Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling

 ,
, 

Abstract
:1. Introduction
2. Results
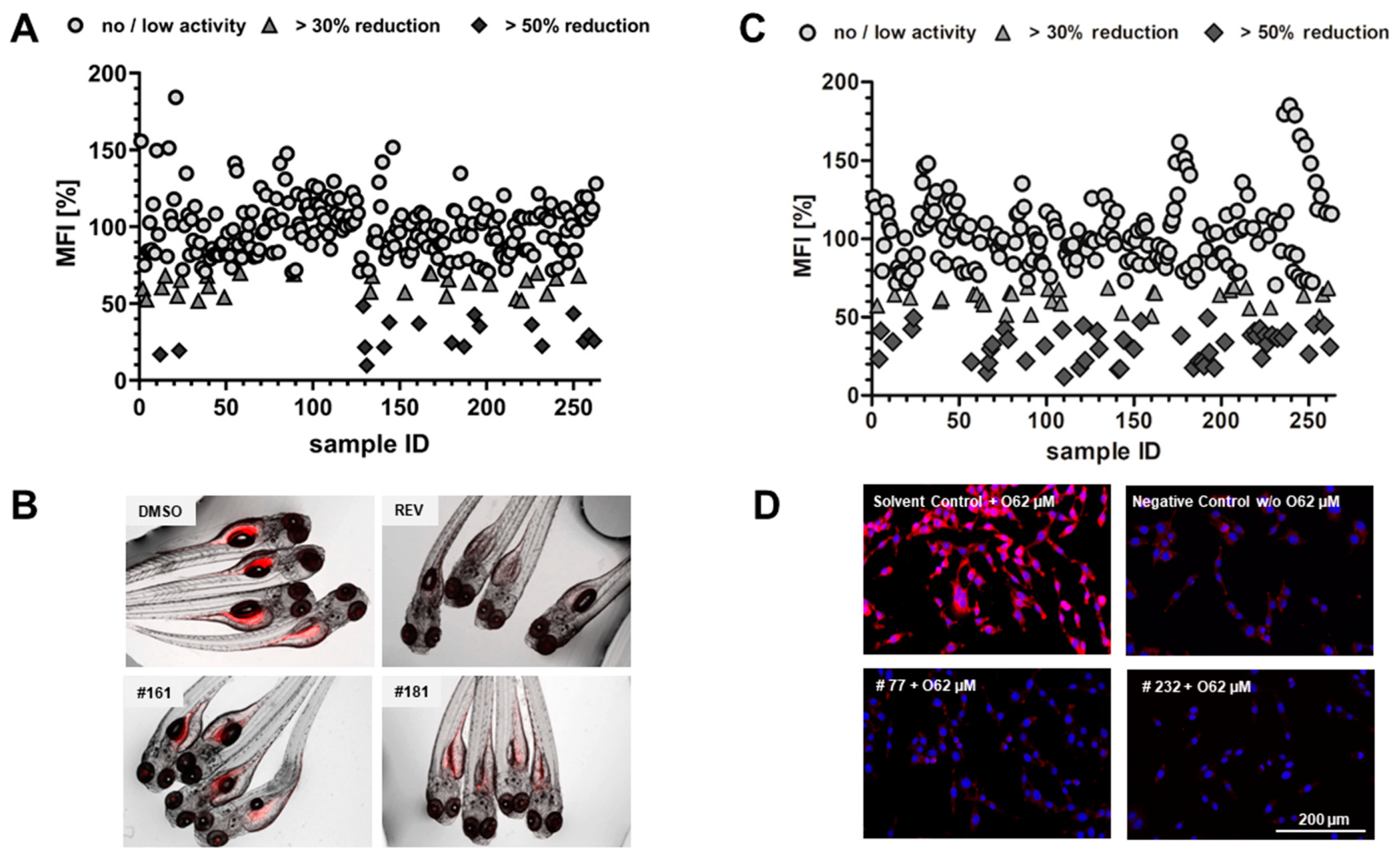
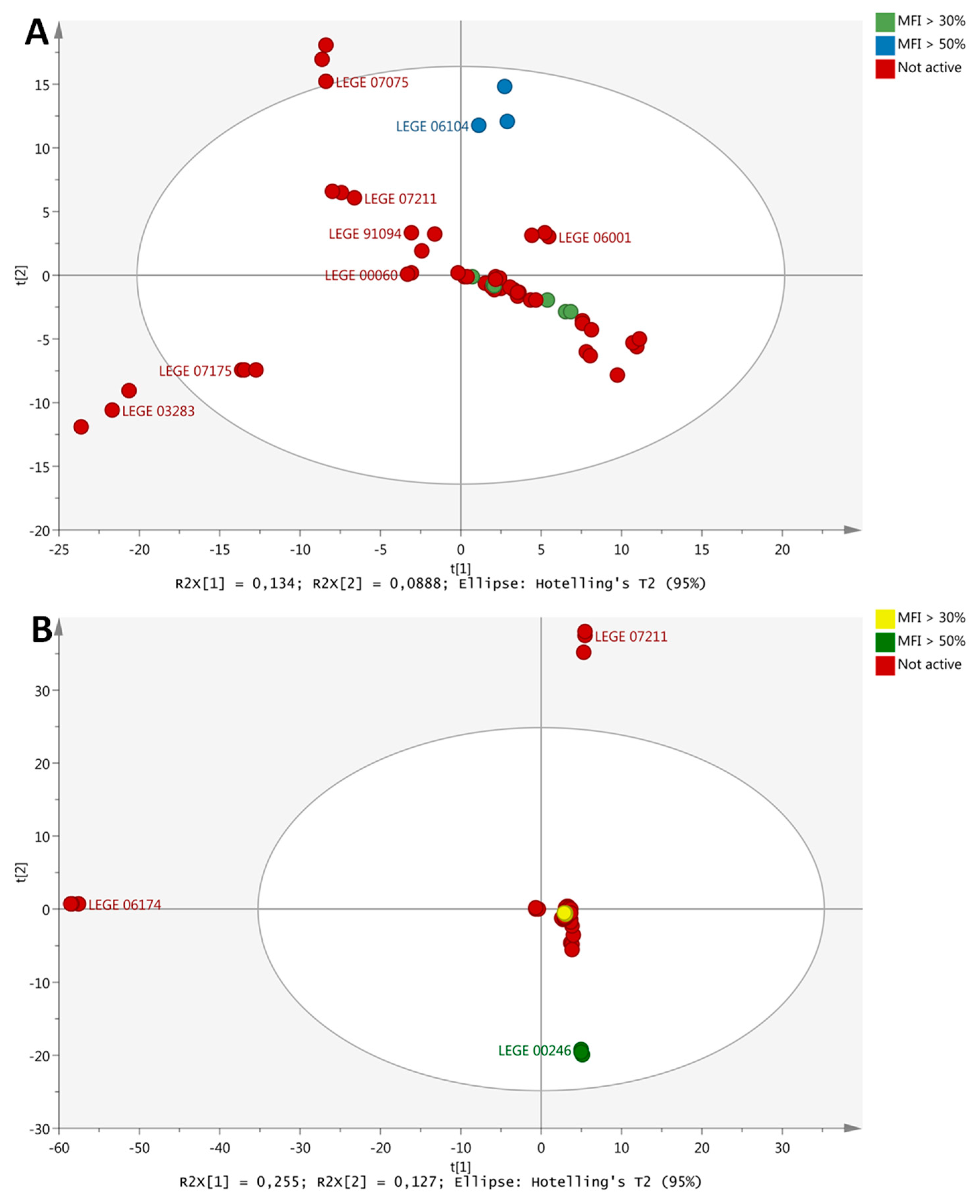
2.1. Lipid Reducing Activity in Zebrafish Larvae
2.2. Anti-Steatosis Activity in HepG2 Cells
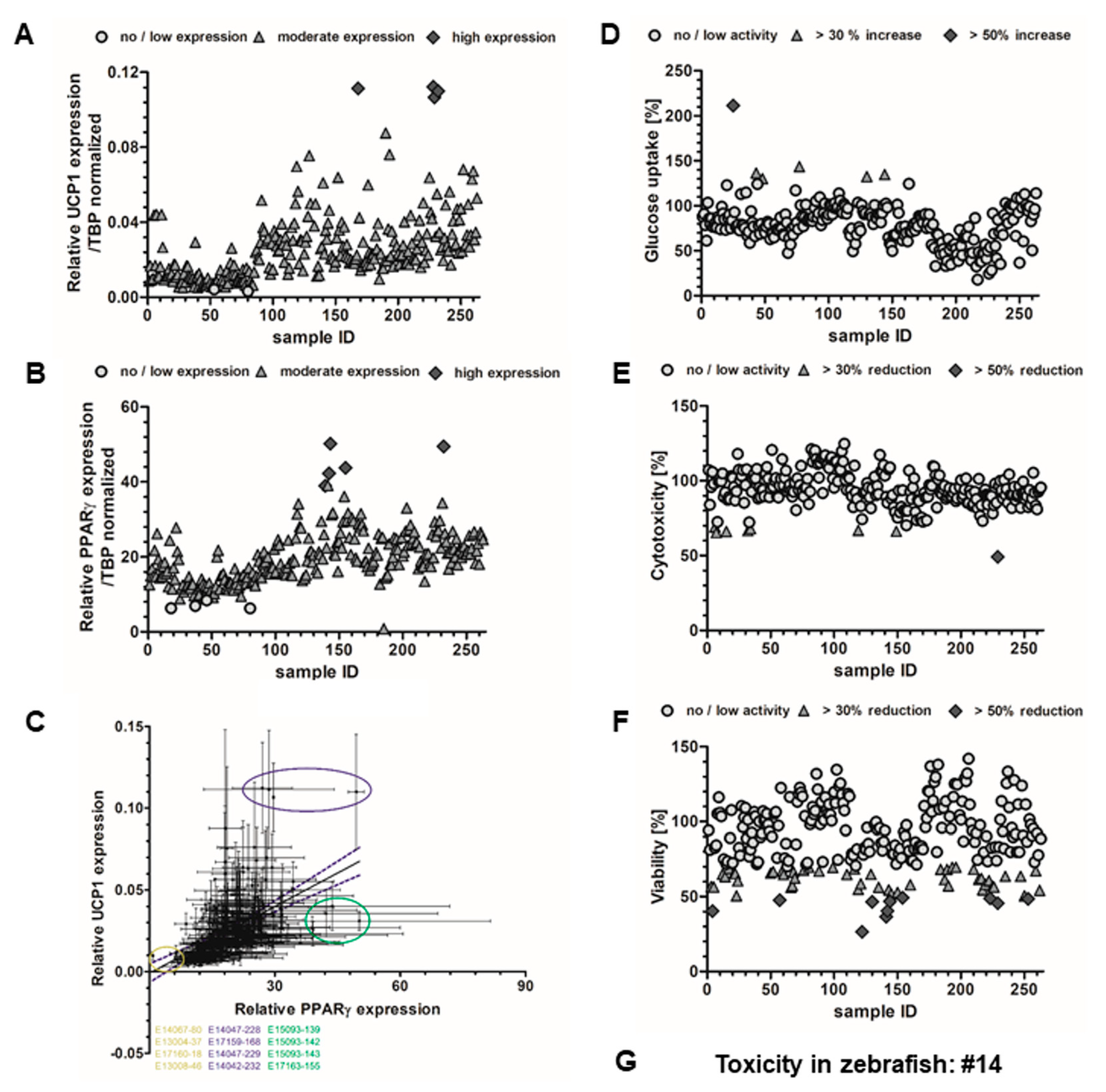
2.3. Effects on Brown Adipocyte Differentiation and Thermogenic Gene Expression
2.4. Glucose Uptake in HepG2 Cells
2.5. Toxicity Evaluation
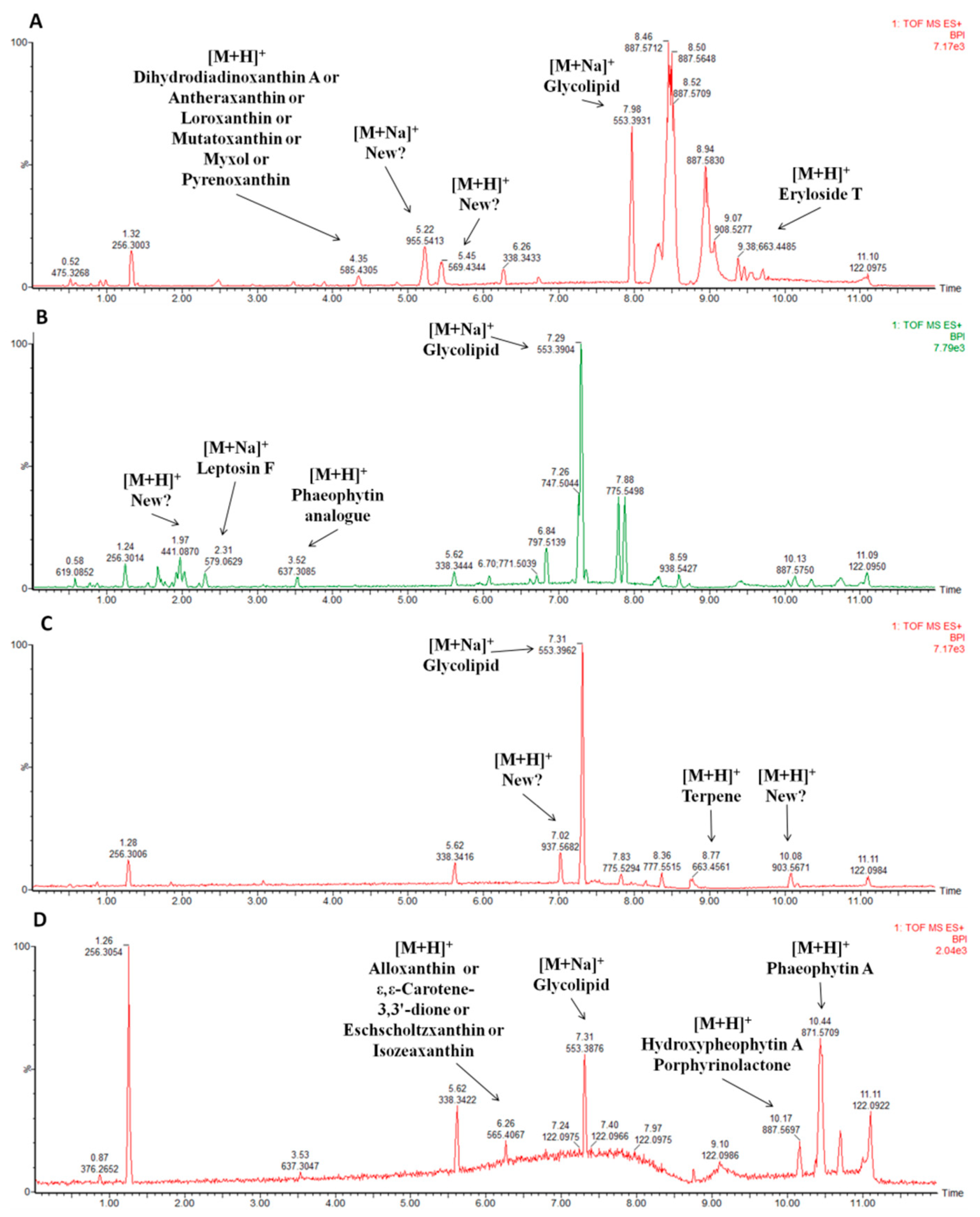
2.6. Metabolite Profiling
3. Discussion
4. Materials and Methods
4.1. Growth of Cyanobacteria and Construction of Screening Library
4.1.1. Vacuum Liquid Chromatography Fractionation
4.1.2. Increasing Polarity Extraction
4.2. Zebrafish Nile Red Fat Metabolism Assay
4.3. Cell Culture
4.4. Anti-Steatosis Assay in HepG2 Cells and Sulforhodamine B (SRB) Assay
4.5. Glucose Uptake Assay in HepG2 Cells and MTT Assay
4.6. Brown Adipocyte Differentiation
PPARγ and UCP-1 mRNA Expression by Real-Time PCR
4.7. Metabolite Profiling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Collaboration, N.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Kelly, T.; Yang, W.; Chen, C.S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Magno, C.P.; Lane, K.T.; Hinojosa, M.W.; Lane, J.S. Association of Hypertension, Diabetes, Dyslipidemia, and Metabolic Syndrome with Obesity: Findings from the National Health and Nutrition Examination Survey, 1999 to 2004. J. Am. Coll. Surg. 2008, 207, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Berger, N.A. Obesity and cancer pathogenesis. Ann. N. Y. Acad. Sci. 