Evaluation of the Inhibitory Potential of Casuarictin, an Ellagitannin Isolated from White Mangrove (Laguncularia racemosa) Leaves, on Snake Venom Secretory Phospholipase A2

 , and
, and 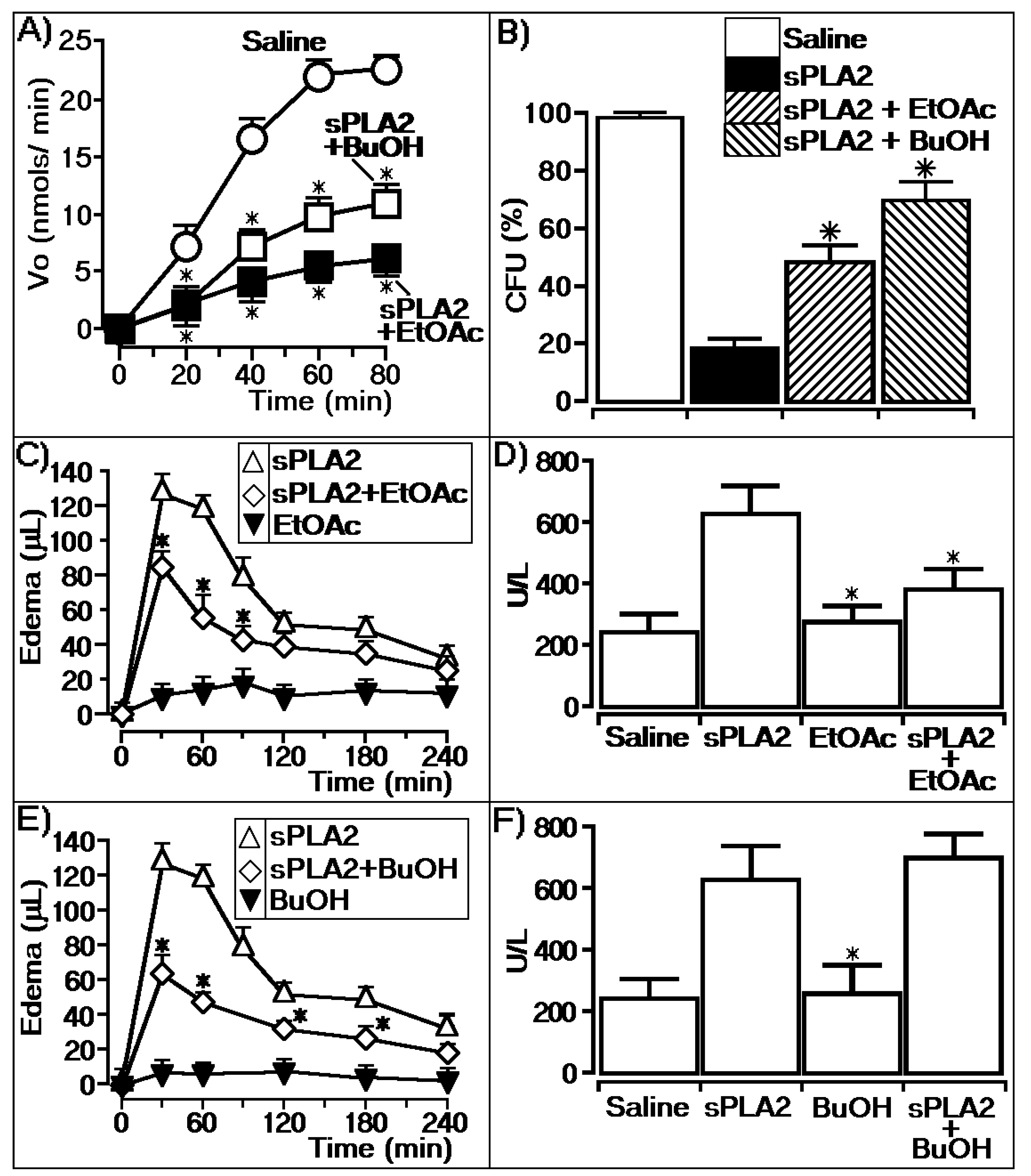{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
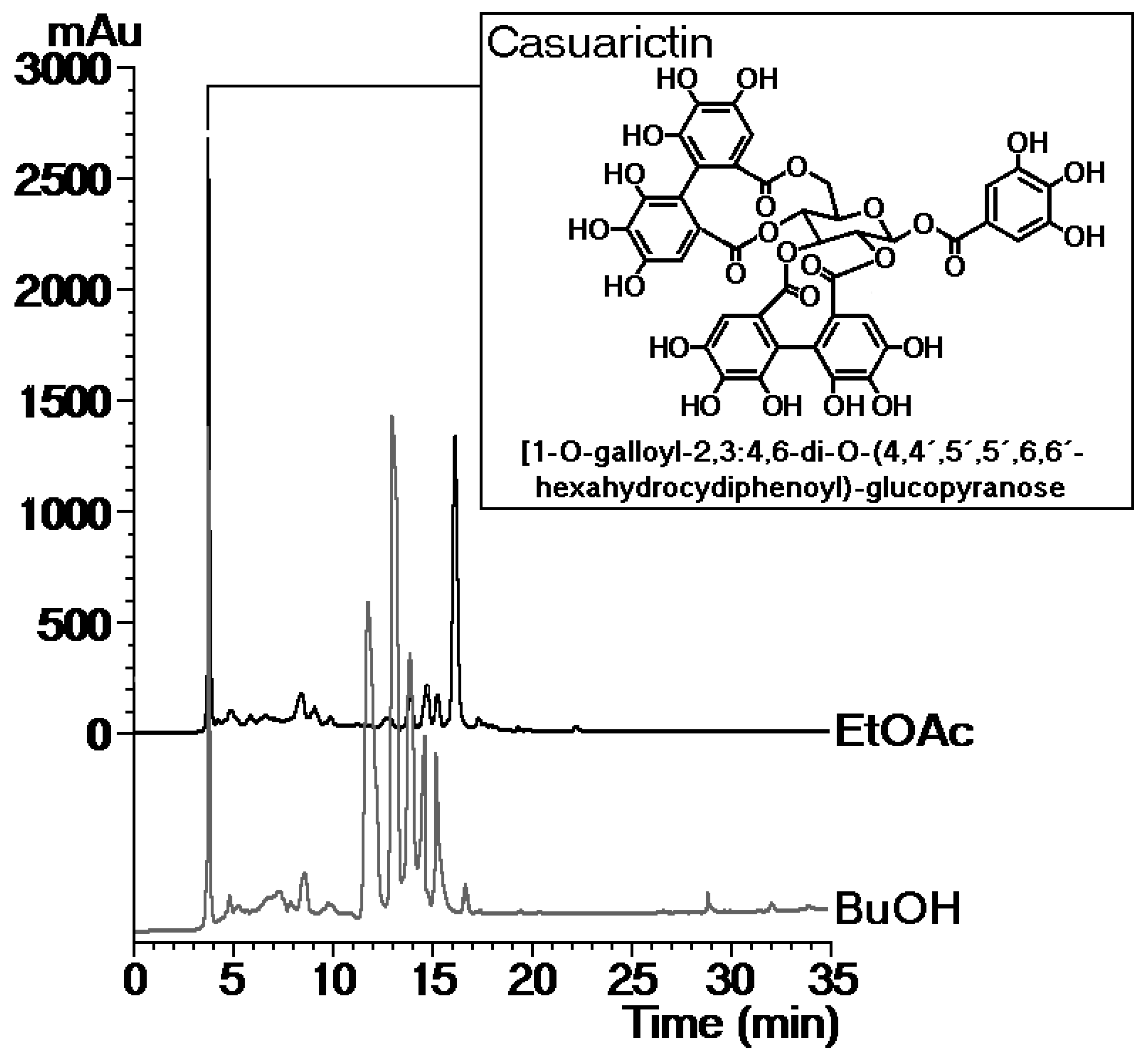
2.1. Isolation, Structural Characterization, and Biological and Pharmacological Trials of Casuarictin, a Tannin from Laguncularia racemosa Leaves
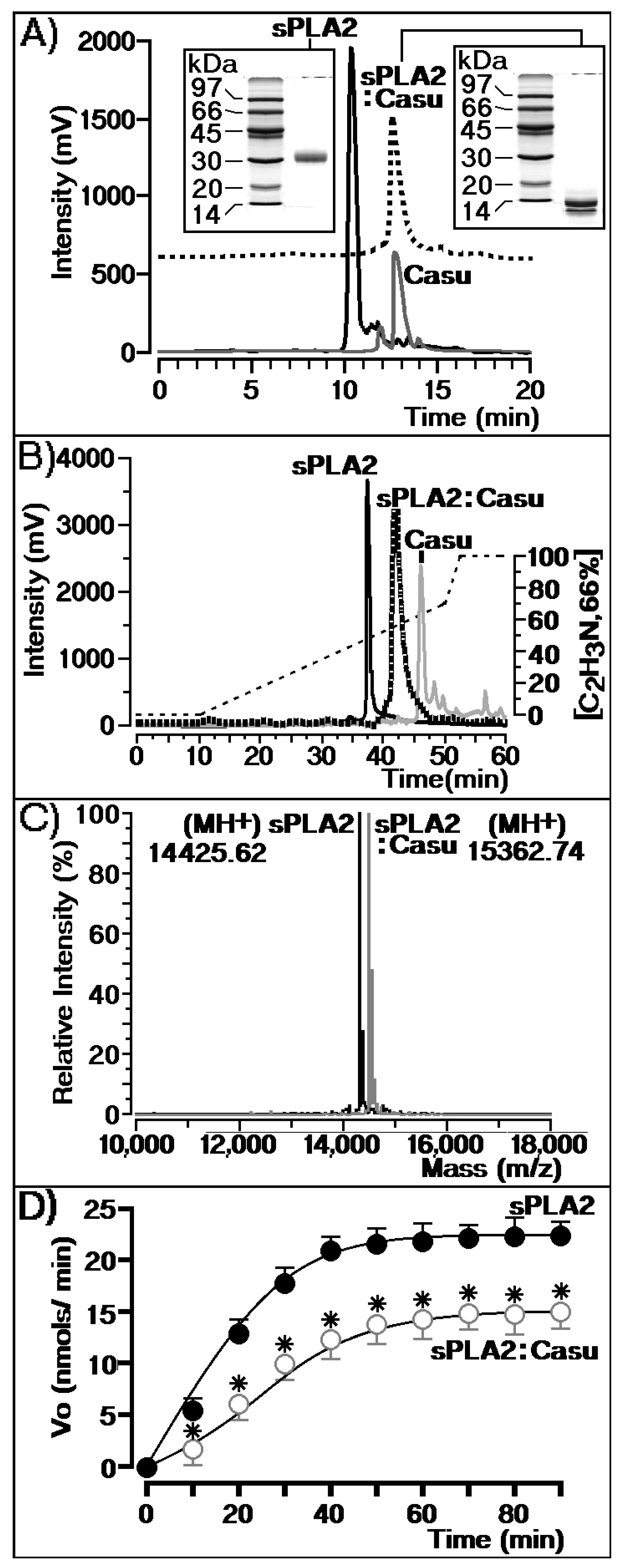
2.2. Biochemical Characterization of Tannin and sPLA2 Interaction
2.3. Effects of Casu on the Enzymatic and Pharmacological Activities of sPLA2
2.4. Antimicrobial Effects of Casu Tannin
3. Discussion
4. Materials and Methods
4.1. Materials and Animals
4.2. Plant Material, Extraction, Isolation, and Identification of Compounds
4.3. Purification of sPLA2 from Commercial Crotoxin
4.4. Biochemical Analysis
4.4.1. Treatment of sPLA2 with Extracts and Mangrove Tannin
4.4.2. Mass Spectrometry
4.4.3. Phospholipase A2 Assay
4.5. Pharmacological Assay and Antimicrobial Assays
4.5.1. Paw Edema
4.5.2. Evaluation of Myonecrosis
4.5.3. Electron Microscopy
4.5.4. Antibacterial Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Oliveira, S.C.; Fonseca, F.V.; Antunes, E.; Camargo, E.A.; Morganti, R.P.; Aparício, R.; Toyama, D.O.; Beriam, L.O.; Nunes, E.V.; Cavada, B.S.; et al. Modulation of the pharmacological effects of enzymatically-active PLA2 by BTL-2, an isolectin isolated from the Bryothamnion triquetrum red alga. Bmc Biochem. 2008, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Sales, T.A.; Marcussi, S.; da Cunha, E.; Kuca, K.; Ramalho, T.C. Can Inhibitors of Snake Venom Phospholipases A₂ Lead to New Insights into Anti-Inflammatory Therapy in Humans? Theoretical Study. Toxins Basel 2017, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, V.O.; Picolo, G.; Fernandes, C.; Fontes, M.; Cury, Y. Secreted