Marine Fungi from the Sponge Grantia compressa: Biodiversity, Chemodiversity, and Biotechnological Potential




 , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Materials and Fungal Isolation
2.2. Fungal Identification
2.3. Small-Scale Fermentation and OSMAC Approach
2.3.1. Multiwell Culture and Co-Culture Conditions
2.3.2. Extraction Procedure and Chemical Analyses
2.4. Scale-Up of Eurotium chevalieri MUT 2316
2.4.1. Solid Media Culture Condition and Secondary Metabolite Extraction from Eurotium chevalieri MUT 2316
2.4.2. Liquid Media Culture Condition and Secondary Metabolite Extraction from Eurotium chevalieri MUT 2316
2.5. High-Resolution Mass Spectrometry Analysis and Molecular Networking Analyses
2.6. Antiviral Assays
2.6.1. Cells and Viruses
2.6.2. Cytotoxicity Assays
2.6.3. Plaque Reduction Assay
2.7. Antibacterial Assays
2.7.1. Bacterial Growth Conditions
2.7.2. Minimal Inhibitory Concentration (MIC) Determination
2.7.3. Minimal Bactericidal Concentration (MBC) Determination
3. Results and Discussion
3.1. The Mycobiota of Grantia compressa
3.2. OSMAC Approach
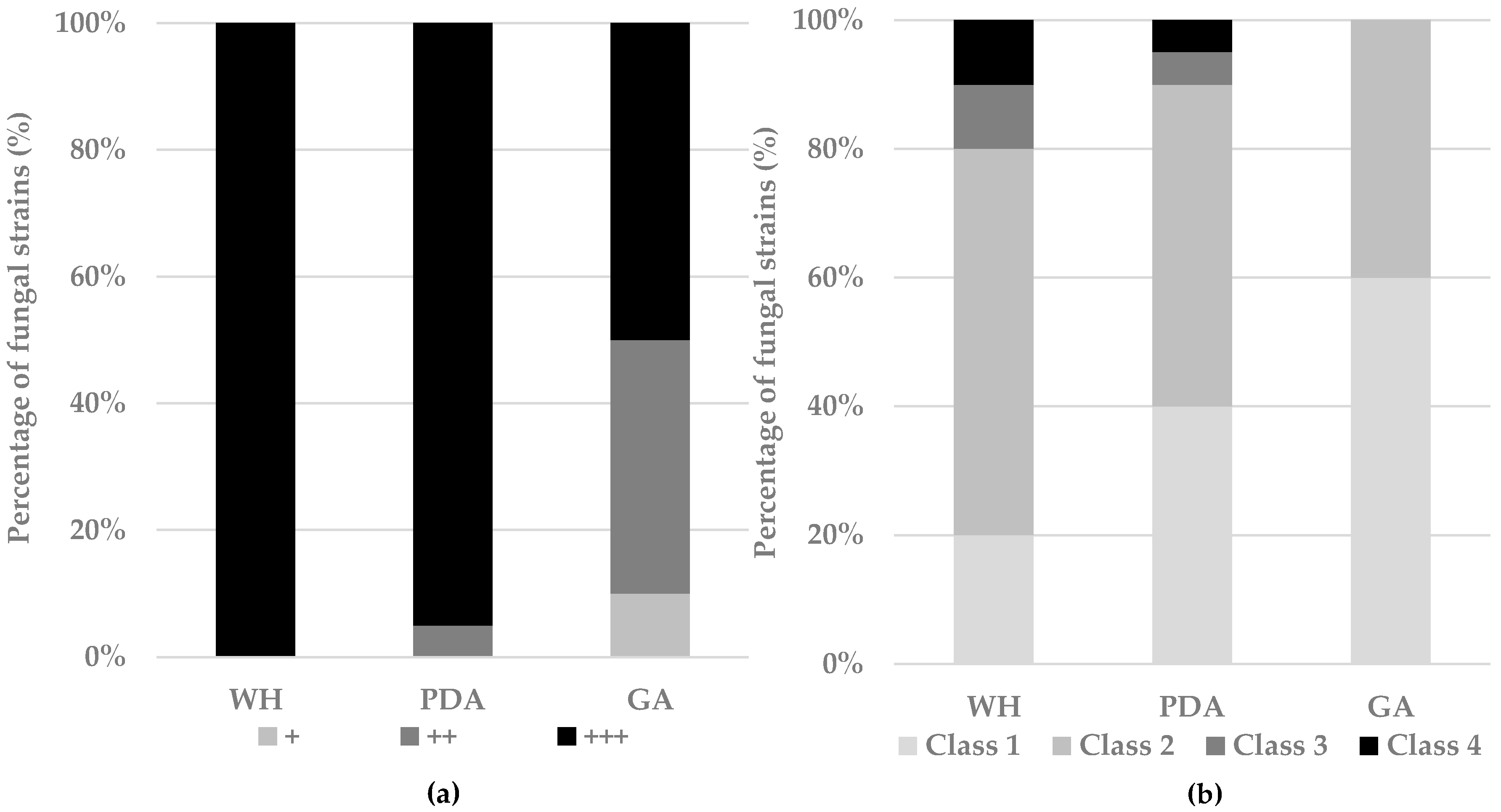
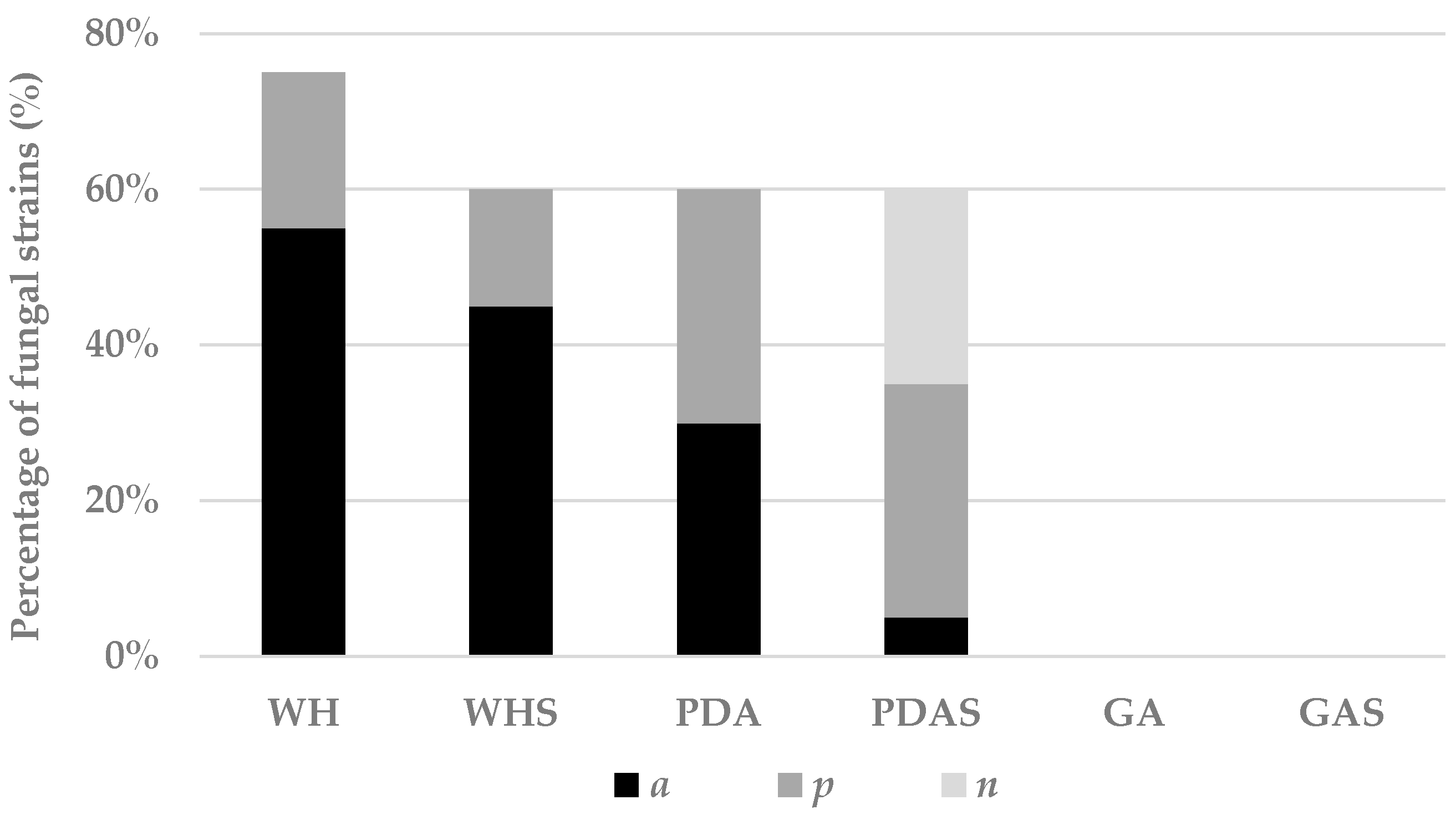
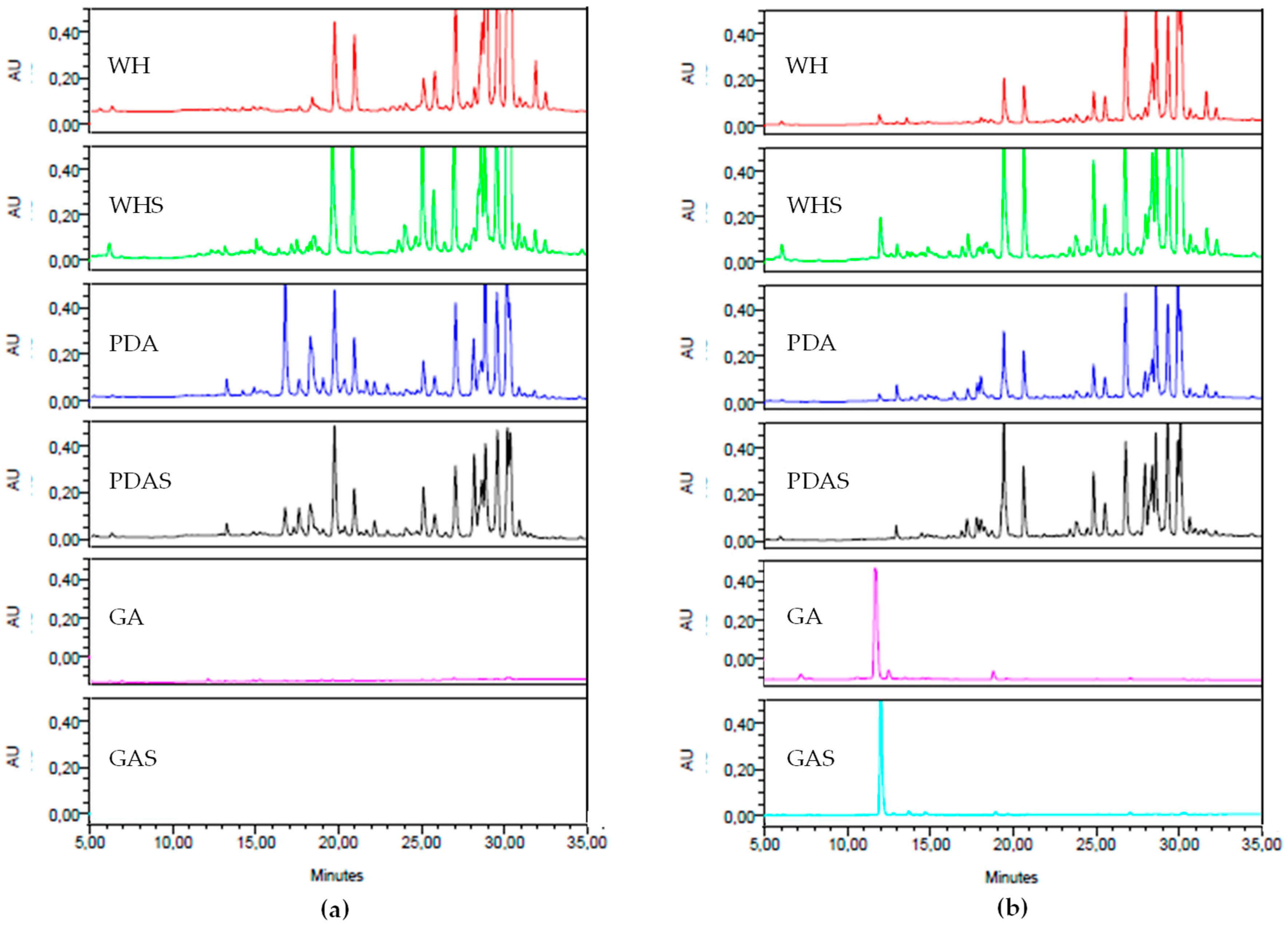
3.2.1. Effect of Media on Fungal Development and Chemical Fingerprint
3.2.2. Effect of Salt on Fungal Development and Chemical Fingerprint
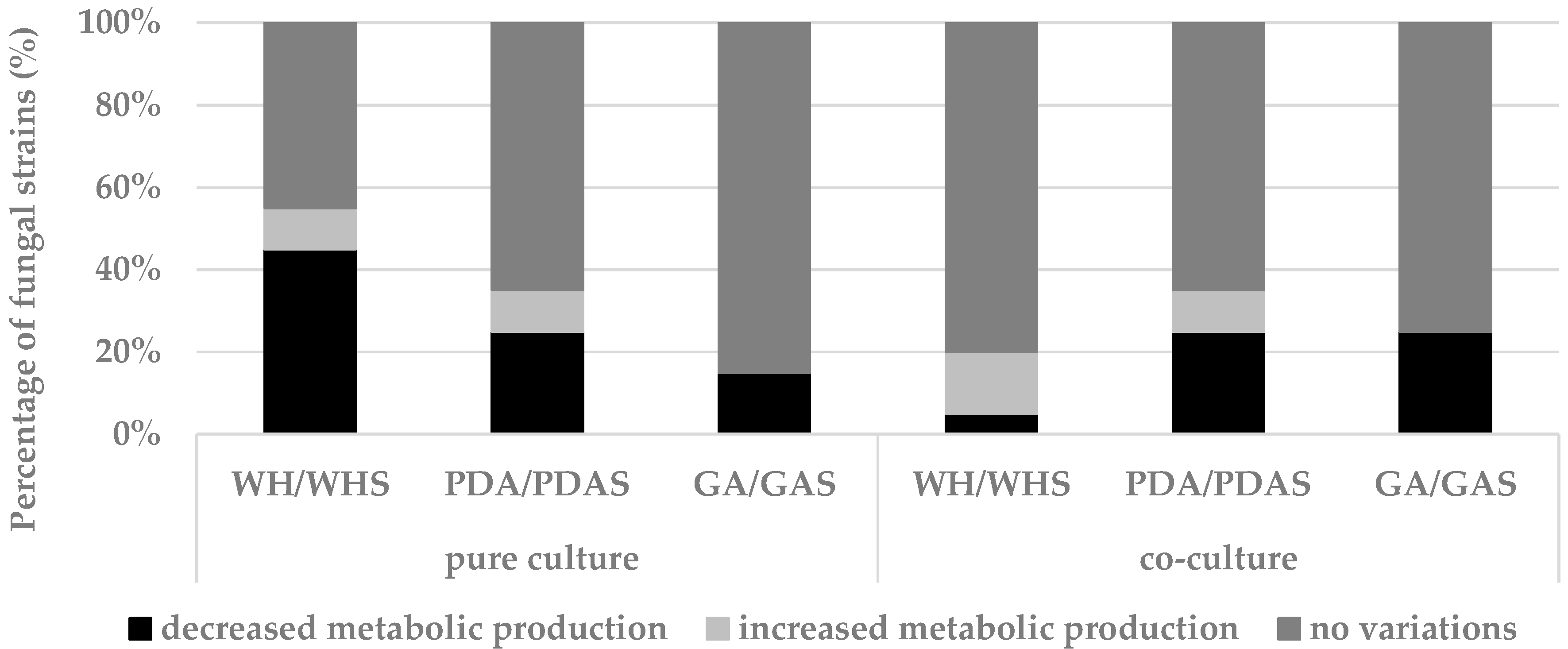
3.2.3. Effect of Co-Culture on Fungal Development and Chemical Fingerprint
3.2.4. General Remarks on the OSMAC Approach
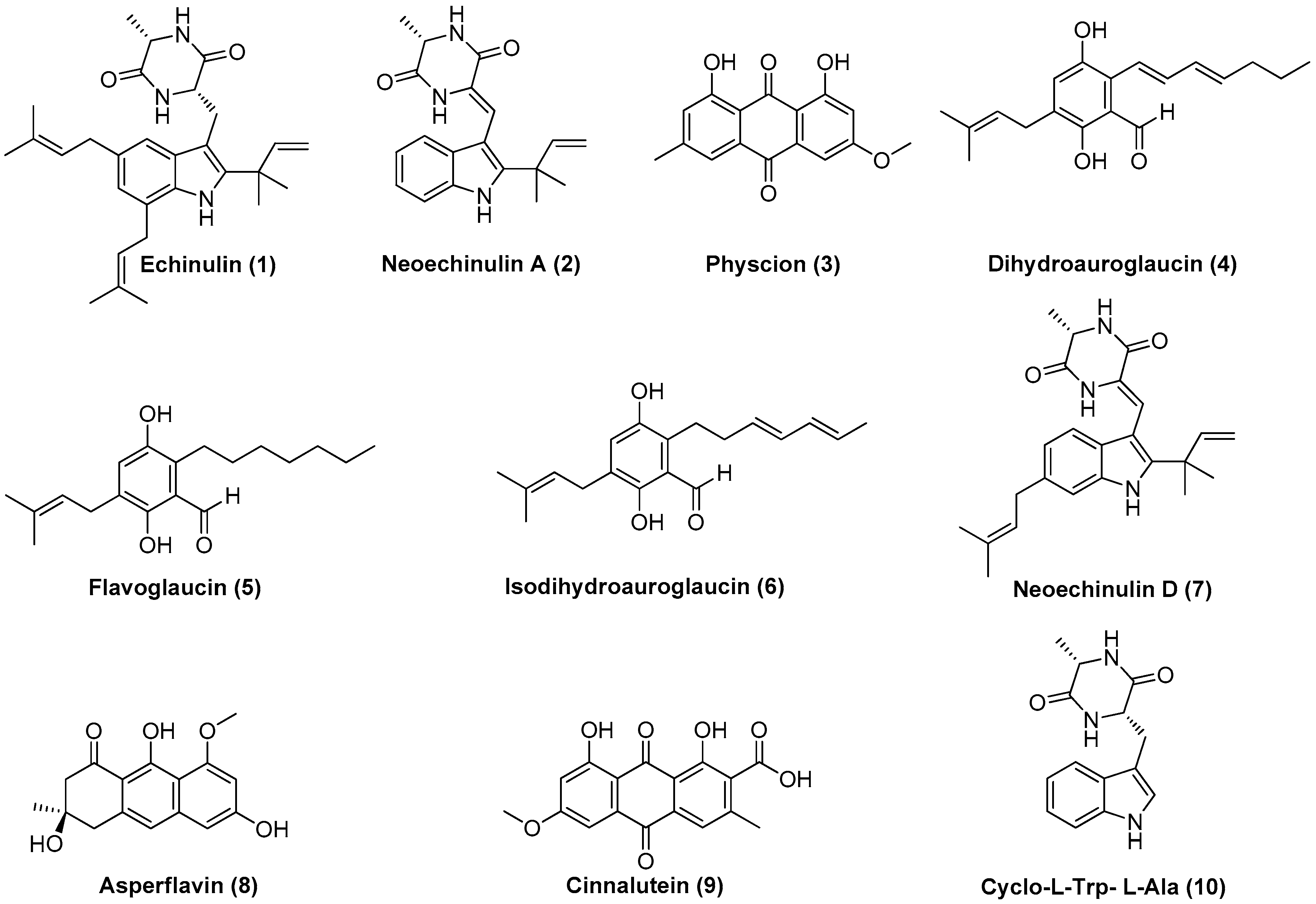
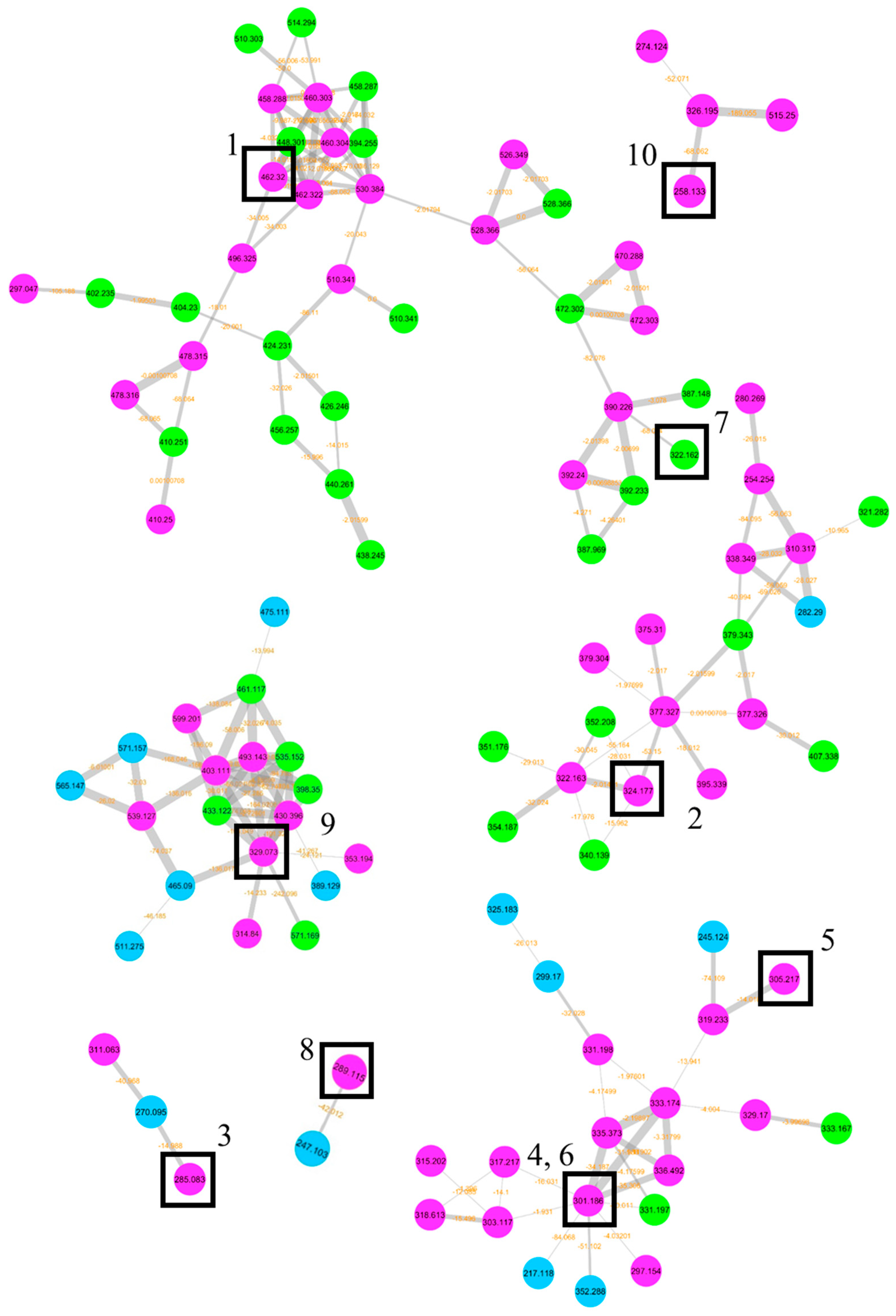
3.3. Eurotium chevalieri MUT 2316: Molecular Networking and Compounds
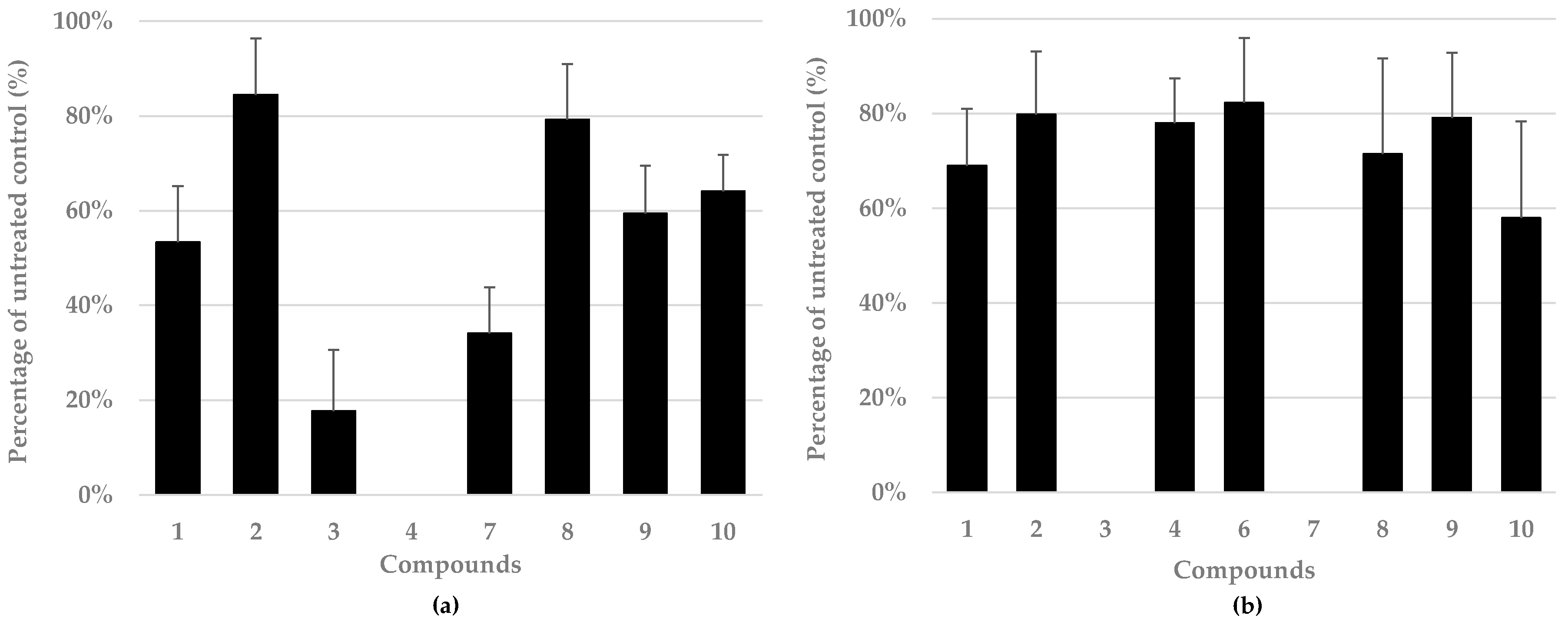
3.4. Eurotium chevalieri-Derived Compounds Show Antiviral Activity against Herpes and Influenza Viruses
3.5. Eurotium chevalieri-Derived Compounds Show Antibacterial Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Adpressa, D.A.; Loesgen, S. Bioprospecting chemical diversity and bioactivity in a marine derived Aspergillus terreus. Chem. Biodivers. 2016, 13, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.; Carroll, A.; Copp, B.; Davis, R.; Keyzers, R.; Prinsep, M. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proksch, P.; Ebel, R.; Edrada, R.A.; Schupp, P.; Lin, W.H.; Wray, V.; Steube, K. Detection of pharmacologically active natural products using ecology. Selected examples from Indopacific marine invertebrates and sponge-derived fungi. Pure Appl. Chem. 2003, 75, 343–352. [Google Scholar] [CrossRef]
- Huryn, D.M.; Wipf, P. Natural product chemistry and cancer drug discovery. In Cancer Drug Design and Discovery, 2nd ed.; Neidle, S., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 91–120. ISBN 978-0-12-396521-9. [Google Scholar]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- González-Menéndez, V.; Pérez-Bonilla, M.; Pérez-Victoria, I.; Martín, J.; Muñoz, F.; Reyes, F.; Tormo, J.R.; Genilloud, O. Multicomponent analysis of the differential induction of secondary metabolite profiles in fungal endophytes. Molecules 2016, 21, 234. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.; Patry, S.; Dobson, A. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yu, H.; Li, R.; Xing, R.; Liu, S.; Li, P. Exploring the antibacterial and antifungal potential of jellyfish-associated marine fungi by cultivation-dependent approaches. PLoS ONE 2015, 10, e0144394. [Google Scholar] [CrossRef] [PubMed]
- VanderMolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. AMB Express 2013, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Platas, G.; Fillola, A.; Jimenez, M.R.; Collado, J.; Vicente, F.; Martín, J.; González, J.; Bur-Zimmermann, J.; Tormo, J.R.; et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallet, M.; Vanbellingen, Q.P.; Fu, T.; Le Caer, J.P.; Della-Negra, S.; Touboul, D.; Duncan, K.R.; Nay, B.; Brunelle, A.; Prado, S. An integrative approach to decipher the chemical antagonism between the competing endophytes Paraconiothyrium variabile and Bacillus subtilis. J. Nat. Prod. 2017, 80, 2863–2873. [Google Scholar] [CrossRef]
- Takahashi, J.A.; Teles, A.P.C.; Bracarense, A.D.A.P.; Gomes, D.C. Classical and epigenetic approaches to metabolite diversification in filamentous fungi. Phytochem. Rev. 2013, 12, 773–789. [Google Scholar] [CrossRef]
- Jones, E.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.L. Classification of marine ascomycota, basidiomycota, blastocladiomycota and chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Pang, K.L.; Jones, E.G. Recent advances in marine mycology. Bot. Mar. 2017, 60, 361–362. [Google Scholar] [CrossRef] [Green Version]
- Garzoli, L.; Poli, A.; Prigione, V.; Gnavi, G.; Varese, G.C. Peacock’s tail with a fungal cocktail: First assessment of the mycobiota associated with the brown alga Padina pavonica. Fungal Ecol. 2018, 35, 87–97. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Bovio, E.; Garzoli, L.; Poli, A.; Prigione, V.; Firsova, D.; McCormack, G.P.; Varese, G.C. The culturable mycobiota associated with three Atlantic sponges, including two new species: Thelebolus balaustiformis and T. spongiae. Fungal Syst. Evol. 2018, 1, 141–167. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S. Fungal endosymbionts of seaweeds. In Biology of Marine Fungi, 1st ed.; Se-Kwon, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 53–69. ISBN 978-3-642-53970-1. [Google Scholar]
- Zin, W.W.M.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inacio, A.; Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.S.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 06. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi; Academic Press: London, UK, 1980; Volume 1. [Google Scholar]
- Von Arx, J.A. The Genera of Fungi Sporulating in Pure Culture, 3rd ed.; Cramer: Vaduz, Liechtenstein, 1981. [Google Scholar]
- Kiffer, E.; Morelet, M. Les deutéromycetes: Classification et clés D’identification Genérique; INRA: Paris, France, 1997. [Google Scholar]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Skidmore, A.M.; Dickinson, C.H. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans. Br. Mycol. Soc. 1976, 66, 57–64. [Google Scholar] [CrossRef]
- Liang, T.M.; Fang, Y.W.; Zheng, J.Y.; Shao, C.L. Secondary metabolites isolated from the gorgonian-derived fungus Aspergillus ruber and their antiviral activity. Chem. Nat. Compd. 2018, 54, 559–561. [Google Scholar] [CrossRef]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Farrah, T.; Lam, H.; Tasman, N.; Sun, Z.; Nilsson, E.; Pratt, B.; Prazen, B.; et al. A guided tour of the Trans-Proteomic Pipeline. Proteomics 2010, 10, 1150–1159. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–838. [Google Scholar] [CrossRef]