2014, 1311, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; le Roux, C.W.; Docherty, N.G. Morbidity and mortality associated with obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M.; Gokce, N. Obesity and cardiovascular disease. Circulation 2012, 125, 1178–1182. [Google Scholar] [CrossRef]
- Kang, J.G.; Park, C.-Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab. 2012, 36, 13–25. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Koyama, T.; Kawazoe, Y.; Iwasaki, A.; Ohno, O.; Suenaga, K.; Uemura, D. Anti-obesity activities of the yoshinone A and the related marine γ-pyrone compounds. J Antibiot. 2016, 69, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Noinart, J.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Urbatzka, R.; Freitas, S.; Lee, M.; Silva, A.M.S.; Pinto, M.M.M.; et al. A New Ergosterol Analog, a New Bis-Anthraquinone and Anti-Obesity Activity of Anthraquinones from the Marine Sponge-Associated Fungus Talaromyces stipitatus KUFA 0207. Mar. Drugs 2017, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural Product Biosynthetic Diversity and Comparative Genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef]
- Ramos, V.; Morais, J.; Castelo-Branco, R.; Pinheiro, Â.; Martins, J.; Regueiras, A.; Pereira, A.L.; Lopes, V.R.; Frazão, B.; Gomes, D.; et al. Cyanobacterial diversity held in microbial biological resource centers as a biotechnological asset: the case study of the newly established LEGE culture collection. J. Appl. Phycol. 2018, 30, 1437–1451. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring bioactive properties of marine cyanobacteria isolated from the Portuguese coast: high potential as a source of anticancer compounds. Mar. Drugs 2013, 12, 98–114. [Google Scholar] [CrossRef]
