Antihypertensive Effect in Vivo of QAGLSPVR and Its Transepithelial Transport Through the Caco-2 Cell Monolayer



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
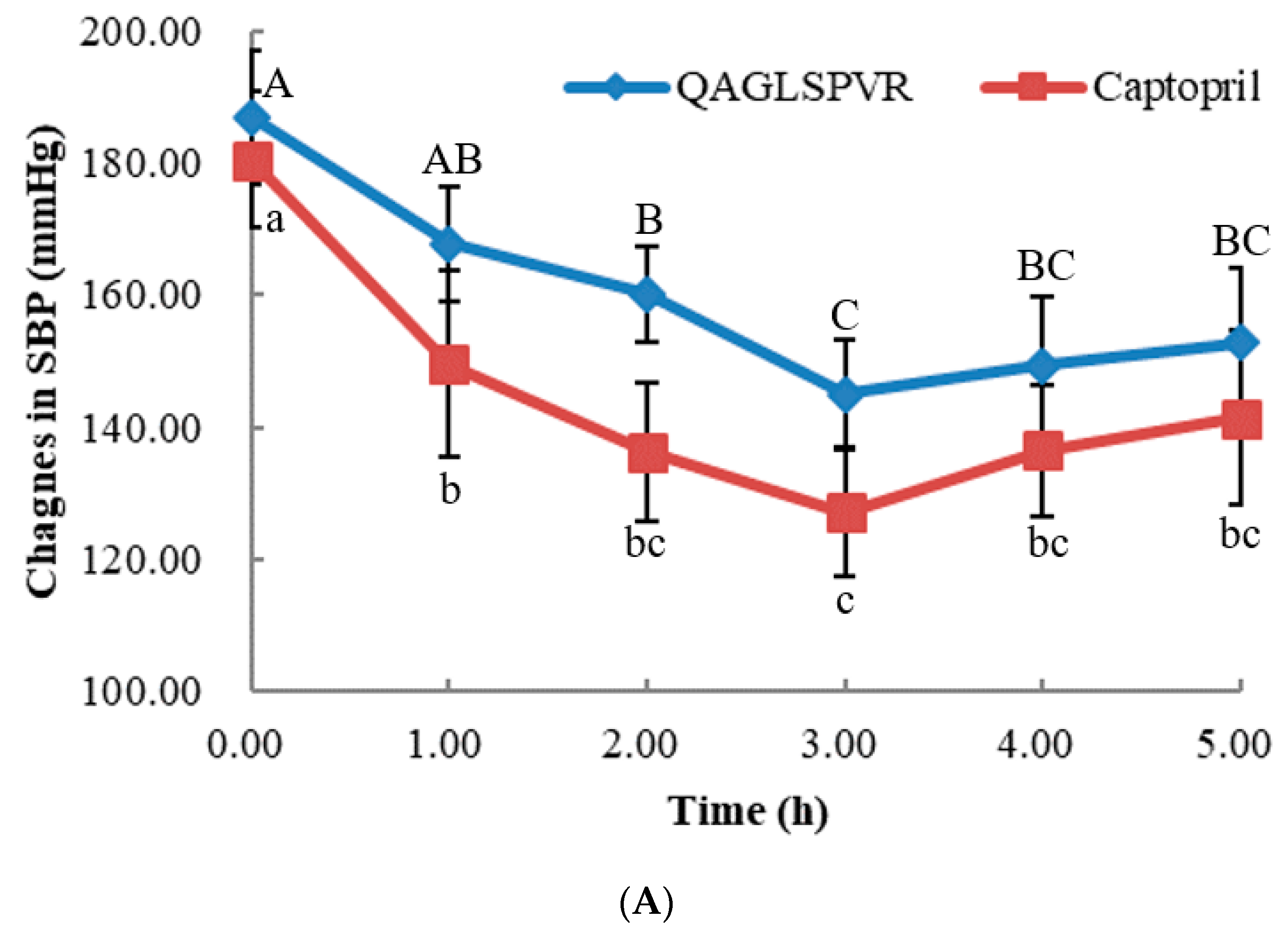
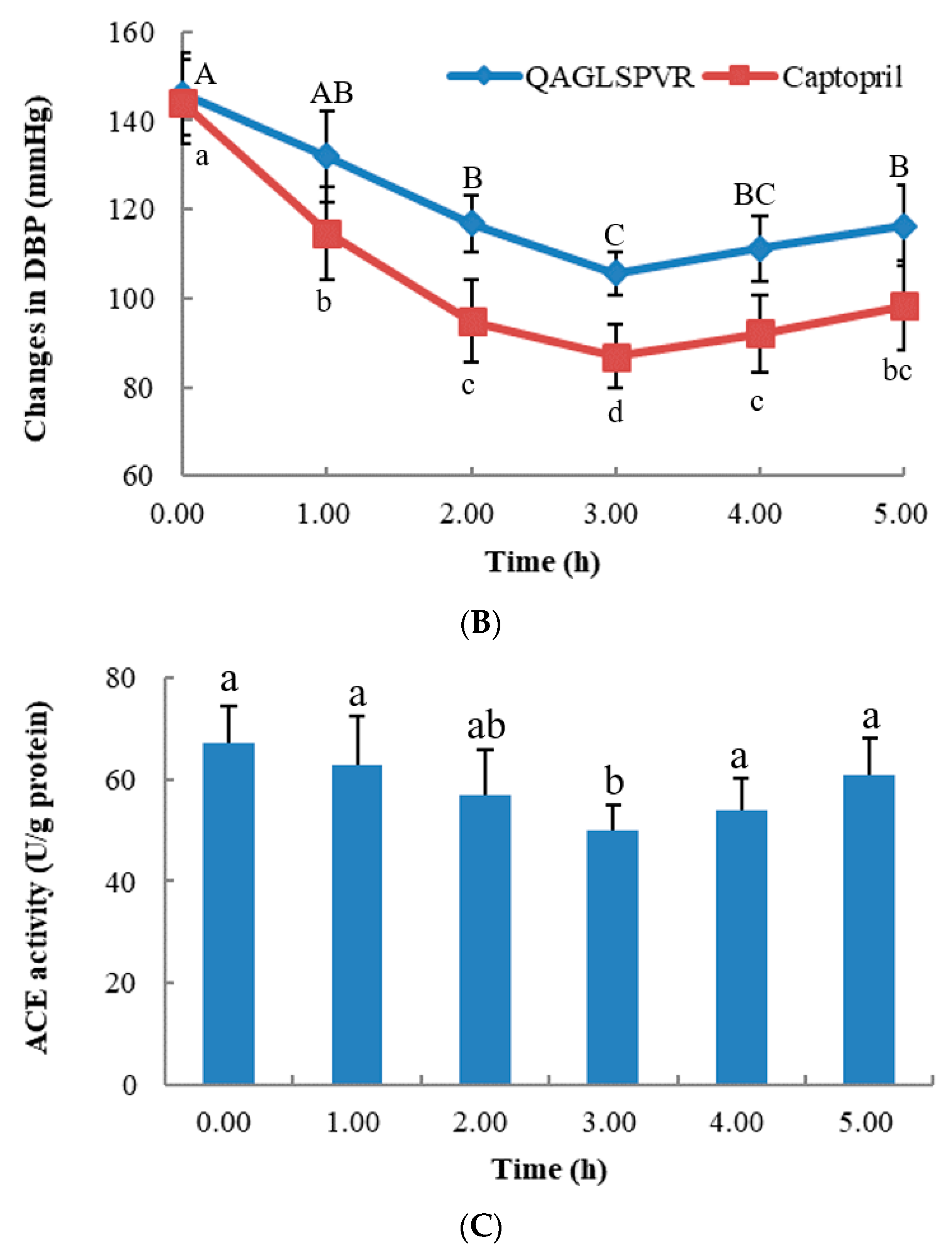
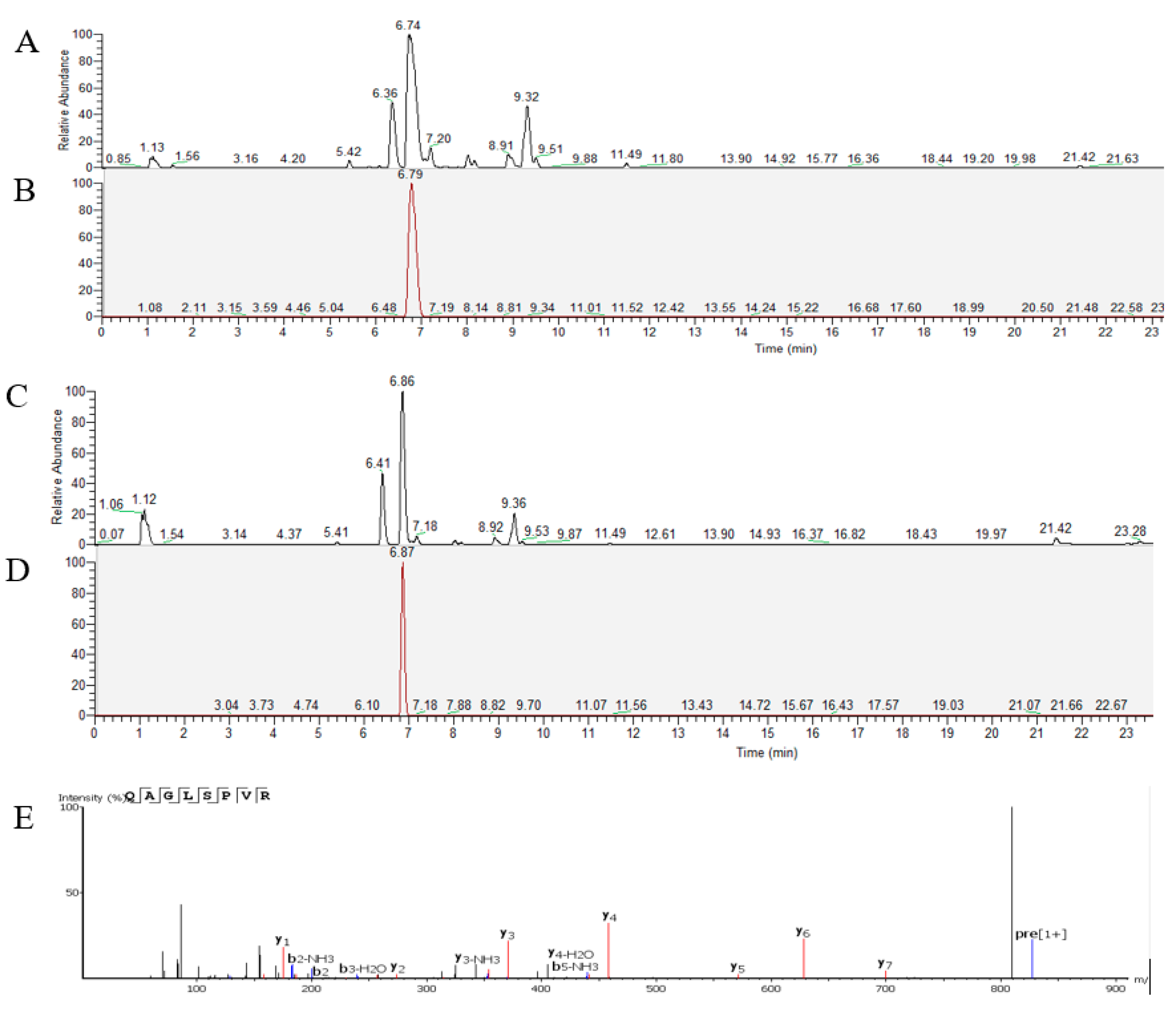
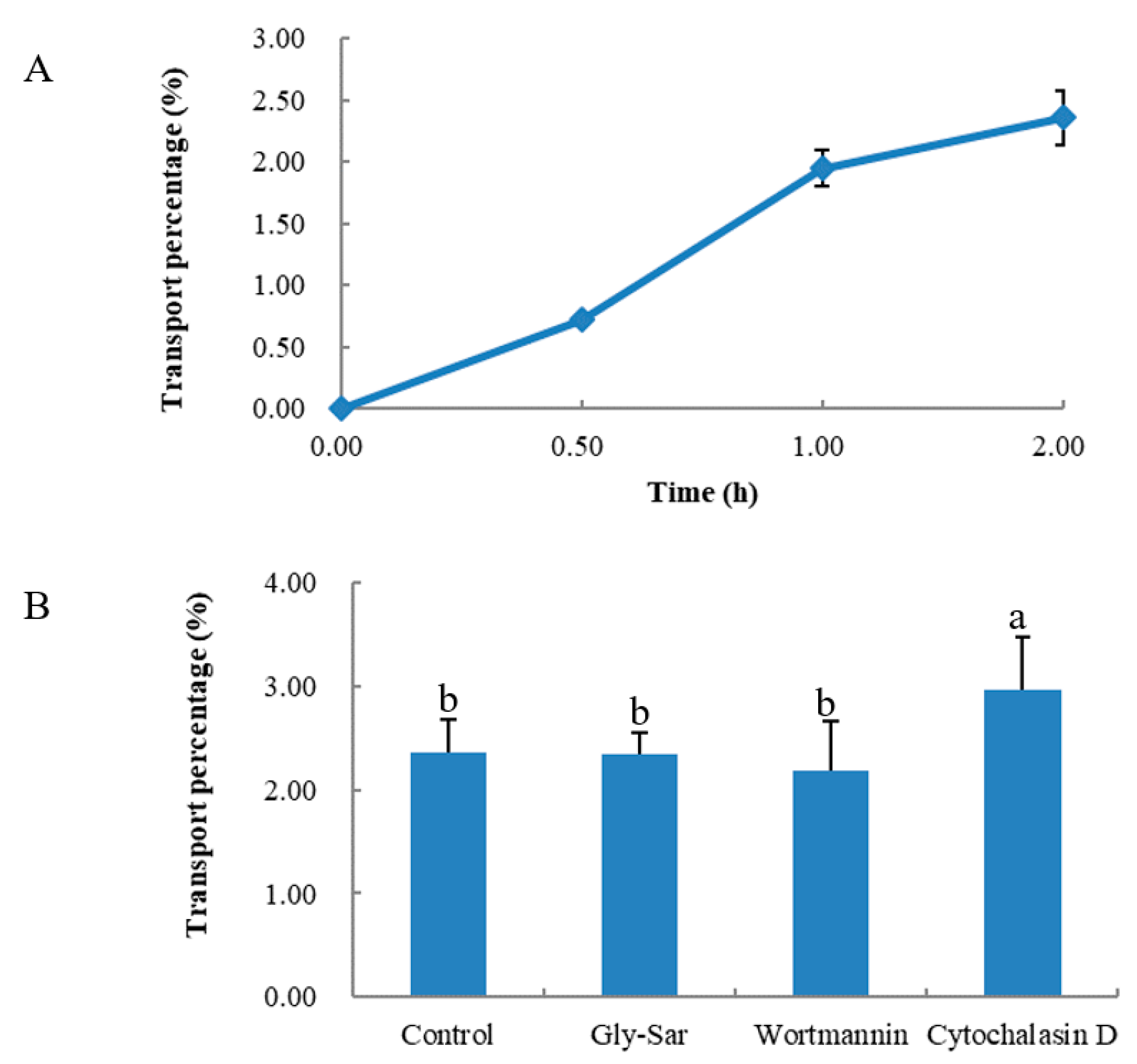
2. Results
2.1. Changes in Blood Pressure over Time
2.2. Transport through the Caco-2 Cell Monolayer
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Animal Treatment
4.2.1. Animals
4.2.2. Measurement of Blood Pressure
4.2.3. Determination of Serum ACE Activities
4.3. Transepithelial Transport of QAGLSPVR
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Toldra, F.; Nasri, M. In silico analysis and molecular docking study of angiotensin I-converting enzyme inhibitory peptides from smooth-hound viscera protein hydrolysates fractionated by ultrafiltration. Food Chem. 2018, 239, 453–463. [Google Scholar] [CrossRef]
- Ngo, D.H.; Vo, T.S.; Ryu, B.M.; Kim, S.K. Angiotensin-I-converting enzyme (ACE) inhibitory peptides from Pacific cod skin gelatin using ultrafiltration membranes. Process Biochem. 2016, 51, 1622–1628. [Google Scholar]
- Zhuang, Y.; Sun, L.; Li, B. Production of the angiotensin-I-converting enzyme (ACE)-inhibitory peptide from hydrolysates of jellyfish (Rhopilemaesculentum) collagen. Food Bio. Technol. 2012, 5, 1622–1629. [Google Scholar] [CrossRef]
