Structure-Function Elucidation of a New α-Conotoxin, MilIA, from Conus milneedwardsi

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of a Novel α-Conotoxin in C. milneedwardsi Venom
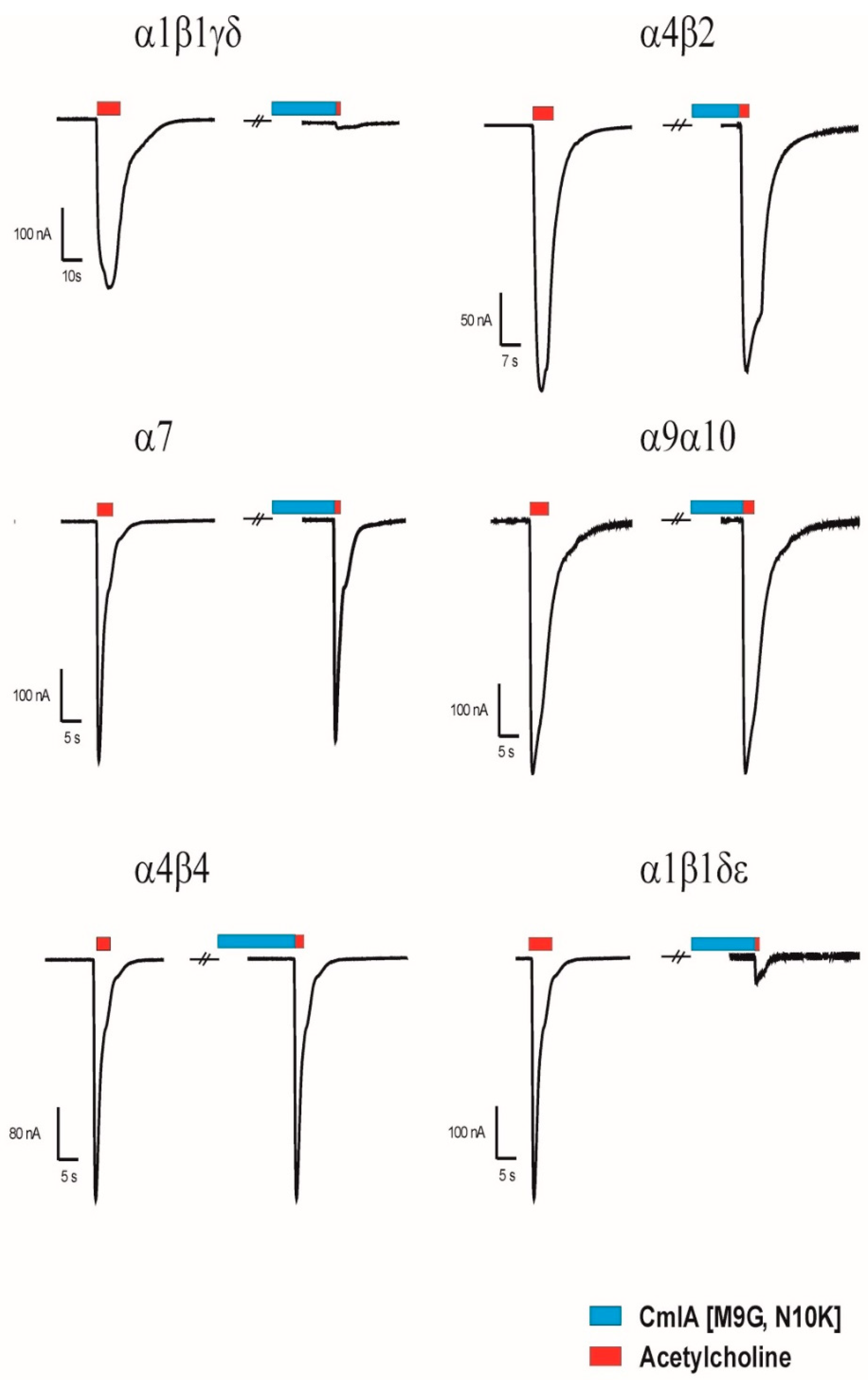
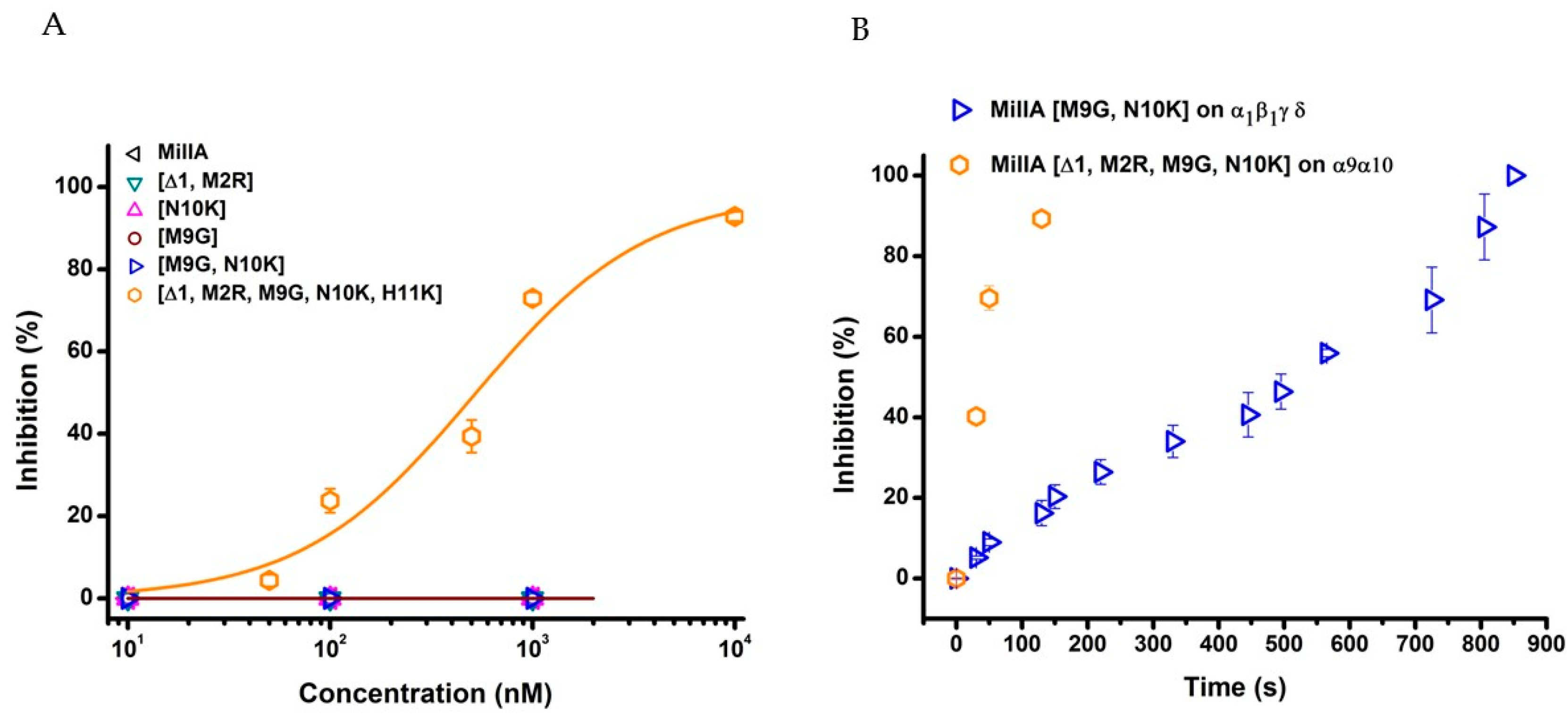
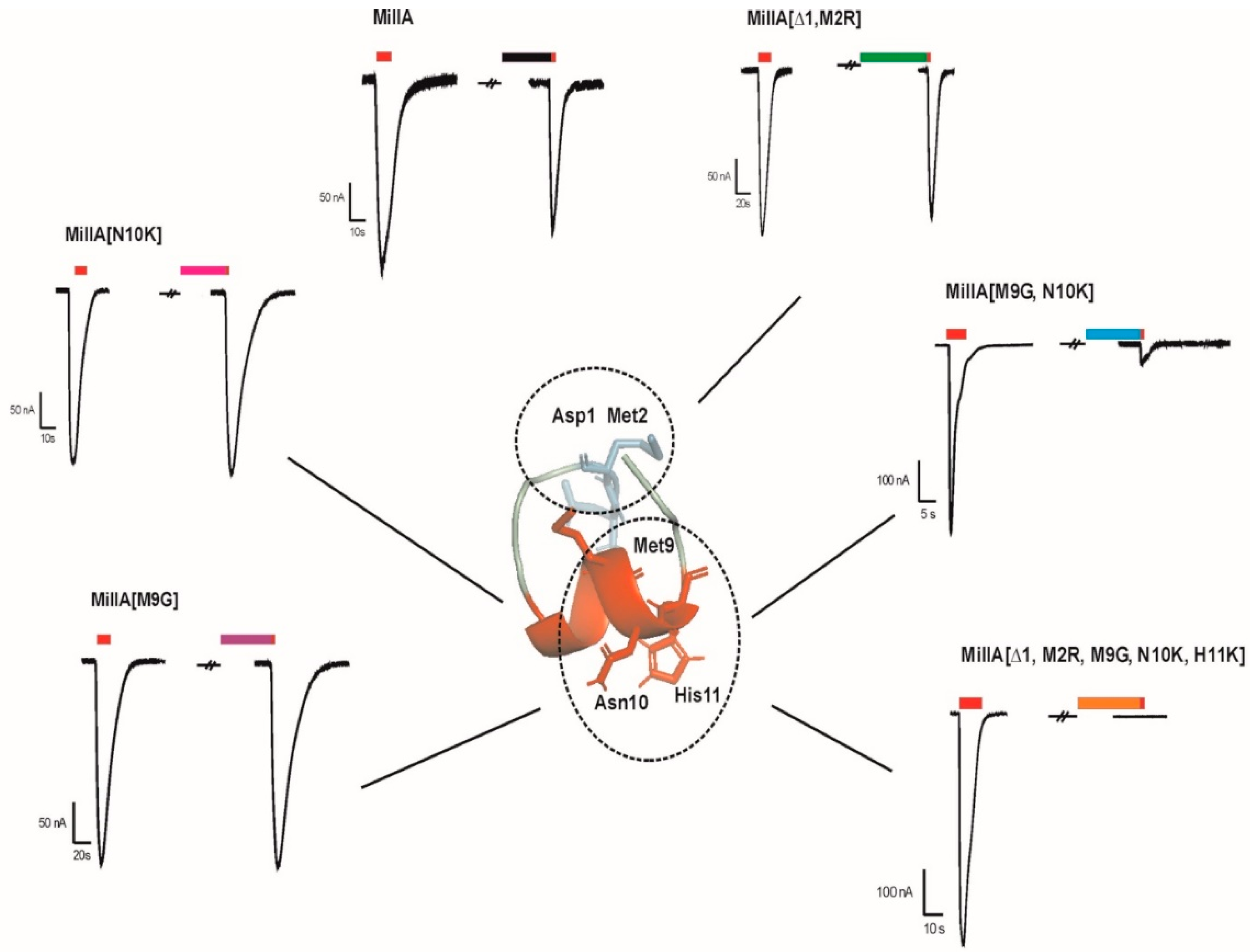
2.2. Functional Characterisation of MilIA and its Analogues
Activity of MilIA and its Analogues on Muscle-Type nAChRs
3. Discussion
4. Materials and Methods
4.1. Crude Venom Extraction from Conus milneedwardsi
4.2. Peptide Purification
4.3. Peptide Sequencing
4.4. Mass Spectrometry Analysis
4.5. Peptide Synthesis and Folding
4.6. Functional Characterization
4.6.1. Expression of Voltage-Gated ion Channels in Xenopus Laevis Oocytes
4.6.2. Electrophysiological Recordings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fu, Y.; Li, C.; Dong, S.; Wu, Y.; Zhangsun, D.; Luo, S. Discovery Methodology of Novel Conotoxins from Conus Species. Mar. Drugs 2018, 16, 417. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.; Lewis, R.J. Neuronal Nicotinic Acetylcholine Receptor Modulators from Cone Snails. Mar. Drugs 2018, 16, 208. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, J.; Dutertre, S. Alpha-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.W.; Marquart, L.A.; Phillips, P.D.; McDougal, O.M. Mutagenesis of alpha-Conotoxins for Enhancing Activity and Selectivity for Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.A.; Mari, F.; Lewis, R.J. Accelerated proteomic visualization of individual predatory venoms of Conus purpurascens reveals separately evolved predation-evoked venom cabals. Sci. Rep. 2018, 8, 330. [Google Scholar] [CrossRef]