- Gómez-Lechón, M.J.; Donato, M.T.; Martínez-Romero, A.; Jiménez, N.; Castell, J.V.; O’Connor, J.-E. A human hepatocellular in vitro model to investigate steatosis. Chem. Biol. Interact. 2007, 165, 106–116. [Google Scholar] [CrossRef]
- MarineLit. Available online: http://pubs.rsc.org/marinlit/ (accessed on 10 January 2019).
- Royal Society of Chemistry. ChemSpider. Available online: http://www.chemspider.com/ (accessed on 4 March 2019).
- Society, A.C. SciFinder. Available online: https://scifinder.cas.org/ (accessed on 4 March 2019).
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Castro, M.; Preto, M.; Vasconcelos, V.; Urbatzka, R. Obesity: The Metabolic Disease, Advances on Drug Discovery and Natural Product Research. Curr. Top. Med. Chem. 2016, 16, 2577–2604. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2008, 8, 69. [Google Scholar] [CrossRef]
- Zheng, W.; Thorne, N.; McKew, J.C. Phenotypic screens as a renewed approach for drug discovery. Drug Discov. Today 2013, 18, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Giacomotto, J.; Ségalat, L. High-throughput screening and small animal models, where are we? Br. J. Pharmacol. 2010, 160, 204–216. [Google Scholar] [CrossRef] [Green Version]
- Urbatzka, R.; Freitas, S.; Palmeira, A.; Almeida, T.; Moreira, J.; Azevedo, C.; Afonso, C.; Correia-da-Silva, M.; Sousa, E.; Pinto, M.; et al. Lipid reducing activity and toxicity profiles of a library of polyphenol derivatives. Eur. J. Med. Chem. 2018, 151, 272–284. [Google Scholar] [CrossRef]