- Zhuang, Y.; Sun, L.; Zhang, Y.; Liu, G. Antihypertensive effect of long-term oral administration of jellyfish (Rhopilemaesculentum) collagen peptides on renovascular hypertension. Mar. Drugs 2012, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Chen, X.; Wu, Y.; Zhang, L.; Huang, W.; Yuan, Y.; Fang, M.; Xie, J.; Wei, D. Angiotensin I-converting enzyme inhibitory peptides from Sipuncula (Phascolosoma esculenta): Purification, identification, molecular docking and antihypertensive effects on spontaneously hypertensive rats. Process Biochem. 2017, 63, 84–95. [Google Scholar] [CrossRef]
- Foltz, M.; van der Pijl, P.C.; Duchateau, G.S. Current in vitro testing of bioactive peptides is not valuable. J. Nutr. 2010, 140, 117–118. [Google Scholar] [CrossRef] [PubMed]
- García-Mora, P.; Martín-Martínez, M.; Bonache, M.A.; González-Múniz, R.; Peñas, E.; Frias, J.; Martinez-Villaluenga, C. Identification, functional gastrointestinal stability and molecular docking studies of lentil peptides with dual antioxidant and angiotensin I converting enzyme inhibitory activities. Food Chem. 2017, 221, 464–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Zhang, Y.; Jiang, Y.; Wang, L.; Liu, B.; Liu, J. Transport of egg white ACE-inhibitory peptide, Gln-Ile-Gly-Leu-Phe, in human intestinal Caco-2 cell monolayers with cytoprotective effect. J. Agric. Food Chem. 2014, 62, 3177–3182. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A.; Vistoli, G.; Lammi, C. Behavior of three hypocholesterolemic peptides from soy protein in an intestinal model based on differentiated Caco-2 cell. J. Func. Food. 2018, 45, 363–370. [Google Scholar] [CrossRef]