Phospholipases A₂ from Animal Venoms in Pain and Analgesia. Toxins 2017, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Toyama, D.O.; Ferreira, M.J.; Romoff, P.; Fávero, O.A.; Gaeta, H.H.; Toyama, M.H. Effect of chlorogenic acid (5-Caffeoylquinic Acid) isolated from Baccharis oxyodonta on the structure and pharmacological activities of secretory phospholipase A2 from Crotalus durissus terrificus. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, C.A.; de Oliveira, S.C.; Diz Filho, E.B.; Fonseca, F.V.; Baldissera, L., Jr.; Antunes, E.; Ximenes, R.M.; Monteiro, H.S.; Rabello, M.M.; Hernandes, M.Z.; et al. Quercetin as an inhibitor of snake venom secretory phospholipase A2. Chem. Biol. Interact. 2011, 189, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.L.G.; Chaves, F.O.; Estrada, G.C.D.; Fernandez, V. Mangrove forests associated with salt flats: A case study from southeast Brazil. Braz. J. Oceanogr. 2017, 65, 102–115. [Google Scholar] [CrossRef]
- Saranraj, P.; Sujitha, D. Mangrove Medicinal Plants: A Review. Am. Eurasian J. Toxicol. Sci. 2015, 7, 146–156. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Gaeta, H.H.; Belchor, M.N.; Ferreira, M.J.P.; Pinho, M.V.; Toyama, D.; Toyama, M.H. Evaluation of Potential Thrombin Inhibitors from the White Mangrove (Laguncularia racemosa (L.) C.F. Gaertn.). Mar. Drugs 2015, 13, 4505–4519. [Google Scholar] [CrossRef] [PubMed]
- Ray, T. Customary of mangrove tree as a folk medicine among the sundarban resource collection. Impact 2014, 2, 43–48. [Google Scholar]
- Arora, K.; Nagpal, M.; Jain, U.; Jain, S. Mangroves: A novel gregarious phyto medicine for diabetes. Int. J. Res. Dev. Pharm. Life Sci. 2014, 3, 1244–1257. [Google Scholar]