- Hibasami, H.; Kyohkon, M.; Ohwaki, S.; Katsuzaki, H.; Imai, K.; Nakagawa, M.; Ishi, Y.; Komiya, T. Pheophorbide a, a moiety of chlorophyll a, induces apoptosis in human lymphoid leukemia molt 4B cells. Int. J. Mol. Med. 2000, 6, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.M.-K.; Chan, J.Y.-W.; Au, S.W.-N.; Kong, S.-K.; Tsui, S.K.-W.; Waye, M.M.-Y.; Mak, T.C.-W.; Fong, W.-P.; Fung, K.-P. Pheophorbide a, an active compound isolated from Scutellaria barbata, possesses photodynamic activities by inducing apoptosis in human hepatocellular carcinoma. Cancer Biol. Ther. 2006, 5, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, A.; Asha, V.V.; Nair, S.A.; Sasidharan, S.P.; Sureshkumar, P.K.; Rajendran, K.N.; Karunagaran, D.; Ramalingam, K. Chlorophyll Revisited: Anti-inflammatory Activities of Chlorophyll a and Inhibition of Expression of TNF-α Gene by the Same. Inflammation 2012, 35, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, J.J.; Park, B.-K.; Yoon, S.J.; Choi, J.E.; Jin, M. Pheophytin a and chlorophyll a suppress neuroinflammatory responses in lipopolysaccharide and interferon-γ-stimulated BV2 microglia. Life Sci. 2014, 103, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Bui-Xuan, N.-H.; Tang, P.M.-K.; Wong, C.-K.; Chan, J.Y.-W.; Cheung, K.K.Y.; Jiang, J.L.; Fung, K.-P. Pheophorbide a: A photosensitizer with immunostimulating activities on mouse macrophage RAW 264.7 cells in the absence of irradiation. Cell. Immunol. 