 146, 513–530. [Google Scholar] [CrossRef]
- Thor, P.; Koski, M.; Tang, K.W.; Jónasdóttir, S.H. Supplemental effects of diet mixing on absorption of ingested organic carbon in the marine copepod Acartia tonsa. Mar. Ecol. Prog. Ser. 2007, 331, 131–138. [Google Scholar] [CrossRef]
- Dutz, J.; Koski, M.; Jónasdóttir, S.H. Copepod reproduction is unaffected by diatom aldehydes or lipid composition. Limnol. Oceanogr. 2008, 53, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Liu, H.; Chen, B. Effects of dietary essential fatty acids on reproduction rates of a subtropical calanoid copepod, Acartia erythraea. Mar. Ecol. Prog. Ser. 2012, 455, 95–110. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H.; Kiørboe, T. Copepod recruitment and food composition: Do diatoms affect hatching success? Mar. Biol. 1996, 125, 743–750. [Google Scholar] [CrossRef]
- Mansour, M.P.; Volkman, J.K.; Jackson, A.E.; Blackburn, S.I. The fatty acid and sterol composition of five marine dinoflagellates. J. Phycol. 1999, 35, 710–720. [Google Scholar] [CrossRef]
- Peltomaa, E.; Johnson, M.D.; Taipale, S.J. Marine cryptophytes are great sources of EPA and DHA. Mar. Drugs 2017, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Jónasdóttir, S.H.; Fields, D.; Pantoja, S. Copepod egg production in Long Island Sound, USA, as a function of the chemical composition of seston. Mar. Ecol. Prog. Ser. 1995, 119, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Md Amin, R.; Koski, M.; Båmstedt, U.; Vidoudez, C. Strain-related physiological and behavioral effects of Skeletonema marinoi on three common planktonic copepods. Mar. Biol. 2011, 158, 1965–1980. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Dutz, J.; Hagen, W. Role of essential fatty acids on the reproductive success of the copepod Temora longicornis in the North Sea. Mar. Ecol. Prog. Ser. 2007, 341, 153–163. [Google Scholar] [CrossRef]
- Mayzaud, P.; Chanut, J.P.; Ackman, R.G. Seasonal changes of the biochemical composition of marine particulate matter with special reference to fatty acids and sterols. Mar. Ecol. Prog. Ser. 1989, 56, 189–204. [Google Scholar] [CrossRef]
- Morris, R.J.; McCartney, M.J.; Robinson, G.A. Studies of a spring phytoplankton bloom in an enclosed experimental ecosystem. I. Biochemical changes in relation to the nutrient chemistry of water. J. Exp. Mar. Biol. Ecol. 1983, 70, 249–262. [Google Scholar] [CrossRef]
- Kattner, G.; Gercken, G.; Eberlein, K. Development of lipids during a spring plankton bloom in the northern North Sea. I. Particulate fatty acids. Mar. Chem. 1983, 14, 149–162. [Google Scholar] [CrossRef]
- Parrish, C.C. Time series of particulate and dissolved lipid classes during spring phytoplankton blooms in Bedford Basin, a marine inlet. Mar. Ecol. Prog. Ser. 1987, 35, 129–139. [Google Scholar] [CrossRef]
- Parrish, C.C.; Wangersky, P. Particulate and dissolved lipid classes in cultures of Phaeodactylum tricornutum grown in cage culture turbidostats with a range of nitrogen supply rates. Mar. Ecol. Prog. Ser. 1987, 35, 119–128. [Google Scholar] [CrossRef]
- Webb, K.; Chu, F.-L. Phytoplankton as a food source for bivalve larvae. In Proceedings of the Second International Conference on Aquaculture Nutrition: Biochemical and Physiological Approaches to Shellfish Nutrition; Pruder, G., Langdon, C., Conklin, D., Eds.; Louisiana State University: Baton Rouge, LA, USA, 1982; pp. 272–291. [Google Scholar]
- Cohen, Z. Products from microalgae. In Handbook for Micralgal Mass Culture.; Richmond, A., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1986; pp. 421–454. [Google Scholar]
- Thompson, P.A.; Guo, M.; Harrison, P.J. Effects of variation in temperature. I. On the biochemical composition of eight species of marine phytoplantkton. J. Phycol. 1992, 28, 481–488. [Google Scholar] [CrossRef]
- Rousch, J.M.M.; Bingham, S.E.E.; Sommerfeld, M.R.R. Changes in fatty acid profiles of thermo-intolerant and thermo-tolerant marine diatoms during temperature stress. J. Exp. Mar. Biol. Ecol. 2003, 295, 145–156. [Google Scholar] [CrossRef]
- Flaim, G.; Obertegger, U.; Guella, G. Changes in galactolipid composition of the cold freshwater dinoflagellate Borghiella dodgei in response to temperature. Hydrobiologia 2012, 698, 285–293. [Google Scholar] [CrossRef]
- Guihéneuf, F.; Mimouni, V.; Tremblin, G.; Ulmann, L. Light intensity regulates LC-PUFA incorporation into lipids of Pavlova lutheri and the final desaturase and elongase activities involved in their biosynthesis. J. Agric. Food Chem. 2015, 63, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Harrison, P.J.; Whyte, J.N.C. Influence of irradiance on the fatty acid composition of phytoplankton. J. Phycol. 1990, 26, 278–288. [Google Scholar] [CrossRef]
- Wainman, B.C.; Smith, R.E.; Rai, H.; Furgal, J.A. Irradiance and lipid production in natural algal populations. In Lipids in Freshwater Ecosystems; Arts, M.T., Wainmann, B.C., Eds.; Springer: New York, NY, USA, 1999; pp. 45–70. [Google Scholar]
- Shifrin, N.S.; Chisholm, S.W. Phytoplankton lipids: Interspecific differences and effects of nitrate, silicate and light-drak cycles. J. Phycol. 1981, 17, 374–384. [Google Scholar] [CrossRef]
- Groeger, A.; Schram, M.; Marzolf, G. Influence of food quality on growth and reproduction in Daphnia. Freshw. Biol. 1991, 26, 11–19. [Google Scholar] [CrossRef]
- Klein Breteler, W.; Schogt, N.; Rampen, S. Effect of diatom nutrient limitation on copepod development: The role of essential lipids. Mar. Ecol. Prog. Ser. 2005, 291, 125–133. [Google Scholar] [CrossRef]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of Nutrient Limitation on Fatty-Acid and Lipid-Content of Marine Microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Dörmann, P. Galactolipids in plant membranes. In Encyclopedia of Life Sciences: Els; John Wiley & Sons Ltd.: Chichester, UK, 2001; Available online: http://www.els.net (accessed on 31 August 2015).