- Toyama, D.d.O.; Gaeta, H.H.; de Pinho, M.V.; Ferreira, M.J.; Romoff, P.; Matioli, F.F.; Magro, A.J.; Fontes, M.R.; Toyama, M.H. An evaluation of 3-rhamnosylquercetin, a glycosylated form of quercetin, against the myotoxic and edematogenic effects of sPLA2 from Crotalus durissus terrificus. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Aljaghthm, O.; Heba, H.; Zeid, A. Bioactive Compounds Extracted from Mangrove Plants (Avicennia marina and Rhizophora mucronata): An Overview. Pathophysiology 2018. [Google Scholar] [CrossRef]
- Smerglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. British J. Pharmacol. 2016, 174, 1244–1262. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.R. Fauna used in popular medicine in Northeast Brazil. J. Ethnobiol Ethnomed 2009, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Moteriya, P.; Dalsaniya, A.; Chanda, S. Antioxidant and antimicrobial activity of a mangrove plant Avicennia marina (Forsk.). J. Coast. Life Med. 2015, 3, 713–717. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Ashida, M.; Yazaki, K. Tannins of Casuarina and Stachyurus species. Part 1. Structures of pendunculagin, casuarictin, strictinin, casuarinin, casuariin, and stachyurin. J. Chem. Soc. Perkin Trans. I 1983, 1765–1772. [Google Scholar] [CrossRef]
- Okuda, T.; Ito, H. Tannins of Constant Structure in Medicinal and Food Plants-Hydrolyzable Tannins and Polyphenols Related to Tannins. Molecules 2011, 16, 2191–2217. [Google Scholar] [CrossRef]
- Naumann, H.D.; Tedeschi, L.O.; Zeller, W.E.; Huntley, N.F. The role of condensed tannins in ruminant animal production: Advances, limitations and future directions. Revista Brasileira de Zootecnia 2017, 46, 929–949. [Google Scholar] [CrossRef]
- Kodikara, K.A.S.; Jayatissa, L.P.; Huxham, M.; Dahdouh-Guebas, F.; Koedam, N. The effects of salinity on growth and survival of mangrove seedlings changes with age. Acta Bot. Bras. 2017, 32, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Silveira, A.G.; Santini, E.J.; Kulczynski, S.M.; Trevisan, R.; Wastowski, A.D.; Gatto, D.A. Tannic extract potential as natural wood preservative of Acacia mearnsii. An. Acad. Bras. Ciênc. 