2011, 269, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Afiyatullov, S.S.; Kalinovsky, A.I.; Antonov, A.S.; Ponomarenko, L.P.; Dmitrenok, P.S.; Aminin, D.L.; Krasokhin, V.B.; Nosova, V.M.; Kisin, A.V. Isolation and Structures of Erylosides from the Carribean Sponge Erylus goffrilleri. J. Nat. Prod. 2007, 70, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.-J.; Wi, H.-R.; Kim, H.-R.; Park, K.W.; Hwang, K.-A. Pinus densiflora Sieb. et Zucc. Alleviates Lipogenesis and Oxidative Stress during Oleic Acid-Induced Steatosis in HepG2 Cells. Nutrients 2014, 6, 2956–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Li, Z.; Manabe, Y.; Kim, M.; Goto, T.; Kawada, T.; Sugawara, T. Siphonaxanthin, a Carotenoid From Green Algae, Inhibits Lipogenesis in Hepatocytes via the Suppression of Liver X Receptor α Activity. Lipids 2018, 53, 41–52. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chen, Y.-L.; Huang, W.-C.; Liou, C.-J. Fucoxanthin attenuates fatty acid-induced lipid accumulation in FL83B hepatocytes through regulated Sirt1/AMPK signaling pathway. Biochem. Biophys. Res. Commun. 2018, 495, 197–203. [Google Scholar] [CrossRef]
- Yanagihara, M.; Sasaki-Takahashi, N.; Sugahara, T.; Yamamoto, S.; Shinomi, M.; Yamashita, I.; Hayashida, M.; Yamanoha, B.; Numata, A.; Yamori, T.; et al. Leptosins isolated from marine fungus Leptoshaeria species inhibit DNA topoisomerases I and/or II and induce apoptosis by inactivation of Akt/protein kinase B. Cancer Sci. 2005, 96, 816–824. [Google Scholar] [CrossRef] [Green Version]