- Luganini, A.; Nicoletto, S.F.; Pizzuto, L.; Pirri, G.; Giuliani, A.; Landolfo, S.; Gribaudo, G. Inhibition of herpes simplex virus type 1 and type 2 infections by peptide-derivatized dendrimers. Antimicrob. Agents Chemother. 2011, 55, 3231–3239. [Google Scholar] [CrossRef]
- Luganini, A.; Terlizzi, M.E.; Catucci, G.; Gilardi, G.; Maffei, M.E.; Gribaudo, G. The cranberry extract Oximacro® exerts in vitro virucidal activity against Influenza virus by interfering with hemagglutinin. Front. Microbiol. 2018, 9, 1826–1838. [Google Scholar] [CrossRef]
- CLSI. Methods for Diluition Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 10th ed.; CLSI Document M07-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Zhao, Y.; Li, S.Q.; Li, H.J.; Lan, W.J. Lanostane triterpenoids from the fungus Ceriporia lacerate associated with Acanthaster planci. Chem. Nat. Compd. 2013, 49, 653–656. [Google Scholar] [CrossRef]
- Zhang, L.; Kang, M.; Huang, Y.; Yang, L. Fungal communities from the calcareous deep-sea sediments in the Southwest India Ridge revealed by Illumina sequencing technology. World J. Microb. Biot. 2016, 32, 1–11. [Google Scholar] [CrossRef]
- Gnavi, G.; Garzoli, L.; Poli, A.; Prigione, V.; Burgaud, G.; Varese, G.C. The culturable mycobiota of Flabellia petiolata: First survey of marine fungi associated to a Mediterranean green alga. PLoS ONE 2017, 12, e0175941. [Google Scholar] [CrossRef]
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 2013, 30, 685–694. [Google Scholar] [CrossRef]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisafi, F.; Varese, G.C. The culturable mycobiota of a Mediterranean marine site after an oil spill: Isolation, identification and potential application in bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef]
- Rämä, T.; Hassett, B.T.; Bubnova, E. Arctic marine fungi: From filaments and flagella to operational taxonomic units and beyond. Bot. Mar. 2017, 60, 433–452. [Google Scholar] [CrossRef]
- Oren, A.; Gunde-Cimerman, N. Fungal life in the Dead Sea. In Biology of Marine Fungi, 1st ed.; Se-Kwon, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 115–132. ISBN 978-3-642-53970-1. [Google Scholar]
- Raghukumar, C.; Ravindran, J. Fungi and their role in corals and coral reef ecosystems. In Biology of Marine Fungi, 1st ed.; Se-Kwon, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 89–113. ISBN 978-3-642-53970-1. [Google Scholar]
- Zajc, J.; Zalar, P.; Plemenitaš, A.; Gunde-Cimerman, N. The mycobiota of the salterns. In Biology of Marine Fungi, 1st ed.; Se-Kwon, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 133–158. ISBN 978-3-642-53970-1. [Google Scholar]
- Henríquez, M.; Vergara, K.; Norambuena, J.; Beiza, A.; Maza, F.; Ubilla, P.; Araya, I.; Chávez, R.; San-Martín, A.; Darias, J.; et al. Diversity of cultivable fungi associated with Antarctic marine sponges and screening for their antimicrobial, antitumoral and antioxidant potential. World J. Microb. Biot. 2014, 30, 65–76. [Google Scholar] [CrossRef]