- Yuan, L.; Sun, L.; Zhuang, Y. Preparation and identification of novel inhibitory angiotensin-I-converting enzyme peptides from tilapia skin gelatin hydrolysates: Inhibition kinetics and molecular docking. Food Func. 2018, 9, 5251–5259. [Google Scholar]
- Balti, R.; Bougatef, A.; Sila, A.; Guillochon, D.; Dhulster, P.; Nedjar-Arroume, N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chem. 2015, 170, 519–525. [Google Scholar] [CrossRef]
- Fujita, H.; Yokoyama, K.; Yoshikawa, M. Classification and antihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar]
- Zhao, Y.; Li, B.; Dong, S.; Liu, Z.; Zhao, X.; Wang, J.; Zeng, M. A novelACE inhibitory peptide isolated from Acaudina molpadioidea hydrolysate. Peptides 2009, 30, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Albaladejo, P.; Bouaziz, H.; Duriez, M.; Gohlke, P.; Levy, B.I.; Safar, M.E.; Benetos, A. Angiotensin converting enzyme inhibition prevents the increase in aortic collagen in rats. Hypertension 1994, 23, 74. [Google Scholar] [CrossRef] [PubMed]
- Boonla, O.; Kukongviriyapan, U.; Pakdeechote, P.; Kukongviriyapan, V.; Pannangpetch, P.; Thawornchinsombut, S. Peptides-derived from thai rice bran improves endothelial function in 2K-1C renovascular hypertensive rats. Nutrients 2015, 7, 5783–5799. [Google Scholar] [CrossRef]