- Chuyen, H.V.; Eun, J.-B. Marine carotenoids: Bioactivities and potential benefits to human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Elissawy, A.M.; El-Shazly, M.; Ebada, S.S.; Singab, A.B.; Proksch, P. Bioactive Terpenes from Marine-Derived Fungi. Mar. Drugs 2015, 13, 1966–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebel, R. Terpenes from Marine-Derived Fungi. Mar. Drugs 2010, 8, 2340–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, J.-K.; Ha, T.K.Q.; Oh, D.-C.; Oh, W.-K.; Oh, K.-B.; Shin, J. Polyoxygenated Steroids from the Sponge Clathria gombawuiensis. J. Nat. Prod. 2017, 80, 3224–3233. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Bishbishy, M.H.E.; Habtemariam, S.; Salehi, B.; Sharifi-Rad, M.; Martins, N.; Sharifi-Rad, J. Looking at Marine-Derived Bioactive Molecules as Upcoming Anti-Diabetic Agents: A Special Emphasis on PTP1B Inhibitors. Molecules 2018, 23, 3334. [Google Scholar] [CrossRef] [PubMed]
- Abdjul, D.B.; Kanno, S.-I.; Yamazaki, H.; Ukai, K.; Namikoshi, M. A dimeric urea of the bisabolene sesquiterpene from the Okinawan marine sponge Axinyssa sp. inhibits protein tyrosine phosphatase 1B activity in Huh-7 human hepatoma cells. Bioorg. Med. Chem. Lett. 2016, 26, 315–317. [Google Scholar] [CrossRef]
- Villarroya, F.; Iglesias, R.; Giralt, M. PPARs in the Control of Uncoupling Proteins Gene Expression. PPAR Res. 2007, 2007, 74364. [Google Scholar] [CrossRef]
- Maeda, H. Nutraceutical effects of fucoxanthin for obesity and diabetes therapy: a review. J. Oleo Sci. 2015, 64, 125–132. [Google Scholar] [CrossRef]