- Rozas, E.E.; Albano, R.M.; Lôbo-Hajdu, G.; Müller, W.E.; Schröder, H.C.; Custódio, M.R. Isolation and cultivation of fungal strains from in vitro cell cultures of two marine sponges (Porifera: Halichondrida and Haplosclerida). Braz. J. Microbiol. 2011, 42, 1560–1568. [Google Scholar] [CrossRef]
- Manriquez, V.; Galdamez, A.; Veliz, B.; Rovirosa, J.; Diaz-Marrero, A.R.; Cueto, M.; Daria, J.; Martínez, C.; San-Martín, A. N-methyl-1H-indole-2-carboxamide from the marine fungus Cladosporium cladosporioides. J. Chil. Chem. Soc. 2009, 54, 314–316. [Google Scholar] [CrossRef]
- San-Martin, A.; Painemal, K.; Díaz, Y.; Martinez, C.; Rovirosa, J. Metabolites from the marine fungus Cladosporium cladosporioides. J. Argent. Chem Soc 2005, 93, 247–251. [Google Scholar]
- Pivkin, M.V.; Aleshko, S.A.; Krasokhin, V.B.; Khudyakova, Y.V. Fungal assemblages associated with sponges of the southern coast of Sakhalin Island. Russ. J. Mar. Biol. 2006, 32, 207–213. [Google Scholar] [CrossRef]
- Sayed, M.A.E.; El-Rahman, T.M.A.; El-Diwany, A.I.; Sayed, S.M. Biodiversity and bioactivity of red sea sponge associated endophytic fungi. IJAAS 2016, 5, 1–15. [Google Scholar]
- Gomes, D.; Cavalcanti, M.A.Q.; Fernandes, M.J.S.; Lima, D.M.M.; Passavante, J.Z.O. Filamentous fungi isolated from sand and water of “Bairro Novo” and ”Casa Caiada” beaches, Olinda, Pernambuco, Brazil. Braz. J. Biol. 2008, 68, 577–582. [Google Scholar] [CrossRef]
- Butinar, L.; Zalar, P.; Frisvad, J.C.; Gunde-Cimerman, N. The genus Eurotium–members of indigenous fungal community in hypersaline waters of salterns. FEMS Microbiol. Ecol. 2005, 51, 155–166. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Lopez-Gresa, M.P.; Gavagnin, M.; Nicoletti, R.; Manzo, E.; Mollo, E.; Guo, Y.W.; Cimino, G. Cytosporin-related compounds from the marine-derived fungus Eutypella scoparia. Tetrahedron 2008, 64, 5365–5369. [Google Scholar] [CrossRef]
- Hatai, K. Diseases of fish and shellfish caused by marine fungi. In Biology of Marine Fungi, 1st ed.; Se-Kwon, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 15–52. ISBN 978-3-642-53970-1. [Google Scholar]
- Raghukumar, S. Fungi in Coastal and Oceanic Marine Ecosystems, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–364. ISBN 978-3-319-54303-1. [Google Scholar]
- Paz, Z.; Komon-Zelazowska, M.; Druzhinina, I.S.; Aveskamp, M.M.; Shnaiderman, A.; Aluma, Y.; Carmeli, S.; Ilan, M.; Yarden, O. Diversity and potential antifungal properties of fungi associated with a Mediterranean sponge. Fungal Divers. 2010, 42, 17–26. [Google Scholar] [CrossRef]
- Bolaños, J.; De León, L.F.; Ochoa, E.; Darias, J.; Raja, H.A.; Shearer, C.A.; Miller, A.N.; Vanderheyden, P.; Porras-Alfaro, A.; Caballero-George, C. Phylogenetic diversity of sponge-associated fungi from the Caribbean and the Pacific of Panama and their in vitro effect on angiotensin and endothelin receptors. Mar. Biotechnol. 2015, 17, 533–564. [Google Scholar] [CrossRef]
- Passarini, M.R.; Santos, C.; Lima, N.; Berlinck, R.G.; Sette, L.D. Filamentous fungi from the Atlantic marine sponge Dragmacidon reticulatum. Arch. Microbiol. 2013, 195, 99–111. [Google Scholar] [CrossRef]