- Ding, L.; Wang, L.; Zhang, T.; Yu, Z.; Liu, J. Hydrolysis and transepithelial transport of two corn gluten derived bioactive peptides in human Caco-2 cell monolayers. Food Res. Int. 2018, 106, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.; Kenny, A.J.; Turner, A.J. A survey of membrane peptidases in two human colonic cell lines, Caco-2 and HT-29. Biochem. J. 1992, 284, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Wang, L.; Yu, Z.; Zhang, T.; Liu, J. Digestion and absorption of an egg white ACE-inhibitory peptide in human intestinal Caco-2 cell monolayers. Int. J. Food Sci. Nut. 2016, 67, 111–116. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Sanchón, J.; Recio, I.; Hernández-Ledesma, B. Transepithelial transport of lunasin and derived peptides: Inhibitory effects on the gastrointestinal cancer cells viability. J. Food Comp. Ana. 2018, 68, 101–110. [Google Scholar] [CrossRef]
- Cakir-Kiefer, C.; Miclo, L.; Balandras, F.; Dary, A.; Soligot, C.; Le Roux, Y. Transport across Caco-2 cell monolayer and sensitivity to hydrolysis of two anxiolytic peptides from alpha(s1)-casein, alpha-casozepine, and alpha(s1)-casein-(f91−97): Effect of bile salts. J. Agric. Food Chem. 2011, 59, 11956–11965. [Google Scholar] [CrossRef]
- Martínez-Maqueda, D.; Miralles, B.; Recio, I.; Hernández-Ledesma, B. Antihypertensive peptides from food proteins: A review. Food Func. 2012, 3, 350–361. [Google Scholar] [CrossRef]