- Bigagli, E.; Cinci, L.; Niccolai, A.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; D’Ambrosio, M.; Lodovici, M.; Tredici, M.R.; Luceri, C. Preliminary data on the dietary safety, tolerability and effects on lipid metabolism of the marine microalga Tisochrysis lutea. Algal Res. 2018, 34, 244–249. [Google Scholar] [CrossRef]
- Wu, C.; Kim, H.K.; van Wezel, G.P.; Choi, Y.H. Metabolomics in the natural products field – a gateway to novel antibiotics. Drug Discov. Today Technol. 2015, 13, 11–17. [Google Scholar] [CrossRef]
- Einarsdottir, E.; Magnusdottir, M.; Astarita, G.; Köck, M.; Ögmundsdottir, H.; Thorsteinsdottir, M.; Rapp, H.; Omarsdottir, S.; Paglia, G. Metabolic Profiling as a Screening Tool for Cytotoxic Compounds: Identification of 3-Alkyl Pyridine Alkaloids from Sponges Collected at a Shallow Water Hydrothermal Vent Site North of Iceland. Mar. Drugs 2017, 15, 52. [Google Scholar] [CrossRef]
- Olsen, E.K.; Soderholm, K.L.; Isaksson, J.; Andersen, J.H.; Hansen, E. Metabolomic Profiling Reveals the N-Acyl-Taurine Geodiataurine in Extracts from the Marine Sponge Geodia macandrewii (Bowerbank). J. Nat. Prod. 2016, 79, 1285–1291. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae Norwegian; Institute for Water Research, Blindern: Oslo, Norway, 1972; Volume B-11/69, p. 5. [Google Scholar]
- Costa, M.S.; Rego, A.; Ramos, V.; Afonso, T.B.; Freitas, S.; Preto, M.; Lopes, V.; Vasconcelos, V.; Magalhães, C.; Leão, P.N. The conifer biomarkers dehydroabietic and abietic acids are widespread in Cyanobacteria. Sci. Rep. 2016, 6, 23436. [Google Scholar] [CrossRef]
- ImageJ. Available online: https://imagej.nih.gov/ij/index.html (accessed on 1 March 2019).