- Wiese, J.; Ohlendorf, B.; Blümel, M.; Schmaljohann, R.; Imhoff, J.F. Phylogenetic identification of fungi isolated from the marine sponge Tethya aurantium and identification of their secondary metabolites. Mar. Drugs 2011, 9, 561–585. [Google Scholar] [CrossRef]
- Sun, Y.L.; He, F.; Liu, K.S.; Zhang, X.Y.; Bao, J.; Wang, Y.F.; Nong, X.H.; Qi, S.H. Cytotoxic dihydrothiophene-condensed chromones from marine-derived fungus Penicillium oxalicum. Planta Med. 2013, 78, 1957–1961. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.C.; Li, Y.; Kim, E.A.; Kang, S.M.; Jeon, Y.J. Asperflavin, an anti-inflammatory compound produced by a marine-derived fungus, Eurotium amstelodami. Molecules 2017, 22, 1823. [Google Scholar] [CrossRef]
- Alwakeel, S.S. Isolation and Identification of Fungal and Bacterial Specimens from the Sand and Seawater of the Red Sea Coastline of Saudi Arabia. Adv. Environ. Biol. 2013, 7, 1366–1373. [Google Scholar]
- Gao, Z.; Li, B.; Zheng, C.; Wang, G. Molecular detection of fungal communities in the Hawaiian marine sponges Suberites zeteki and Mycale armata. Appl. Environ. Microb. 2008, 74, 6091–6101. [Google Scholar] [CrossRef]
- Li, C.S.; Li, X.M.; Gao, S.S.; Lu, Y.H.; Wang, B.G. Cytotoxic anthranilic acid derivatives from deep sea sediment-derived fungus Penicillium paneum SD-44. Mar. Drugs 2013, 11, 3068–3076. [Google Scholar] [CrossRef]
- Takami, H. Isolation and characterization of microorganisms from deep-sea mud. In Extremophiles in Deep-Sea Environments, 1st ed.; Horikoshi, K., Tsujii, K., Eds.; Springer: Tokyo, Japan, 1999; pp. 3–26. ISBN 978-4-431-68008-6. [Google Scholar]
- Kachalkin, A.V. Yeasts of the White Sea intertidal zone and description of Glaciozyma litorale sp. nov. Antonie Van Leeuw. J. Microb. 2014, 105, 1073–1083. [Google Scholar] [CrossRef]
- Rämä, T.; Nordén, J.; Davey, M.L.; Mathiassen, G.H.; Spatafora, J.W.; Kauserud, H. Fungi ahoy! Diversity on marine wooden substrata in the high North. Fungal Ecol. 2014, 8, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Yin, Y.; Zhang, F.; Li, Z. Recovery and phylogenetic diversity of culturable fungi associated with marine sponges Clathrina luteoculcitella and Holoxea sp. in the South China Sea. Mar. Biotechnol. 2011, 13, 713–721. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G. Diversity of fungal isolates from three Hawaiian marine sponges. Microbiol. Res. 2009, 164, 233–241. [Google Scholar] [CrossRef]
- Höller, U.; Wright, A.D.; Matthee, G.F.; Konig, G.M.; Draeger, S.; Aust, H.; Schulz, B. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol. Res. 2000, 104, 1354–1365. [Google Scholar] [CrossRef]
- Proksch, P.; Ebel, R.; Edrada, R.; Riebe, F.; Liu, H.; Diesel, A.; Bayer, M.; Li, X.; Lin, W.H.; Grebenyuk, V.; et al. Sponge-associated fungi and their bioactive compounds: The Suberites case. Bot. Mar. 2008, 51, 209–218. [Google Scholar] [CrossRef]
- Thirunavukkarasu, N.; Suryanarayanan, T.S.; Girivasan, K.P.; Venkatachalam, A.; Geetha, V.; Ravishankar, J.P.; Doble, M. Fungal symbionts of marine sponges from Rameswaram, southern India: Species composition and bioactive metabolites. Fungal Divers. 2012, 55, 37–46. [Google Scholar] [CrossRef]