- Kumar, P.S.; Kumar, D.S.; Umamaheswari, S. A perspective on toxicology of Conus venom peptides. Asian Pac. J. Trop. Med. 2015, 8, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Thapa, P.; Zhang, R.Y.; Menon, V.; Bingham, J.P. Native chemical ligation: A boon to peptide chemistry. Molecules 2014, 19, 14461–14483. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Brogan, S.E.; Olivera, B.M. Pain therapeutics from cone snail venoms: From Ziconotide to novel non-opioid pathways. J. Proteom. 2019, 190, 12–20. [Google Scholar] [CrossRef]
- Essack, M.; Bajic, V.B.; Archer, J.A. Conotoxins that confer therapeutic possibilities. Mar. Drugs 2012, 10, 1244–1265. [Google Scholar] [CrossRef]
- Bertrand, D.; Terry, A.V., Jr. The wonderland of neuronal nicotinic acetylcholine receptors. Biochem. Pharmacol. 2018, 151, 214–225. [Google Scholar] [CrossRef]
- Hone, A.J.; McIntosh, J.M. Nicotinic acetylcholine receptors in neuropathic and inflammatory pain. FEBS Lett. 2018, 592, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; Talley, T.T.; Bobango, J.; Melo, C.H.; Hararah, F.; Gajewiak, J.; Christensen, S.; Harvey, P.J.; Craik, D.J.; McIntosh, J.M. Molecular determinants of alpha-conotoxin potency for inhibition of human and rat alpha6beta4 nicotinic acetylcholine receptors. J. Biol. Chem. 2018, 293, 17838–17852. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.G.; Qian, Y.H. Alpha 7 nicotinic acetylcholine receptor and its effects on Alzheimer’s disease. Neuropeptides 2019, 73, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Yu, R.; Jin, A.H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef] [PubMed]
- Groebe, D.R.; Gray, W.R.; Abramson, S.N. Determinants involved in the affinity of alpha-conotoxins GI and SI for the muscle subtype of nicotinic acetylcholine receptors. Biochemistry 1997, 36, 6469–6474. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; McIntosh, J.M. Iodo-alpha-conotoxin MI selectively binds the alpha/delta subunit interface of muscle nicotinic acetylcholine receptors. Biochemistry 2004, 43, 6656–6662. [Google Scholar] [CrossRef]