- Taguchi, S.; Hirata, J.A.; Laws, E.A. Silicate Deficiency and Lipid Synthesis of Marine Diatoms. J. Phycol. 1987, 23, 260–267. [Google Scholar] [CrossRef]
- Roessler, P.G. Effects of silicon deficiency on lipid composition and metabolism in the diatom Cyclotella cryptica. J. Phycol. 1988, 24, 394–400. [Google Scholar] [CrossRef]
- Gavalas, N.; Clark, H. On the Role of Manganese in Photosynthesis Kinetics of Photoinhibition in Manganese-deficent and 3-(4-Chlorophenyl)-1, 1-dimethylurea-inhibited Euglena gracilis. Plant Physiol. 1971, 47, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Shanklin, J.; Cahoon, E.B. Desaturation and related modifications of fatty acids. Annu. Rev. Plant Biol. 1998, 49, 611–641. [Google Scholar] [CrossRef] [PubMed]
- Los, D.A.; Murata, N. Structure and expression of fatty acid desaturases. Biochim. Biophys. Acta 1998, 1394, 3–15. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef] [PubMed]
- Pike, I.H.; Jackson, A. Fish oil: Production and use now and in the future. Lipid Technol. 2010, 22, 59–61. [Google Scholar] [CrossRef]
- Greenberg, P. The Omega Principle: Seafood and the Quest for a Long Life and a Healthier Planet; Penguin Books: London, UK, 2018. [Google Scholar]
- Calder, P.C. Mechanisms of Action of (n-3) Fatty Acids, 2. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed]
- IPCC Climate Change 2014 Synthesis Report Summary Chapter for Policymakers; IPCC: Geneva, Switzerland, 2014; p. 32.
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland Galway: Galway, Ireland, 2018; Available online: http://www.algaebase.org (accessed on 20 December 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phyla | Total FA | PUFA | HUFA | HUFA + 18:5 | ω3/ω6 | DHA/EPA |
|---|---|---|---|---|---|---|
| Chlorophyta | 8.6 ± 5.1 | 5.3 ± 3.6 | 0.3 ± 0.4 | 0.4 ± 0.5 | 8.1 ± 13.1 | 3.1 ± 4.8 |
| Diatoma | 13.6 ± 18.1 | 4.5 ± 6.0 | 2.3 ± 3.1 | 2.3 ± 3.1 | 15.2 ± 17.5 | 9.2 ± 9.0 |
| Dinophyta | 5.5 ± 2.2 | 1.6 ± 0.9 | 0.6 ± 0.5 | 1.1 ± 0.8 | 23.1 ± 15.8 | 0.3 ± 0.3 |
| Haptophyta | 14 ± 3.7 | 5.5 ± 2.2 | 2.5 ± 2.3 | 2.9 ± 2.0 | 5.0 ± 5.4 | 1.2 ± 3.2 |
| Cyanobacteria | ? | 0 | 0 | 0 | 2.5 ± 2.3 | - |
| Cryptophyta | 9.4 ± 7.7 | 5.9 ± 5.3 | 1.8 ± 1.2 | 1.8 ± 1.2 | 7.0 ± 7.3 | 1.8 ± 1.1 |
| Ochrophyta | 13.1 | 4.5 | 3.9 | 3.9 | 5.4 | 0 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jónasdóttir, S.H. Fatty Acid Profiles and Production in Marine Phytoplankton. Mar. Drugs 2019, 17, 151. https://doi.org/10.3390/md17030151
Jónasdóttir SH. Fatty Acid Profiles and Production in Marine Phytoplankton. Marine Drugs. 2019; 17(3):151. https://doi.org/10.3390/md17030151
Chicago/Turabian StyleJónasdóttir, Sigrún Huld. 2019. "Fatty Acid Profiles and Production in Marine Phytoplankton" Marine Drugs 17, no. 3: 151. https://doi.org/10.3390/md17030151
APA StyleJónasdóttir, S. H. (2019). Fatty Acid Profiles and Production in Marine Phytoplankton. Marine Drugs, 17(3), 151. https://doi.org/10.3390/md17030151