- Wang, G.; Li, Q.; Zhu, P. Phylogenetic diversity of culturable fungi associated with the Hawaiian sponges Suberites zeteki and Gelliodes fibrosa. Antonie Van Leeuw. J. Microb. 2008, 93, 163–174. [Google Scholar] [CrossRef]
- Reich, M.; Labes, A. How to boost marine fungal research: A first step towards a multidisciplinary approach by combining molecular fungal ecology and natural products chemistry. Mar. Genom. 2017, 36, 57–75. [Google Scholar] [CrossRef]
- Imhoff, J.F. Natural products from marine fungi—Still an underrepresented resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef]
- Poli, A.; Vizzini, A.; Prigione, V.; Varese, G.C. Basidiomycota isolated from the Mediterranean Sea–Phylogeny and putative ecological roles. Fungal Ecol. 2018, 36, 51–62. [Google Scholar] [CrossRef]
- Baker, P.W.; Kennedy, J.; Dobson, A.D.; Marchesi, J.R. Phylogenetic diversity and antimicrobial activities of fungi associated with Haliclona simulans isolated from Irish coastal waters. Mar. Biotechnol. 2009, 11, 540–547. [Google Scholar] [CrossRef]
- Macintyre, L.; Zhang, T.; Viegelmann, C.; Martinez, I.J.; Cheng, C.; Dowdells, C.; Abdelmohsen, U.R.; Gernert, C.; Hentschel, U.; Edrada-Ebel, R. Metabolomic tools for secondary metabolite discovery from marine microbial symbionts. Mar. Drugs 2014, 12, 3416–3448. [Google Scholar] [CrossRef]
- Linde, T.; Hansen, N.B.; Lübeck, M.; Lübeck, P.S. Fermentation in 24-well plates is an efficient screening platform for filamentous fungi. Lett. Appl. Microbiol. 2014, 59, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Proksch, P. Endophytes and associated marine derived fungi—Ecological and chemical perspectives. Fungal Divers. 2012, 57, 45–83. [Google Scholar] [CrossRef]
- Jančič, S.; Frisvad, J.C.; Kocev, D.; Gostinčar, C.; Džeroski, S.; Gunde-Cimerman, N. Production of secondary metabolites in extreme environments: Food-and airborne Wallemia spp. produce toxic metabolites at hypersaline conditions. PLoS ONE 2016, 11, e0169116. [Google Scholar] [CrossRef]
- Kalinina, S.A.; Jagels, A.; Cramer, B.; Geisen, R.; Humpf, H.U. Influence of environmental factors on the production of Penitrems A–F by Penicillium crustosum. Toxins 2017, 9, 210. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Sun, K.; Zhu, W. Effects of high salt stress on secondary metabolite production in the marine-derived fungus Spicaria elegans. Mar. Drugs 2011, 9, 535–542. [Google Scholar] [CrossRef]
- Shehata, H.R.; Lyons, E.M.; Jordan, K.S.; Raizada, M.N. Relevance of in vitro agar based screens to characterize the anti-fungal activities of bacterial endophyte communities. BMC Microbiol. 2016, 16, 8. [Google Scholar] [CrossRef]
- Chen, X.; Si, L.; Liu, D.; Proksch, P.; Zhang, L.; Zhou, D.; Lin, W. Neoechinulin B and its analogues as potential entry inhibitors of influenza viruses, targeting viral hemagglutinin. Eur. J. Med. Chem. 2015, 93, 182–195. [Google Scholar] [CrossRef]
- Gao, J.; Radwan, M.M.; León, F.; Wang, X.; Jacob, M.R.; Tekwani, B.L.; Khan, S.I.; Lupien, S.; Hill, R.A.; Dugan, F.M.; et al. Antimicrobial and antiprotozoal activities of secondary metabolites from the fungus Eurotium repens. Med. Chem. Res. 2012, 21, 3080–3086. [Google Scholar] [CrossRef]