- McIntosh, J.M.; Azam, L.; Staheli, S.; Dowell, C.; Lindstrom, J.M.; Kuryatov, A.; Garrett, J.E.; Marks, M.J.; Whiteaker, P. Analogs of alpha-conotoxin MII are selective for alpha6-containing nicotinic acetylcholine receptors. Mol. Pharmacol. 2004, 65, 944–952. [Google Scholar] [CrossRef]
- Papineni, R.V.; Sanchez, J.U.; Baksi, K.; Willcockson, I.U.; Pedersen, S.E. Site-specific charge interactions of alpha-conotoxin MI with the nicotinic acetylcholine receptor. J. Biol. Chem. 2001, 276, 23589–23598. [Google Scholar] [CrossRef]
- Kasheverov, I.E.; Utkin, Y.N.; Tsetlin, V.I. Naturally occurring and synthetic peptides acting on nicotinic acetylcholine receptors. Curr. Pharm. Des. 2009, 15, 2430–2452. [Google Scholar] [CrossRef]
- El Hamdaoui, Y.; Wu, X.; Clark, R.J.; Giribaldi, J.; Anangi, R.; Craik, D.J.; King, G.F.; Dutertre, S.; Kaas, Q.; Herzig, V.; et al. Periplasmic Expression of 4/7 alpha-Conotoxin TxIA Analogs in E. coli Favors Ribbon Isomer Formation—Suggestion of a Binding Mode at the alpha7 nAChR. Front. Pharmacol. 2019, 10, 577. [Google Scholar] [CrossRef]
- Almquist, R.G.; Kadambi, S.R.; Yasuda, D.M.; Weitl, F.L.; Polgar, W.E.; Toll, L.R. Paralytic activity of (des-Glu1)conotoxin GI analogs in the mouse diaphragm. Int. J. Pept. Protein Res. 1989, 34, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Bren, N.; Sine, S.M. Hydrophobic pairwise interactions stabilize alpha-conotoxin MI in the muscle acetylcholine receptor binding site. J. Biol. Chem. 2000, 275, 12692–12700. [Google Scholar] [CrossRef] [PubMed]
- Mordvintsev, D.Y.; Polyak, Y.L.; Kuzmine, D.A.; Levtsova, O.V.; Tourleigh, Y.V.; Kasheverov, I.E. A model for short alpha-neurotoxin bound to nicotinic acetylcholine receptor from Torpedo californica. J. Mol. Neurosci. 2006, 30, 71–72. [Google Scholar] [CrossRef]