2017, 89, 3031–3038. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic engineering of anthocyanins and condensed tannins in plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.M.; Alkhafadji, L.; Stringano, E.; Nilsson, S.; Mueller-Harvey, I.; Udén, P. Relationship between condensed tannin structures and their ability to precipitate feed proteins in the rumen. J. Sci. Food Agric. 2014, 94, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Calixto, J.B.; Nicolau, M.; Rae, G.A. Pharmacological Actions of Tannic Acid. I. Effects on Isolated Smooth and Cardiac Muscles and on Blood Pressure. Planta Med. 1986, 52, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Yamada, P.; Ono, T.; Shigemori, H.; Han, J.; Isoda, H. Inhibitory effect of tannins from galls of Carpinus tschonoskii on the degranulation of RBL-2H3 Cells. Cytotechnology 2012, 64, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Moltke, I.; Grarup, N.; Racimo, F.; Bjerregaard, P.; Jørgensen, M.E.; Korneliussen, T.S.; Gerbault, P.; Skotte, L.; Linneberg, A.; et al. Greenlandic Inuit show genetic signatures of diet and climate adaptation. Science 2015, 349, 1343–1347. [Google Scholar] [CrossRef] [PubMed]
- Williams, C. Medicinal Plants in Australia Volume 2: Gums, Resins, Tannin and Essential Oils; Rosenberg Publishing: Dural, NSW, Australia, 2011. [Google Scholar]
- Santos, M.L.; Toyama, D.O.; Oliveira, S.C.; Cotrim, C.A.; Diz-Filho, E.B.; Fagundes, F.H.; Soares, V.C.; Aparicio, R.; Toyama, M.H. Modulation of the pharmacological activities of secretory phospholipase A2 from Crotalus durissus cascavella induced by naringin. Molecules 2011, 16, 738–761. [Google Scholar] [CrossRef] [PubMed]
- Shridas, P.; Nancy, R.; Webb, N.R. Diverse Functions of Secretory Phospholipases A2. Adv. Vasc. Med. 2014, 11, 2014. [Google Scholar] [CrossRef]