- Ganihigama, D.U.; Sureram, S.; Sangher, S.; Hongmanee, P.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Antimycobacterial activity of natural products and synthetic agents: Pyrrolodiquinolines and vermelhotin as anti-tubercular leads against clinical multidrug resistant isolates of Mycobacterium tuberculosis. Eur. J. Med. Chem. 2015, 89, 1–12. [Google Scholar] [CrossRef]
- Basu, S.; Ghosh, A.; Hazra, B. Evaluation of the antibacterial activity of Ventilago madraspatana Gaertn., Rubia cordifolia Linn. and Lantana camara Linn.: Isolation of emodin and physcion as active antibacterial agents. Phytother. Res. 2005, 19, 888–894. [Google Scholar] [CrossRef]
- Tian, S.Z.; Pu, X.; Luo, G.; Zhao, L.X.; Xu, L.H.; Li, W.J.; Luo, Y. Isolation and characterization of new p-terphenyls with antifungal, antibacterial, and antioxidant activities from halophilic actinomycete Nocardiopsis gilva YIM 90087. J. Agric. Food Chem. 2013, 61, 3006–3012. [Google Scholar] [CrossRef]
- Pereira, F.; Madureira, A.M.; Sancha, S.; Mulhovo, S.; Luo, X.; Duarte, A.; Ferreira, M.J.U. Cleistochlamys kirkii chemical constituents: Antibacterial activity and synergistic effects against resistant Staphylococcus aureus strains. J. Ethnopharmacol. 2016, 178, 180–187. [Google Scholar] [CrossRef]
- Meng, L.H.; Du, F.Y.; Li, X.M.; Pedpradab, P.; Xu, G.M.; Wang, B.G. Rubrumazines A–C, indolediketopiperazines of the isoechinulin class from Eurotium rubrum MA-150, a fungus obtained from marine mangrove-derived rhizospheric soil. J. Nat. Prod. 2015, 78, 909–913. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MUT CODE | Taxa | RA (%) | Isolation Method | Isolation Media | Incubation Temperature | Report | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DP | HO | CMASW | GASW | MEA | SWA | 15 °C | 25 °C | MA | SP | |||
| 2282 | Coprinellus sp. * | 8.7 | x | x | x | - | - | |||||
| 2332 | x | x | x | |||||||||
| 2288 | Ceriporia lacerate * | 4.3 | x | x | x | [34,35] | FR | |||||
| 2307 | Cladosporium allicinum | 13.0 | x | x | x | [16,36] | [18] | |||||
| 2313 | x | x | x | |||||||||
| 2314 | Cladosporium cladosporioides | 4.3 | x | x | x | [15,16,36,37,38,39,40,41,42] | [18,43,44,45,46,47,48] | |||||
| 2315 | Cladosporium pseudocladosporioides | 8.7 | x | x | x | [16,38] | [18] | |||||
| 2273 | Emericellopsis pallida * | 8.7 | x | x | x | [15] | [18] | |||||
| 2274 | x | x | x | |||||||||
| 2316 | Eurotium chevalieri | 4.3 | x | x | x | [15,40,42,49,50] | [47] | |||||
| 2334 | Euthypella scoparia * | 4.3 | x | x | x | [51] | [8] | |||||
| 2317 | Fusarium solani | 4.3 | x | x | x | [52,53] | [54,55] | |||||
| 2321 | Penicillium chrysogenum | 4.3 | x | x | x | [15,38,39,42] | [47,54,56,57] | |||||
| 2328 | Penicillium oxalicum | 4.3 | x | x | x | [15,42,53,58,59,60] | [61] | |||||
| 2322 | Penicillium paneum | 8.7 | x | x | x | [15,62] | FR | |||||
| 2326 | x | x | x | |||||||||
| 2331 | Psathyrella candolleana * | 4.3 | x | x | x | FR | FR | |||||
| 2415 | Rhodotorula mucilaginosa | 4.3 | x | x | x | [15,63,64] | FR | |||||
| - | Tetracladium sp. * | 4.3 | x | x | x | - | - | |||||
| 2410 | Tolypocladium cylindrosporum | 8.7 | x | x | x | [39,53,65] | [18] | |||||
| 2413 | x | x | x | |||||||||