- Ning, J.; Li, R.; Ren, J.; Zhangsun, D.; Zhu, X.; Wu, Y.; Luo, S. Alanine-Scanning Mutagenesis of alpha-Conotoxin GI Reveals the Residues Crucial for Activity at the Muscle Acetylcholine Receptor. Mar. Drugs 2018, 16, 507. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N.; Kuch, U.; Kasheverov, I.E.; Lebedev, D.S.; Cederlund, E.; Molles, B.E.; Polyak, I.; Ivanov, I.A.; Prokopev, N.A.; Ziganshin, R.H.; et al. Novel long-chain neurotoxins from Bungarus candidus distinguish the two binding sites in muscle-type nicotinic acetylcholine receptors. Biochem. J. 2019, 476, 1285–1302. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chew, G.; Hawrot, E.; Chi, C.; Wang, C. Two potent alpha3/5 conotoxins from piscivorous Conus achatinus. Acta Biochim. Biophys. Sin. (Shanghai) 2007, 39, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Xiang, S.; Li, M. Residues Responsible for the Selectivity of alpha-Conotoxins for Ac-AChBP or nAChRs. Mar. Drugs 2016, 14, 173. [Google Scholar] [CrossRef] [PubMed]
- Inserra, M.C.; Kompella, S.N.; Vetter, I.; Brust, A.; Daly, N.L.; Cuny, H.; Craik, D.J.; Alewood, P.F.; Adams, D.J.; Lewis, R.J. Isolation and characterization of alpha-conotoxin LsIA with potent activity at nicotinic acetylcholine receptors. Biochem. Pharmacol. 2013, 86, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Zhu, X.; Xu, P.; Li, R.; Fu, Y.; Dong, S.; Zhangsun, D.; Wu, Y.; Luo, S. d-Amino Acid Substitution of alpha-Conotoxin RgIA Identifies its Critical Residues and Improves the Enzymatic Stability. Mar. Drugs 2019, 17, 142. [Google Scholar] [CrossRef]
- Pucci, L.; Grazioso, G.; Dallanoce, C.; Rizzi, L.; De Micheli, C.; Clementi, F.; Bertrand, S.; Bertrand, D.; Longhi, R.; De Amici, M.; et al. Engineering of alpha-conotoxin MII-derived peptides with increased selectivity for native alpha6beta2* nicotinic acetylcholine receptors. FASEB J. 2011, 25, 3775–3789. [Google Scholar] [CrossRef]