- Chen, M.; Li, B. The effect of molecular weights on the survivability of caseinderived antioxidant peptides after the simulated gastrointestinal digestion. Innov. Food Sci. Emer. Technol. 2012, 16, 341–348. [Google Scholar] [CrossRef]
- Miner-Williams, W.M.; Stevens, B.R.; Moughan, P.J. Are intact peptides absorbed from the healthy gut in the adult human. Nutr. Res. Rev. 2014, 27, 308–329. [Google Scholar] [CrossRef]
- Terada, T.; Inui, K.I. (Section A: Molecular, structural, and cellular biology of drug transporters) Peptide transporters: Structure, function, regulation and application for drug delivery. Curr. Drug Metab. 2004, 5, 85–94. [Google Scholar] [CrossRef]
- Ding, L.; Wang, L.; Zhang, Y.; Liu, J. Transport of antihypertensive peptide RVPSL, ovotransferrin 328–332, in human intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2015, 63, 8143–8150. [Google Scholar] [CrossRef]
- Shimizu, K.; Sato, M.; Zhang, Y.; Kouguchi, T.; Takahata, Y.; Morimatsu, F.; Shimizu, M. Molecular size of collagen peptide reverses the permeability of Caco-2 cells. Biosci. Biotechnol. Biochem. 2010, 74, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Quirόs, A.; Contreras, M.D.M.; Ramos, M.; Amigo, L.; Recio, I. Stability to gastrointestinal enzymes and structure–activity relationship of b-casein peptides with antihypertensive properties. Peptides 2009, 30, 1848–1853. [Google Scholar] [CrossRef]