| Bacteria | Staphylococcus aureus | Enterococcus faecalis | Streptococcus pneumoniae | Pseudomonas aeruginosa | Escherichia coli | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compounds | ATCC 29213 | Monza-PFI | Monza-FD1 | ATCC 29212 | ATCC 49619 | Monza 82 | ATCC 27853 | ATCC 25922 | ||||||||
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| Echinulin (1) | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A |
| Neoechinulin A (2) | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A |
| Physcion (3) | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | 16 | >32 | >32 | N/A | >32 | N/A |
| Dihydroauroglaucin (4) | 128 | >128 | 128 | >128 | 128 | >128 | 64 | >128 | >128 | N/A | 8 | 32 | >128 | N/A | >128 | N/A |
| Flavoglaucin (5) | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A | >32 | N/A |
| Isodihydroauroglaucin (6) | 64 | 128 | 64 | 128 | 32 | 64 | 64 | >128 | >128 | N/A | 4 | 16 | >128 | N/A | >128 | N/A |
| Neoechinulin D (7) | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | 64 | >128 | >128 | N/A | >128 | N/A |
| Asperflavin (8) | 64 | >128 | 128 | >128 | 64 | >128 | >128 | N/A | >128 | N/A | 32 | 128 | >128 | N/A | >128 | N/A |
| Cinnalutein (9) | >128 | N/A | >128 | N/A | 128 | >128 | >128 | N/A | >128 | N/A | 32 | >128 | >128 | N/A | >128 | N/A |
| Cyclo-l-Trp-l-Ala (10) | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A | >128 | N/A |
| Gentamicin | 0.5 | N/D | >128 | N/D | 1 | N/D | >128 | N/D | N/D | N/D | N/D | N/D | 1 | N/D | 0.5 | N/D |
| Ciprofloxacin | 0.25 | N/D | 64 | N/D | 8 | N/D | 0.25 | N/D | 0.5 | N/D | 1 | N/D | 0.12 | N/D | ≤0.06 | N/D |
| Erythromycin | 0.5 | N/D | 8 | N/D | 0.5 | N/D | 2 | N/D | 0.12 | N/D | 16 | N/D | N/D | N/D | N/D | N/D |
| Tetracycline | 0.12 | N/D | 1 | N/D | 1 | N/D | 8 | N/D | 0.12 | N/D | ≤0.06 | N/D | 8 | N/D | 1 | N/D |
| Fusidic acid | 0.12 | N/D | N/D | N/D | N/D | N/D | 4 | N/D | 4 | N/D | N/D | N/D | N/D | N/D | N/D | N/D |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bovio, E.; Garzoli, L.; Poli, A.; Luganini, A.; Villa, P.; Musumeci, R.; McCormack, G.P.; Cocuzza, C.E.; Gribaudo, G.; Mehiri, M.; et al. Marine Fungi from the Sponge Grantia compressa: Biodiversity, Chemodiversity, and Biotechnological Potential. Mar. Drugs 2019, 17, 220. https://doi.org/10.3390/md17040220
Bovio E, Garzoli L, Poli A, Luganini A, Villa P, Musumeci R, McCormack GP, Cocuzza CE, Gribaudo G, Mehiri M, et al. Marine Fungi from the Sponge Grantia compressa: Biodiversity, Chemodiversity, and Biotechnological Potential. Marine Drugs. 2019; 17(4):220. https://doi.org/10.3390/md17040220
Chicago/Turabian StyleBovio, Elena, Laura Garzoli, Anna Poli, Anna Luganini, Pietro Villa, Rosario Musumeci, Grace P. McCormack, Clementina E. Cocuzza, Giorgio Gribaudo, Mohamed Mehiri, and et al. 2019. "Marine Fungi from the Sponge Grantia compressa: Biodiversity, Chemodiversity, and Biotechnological Potential" Marine Drugs 17, no. 4: 220. https://doi.org/10.3390/md17040220
APA StyleBovio, E., Garzoli, L., Poli, A., Luganini, A., Villa, P., Musumeci, R., McCormack, G. P., Cocuzza, C. E., Gribaudo, G., Mehiri, M., & Varese, G. C. (2019). Marine Fungi from the Sponge Grantia compressa: Biodiversity, Chemodiversity, and Biotechnological Potential. Marine Drugs, 17(4), 220. https://doi.org/10.3390/md17040220