- Pramme-Steinwachs, I.; Jastroch, M.; Ussar, S. Extracellular calcium modulates brown adipocyte differentiation and identity. Sci Rep. 2017, 7, 8888. [Google Scholar] [CrossRef] [Green Version]
- Donato, M.T.; Tolosa, L.; Jiménez, N.; Castell, J.V.; Gómez-Lechón, M.J. High-Content Imaging Technology for the Evaluation of Drug-Induced Steatosis Using a Multiparametric Cell-Based Assay. J. Biomol. Screen. 2011, 17, 394–400. [Google Scholar] [CrossRef]
- Dave, T.; Tilles, A.W.; Vemula, M. A Cell-Based Assay to Investigate Hypolipidemic Effects of Nonalcoholic Fatty Liver Disease Therapeutics. SLAS Discov. 2018, 23, 274–282. [Google Scholar] [CrossRef]
- Hassanein, M.; Weidow, B.; Koehler, E.; Bakane, N.; Garbett, S.; Shyr, Y.; Quaranta, V. Development of High-Throughput Quantitative Assays for Glucose Uptake in Cancer Cell Lines. Mol. Imaging Biol. 2011, 13, 840–852. [Google Scholar] [CrossRef]
- Plaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction/Extract | Markers | Variance PC1 (%) | Variance PC2 (%) | Fractions with Potential to Produce Different Metabolites |
|---|---|---|---|---|
| A (IPE) | 485 | 18 | 14 | 201, 216, 225, 228, 234, 240, 243, 249, 255, 258 |
| B (IPE) | 1131 | 19 | 9 | 199, 202, 205, 214, 217, 220, 226, 229, 235, 244, 253, 256 |
| C (IPE) | 1028 | 13 | 13 | 197, 221, 224, 248, 251, 254, 260 |
| A (VLC) | 815 | 15 | 11 | 1, 54, 75, 106 |
| B (VLC) | 628 | 19 | 11 | 20, 29, 46, 64, 139 |
| C (VLC) | 816 | 15 | 14 | 3, 12, 21, 77, 108 |
| D (VLC) | 943 | 16 | 10 | 19, 66, 88, 141 |
| E (VLC) | 1002 | 13 | 9 | 23, 40, 58, 67, 110, 130, 142, 160 |
| F (VLC) | 914 | 26 | 11 | 80, 121 |
| G (VLC) | 1228 | 26 | 13 | 69, 122, 180 |
| H (VLC) | 1178 | 11 | 10 | 92, 103, 123, 134, 154, 181 |
| I (VLC) | 1105 | 27 | 15 | 83, 164 |
| Bioactivity | Selected Fraction |
|---|---|
| Zebrafish—Lipid reducing | LEGE07175 H/#134 |
| LEGE00246 G/#180 | |
| LEGE07172 A/#240 | |
| LEGE07172 C/#242 | |
| LEGE07173 B/#256 | |
| HepG2—Anti-steatosis | LEGE07084 D/#48 |
| LEGE03283 C/#108 | |
| LEGE03283 D/#109 | |
| LEGE07167 B/#199 | |
| LEGE07160 B/#202 | |
| LEGE06134 B/#220 | |
| HepG2—Glucose uptake | LEGE06001 G/#25 |
| LEGE06104 E/#58 | |
| LEGE07212 C/#77 | |
| LEGE07175 E/#130 | |
| Brown adipocytes—PPARγ and UCP-1 inducing activities | LEGE00247 D/#168 |
| LEGE06137 A/#228 | |
| LEGE06097 B/#232 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, M.; Rosa, F.; Ribeiro, T.; Hernandez-Bautista, R.; Bonaldo, M.; Gonçalves Silva, N.; Eiríksson, F.; Thorsteinsdóttir, M.; Ussar, S.; Urbatzka, R. Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling. Mar. Drugs 2019, 17, 280. https://doi.org/10.3390/md17050280
Costa M, Rosa F, Ribeiro T, Hernandez-Bautista R, Bonaldo M, Gonçalves Silva N, Eiríksson F, Thorsteinsdóttir M, Ussar S, Urbatzka R. Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling. Marine Drugs. 2019; 17(5):280. https://doi.org/10.3390/md17050280
Chicago/Turabian StyleCosta, Margarida, Filipa Rosa, Tiago Ribeiro, Rene Hernandez-Bautista, Marco Bonaldo, Natália Gonçalves Silva, Finnur Eiríksson, Margrét Thorsteinsdóttir, Siegfried Ussar, and Ralph Urbatzka. 2019. "Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling" Marine Drugs 17, no. 5: 280. https://doi.org/10.3390/md17050280
APA StyleCosta, M., Rosa, F., Ribeiro, T., Hernandez-Bautista, R., Bonaldo, M., Gonçalves Silva, N., Eiríksson, F., Thorsteinsdóttir, M., Ussar, S., & Urbatzka, R. (2019). Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling. Marine Drugs, 17(5), 280. https://doi.org/10.3390/md17050280