- Guo, Y.; Gan, J.; Zhu, Q.; Zeng, X.; Sun, Y.; Wu, Z.; Pan, D. Transepithelial transport of milk-derived angiotensin I-converting enzyme inhibitory peptide with the RLSFNP sequence. J. Sci. Food Agric. 2017, 98, 976–983. [Google Scholar] [CrossRef]
- Sun, L.; Liu, Q.; Fan, J.; Li, X.; Zhuang, Y. Purification and characterization of peptides inhibiting MMP-1 activity with C-terminate of Gly-Leu from simulated gastrointestinal digestion hydrolysates of tilapia (Oreochromis niloticus) skin gelatin. J. Agric. Food Chemi. 2018, 66, 593–601. [Google Scholar]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Wu, B.; Yan, M.; Hou, H.; Zhuang, Y. Antihypertensive Effect in Vivo of QAGLSPVR and Its Transepithelial Transport Through the Caco-2 Cell Monolayer. Mar. Drugs 2019, 17, 288. https://doi.org/10.3390/md17050288
Sun L, Wu B, Yan M, Hou H, Zhuang Y. Antihypertensive Effect in Vivo of QAGLSPVR and Its Transepithelial Transport Through the Caco-2 Cell Monolayer. Marine Drugs. 2019; 17(5):288. https://doi.org/10.3390/md17050288
Chicago/Turabian StyleSun, Liping, Beiyi Wu, Mingyan Yan, Hu Hou, and Yongliang Zhuang. 2019. "Antihypertensive Effect in Vivo of QAGLSPVR and Its Transepithelial Transport Through the Caco-2 Cell Monolayer" Marine Drugs 17, no. 5: 288. https://doi.org/10.3390/md17050288
APA StyleSun, L., Wu, B., Yan, M., Hou, H., & Zhuang, Y. (2019). Antihypertensive Effect in Vivo of QAGLSPVR and Its Transepithelial Transport Through the Caco-2 Cell Monolayer. Marine Drugs, 17(5), 288. https://doi.org/10.3390/md17050288