- Petrova, S.D.; Atanasov, V.N.; Balashey, K. Vipoxin and Its Components: Structure–Function Relationship. Adv. Protein Chem. Struct. Biol. 2012, 87, 117–153. [Google Scholar] [CrossRef] [PubMed]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory phospholipase A2 enzymes as pharmacological targets for treatment of disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Yarla, N.S.; Bishayee, A.; Vadlakonda, L.; Chintala, R.; Duddukuri, G.R.; Reddanna, P.; Dowluru, K.S. Phospholipase A2 Isoforms as Novel Targets for Prevention and Treatment of Inflammatory and Oncologic Diseases. Curr. Drug. Targets. 2016, 17, 1940–1962. [Google Scholar] [CrossRef]
- Pereañez, J.A.; Patiño, A.C.; Núñez, V.; Osorio, E. The biflavonoid morelloflavone inhibits the enzymatic and biological activities of a snake venom phospholipase A2. Chem. Biol. Interact. 2014, 220, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.L.R.; Thomas, G.; Barbosa Filho, J.M. Anti-inflammatory actions of tannins isolated from the bark of Anacardium occidentale L. J. Ethnopharmacol. 1985, 13, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Formagio, A.S.; Volobuff, C.R.; Santiago, M.; Cardoso, C.A.; Vieira, M.d.C.; Valdevina, P.Z. Evaluation of Antioxidant Activity, Total Flavonoids, Tannins and Phenolic Compounds in Psychotria Leaf Extracts. Antioxidants 2014, 3, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Ding, Y.S.; Zhang, Y.; Chen, J.B.; Cui, B.S.; Bai, J.Y.; Lin, M.B.; Hou, Q.; Zhang, P.C.; Li, S. Anti-inflammatory Hydrolyzable Tannins from Myricaria bracteata. J. Nat. Prod. 2015, 78, 1015–1025. [Google Scholar] [CrossRef]
- Stachowska, E.; Baśkiewicz-Masiuk, M.; Dziedziejko, V.; Gutowska, I.; Baranowska-Bosiacka, I.; Marchlewicz, M.; Dołegowska, B.; Wiszniewska, B.; Machaliński, B.; Chlubek, D. Conjugated linoleic acid increases intracellular ROS synthesis and oxygenation of arachidonic acid in macrophages. Nutrition 2008, 24, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Benavente-García, O.; Castillo, J. Update on uses and properties of Citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, C.F.B.; Ferreira, M.J.P.; Belchor, M.N.; Costa, C.R.C.; Novaes, D.P.; dos Santos Junior, A.B.; Tamayose, C.I.; Pinho, M.V.T.; de Oliveira, M.A.; Toyama, M.H. Evaluation of the Inhibitory Potential of Casuarictin, an Ellagitannin Isolated from White Mangrove (Laguncularia racemosa) Leaves, on Snake Venom Secretory Phospholipase A2. Mar. Drugs 2019, 17, 403. https://doi.org/10.3390/md17070403
Rodrigues CFB, Ferreira MJP, Belchor MN, Costa CRC, Novaes DP, dos Santos Junior AB, Tamayose CI, Pinho MVT, de Oliveira MA, Toyama MH. Evaluation of the Inhibitory Potential of Casuarictin, an Ellagitannin Isolated from White Mangrove (Laguncularia racemosa) Leaves, on Snake Venom Secretory Phospholipase A2. Marine Drugs. 2019; 17(7):403. https://doi.org/10.3390/md17070403
Chicago/Turabian StyleRodrigues, Caroline Fabri Bittencourt, Marcelo José Pena Ferreira, Mariana Novo Belchor, Caroline R. C. Costa, Danielle P. Novaes, Adeilso Bispo dos Santos Junior, Cinthia I. Tamayose, Marcus Vinícius Terashima Pinho, Marcos Antonio de Oliveira, and Marcos Hikari Toyama. 2019. "Evaluation of the Inhibitory Potential of Casuarictin, an Ellagitannin Isolated from White Mangrove (Laguncularia racemosa) Leaves, on Snake Venom Secretory Phospholipase A2" Marine Drugs 17, no. 7: 403. https://doi.org/10.3390/md17070403
APA StyleRodrigues, C. F. B., Ferreira, M. J. P., Belchor, M. N., Costa, C. R. C., Novaes, D. P., dos Santos Junior, A. B., Tamayose, C. I., Pinho, M. V. T., de Oliveira, M. A., & Toyama, M. H. (2019). Evaluation of the Inhibitory Potential of Casuarictin, an Ellagitannin Isolated from White Mangrove (Laguncularia racemosa) Leaves, on Snake Venom Secretory Phospholipase A2. Marine Drugs, 17(7), 403. https://doi.org/10.3390/md17070403