- Mohammadi, S.A.; Christie, M.J. Conotoxin Interactions with α9α10-nAChRs: Is the α9α10-Nicotinic Acetylcholine Receptor an Important Therapeutic Target for Pain Management? Toxins 2015, 7, 3916–3932. [Google Scholar] [CrossRef] [PubMed]
- Dutton, J.L.; Craik, D.J. alpha-Conotoxins: Nicotinic acetylcholine receptor antagonists as pharmacological tools and potential drug leads. Curr. Med. Chem. 2001, 8, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Kompella, S.N.; Adams, D.J.; Craik, D.J.; Kaas, Q. Determination of the alpha-conotoxin Vc1.1 binding site on the alpha9alpha10 nicotinic acetylcholine receptor. J. Med. Chem. 2013, 56, 3557–3567. [Google Scholar] [CrossRef] [PubMed]
- Zouridakis, M.; Papakyriakou, A.; Ivanov, I.A.; Kasheverov, I.E.; Tsetlin, V.; Tzartos, S.; Giastas, P. Crystal Structure of the Monomeric Extracellular Domain of alpha9 Nicotinic Receptor Subunit in Complex With alpha-Conotoxin RgIA: Molecular Dynamics Insights into RgIA Binding to alpha9alpha10 Nicotinic Receptors. Front. Pharmacol. 2019, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Bhimani, R.; Anderson, L. Clinical understanding of spasticity: Implications for practice. Rehabil. Res. Pract. 2014, 2014, 279175. [Google Scholar] [CrossRef]
- Peigneur, S.; Cheneval, O.; Maiti, M.; Leipold, E.; Heinemann, S.H.; Lescrinier, E.; Herdewijn, P.; De Lima, M.E.; Craik, D.J.; Schroeder, C.I.; et al. Where cone snails and spiders meet: Design of small cyclic sodium-channel inhibitors. FASEB J. 2019, 33, 3693–3703. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CmIA | -DMCCH-PACMNHFN--C* | |
|---|---|---|
| CmIA [M9G] | -DMCCH-PACGNHFN--C* | |
| CmIA [N10K] | -DMCCH-PACMKHFN--C* | |
| CmIA [∆1,M2R] | --RCCH-PACMNHFN--C* | |
| CmIA [M9G,N10K] | -DMCCH-PACGKHFN--C* | |
| CmIA [∆1,M2R,M9G,N10K,H11K] | --RCCH-PACGKKFN--C* | |
| GI | --ECCN-PACGRHYS--C* | |
| MI | -GRCCH-PACGKNYS--C* | |
| SI | --ICCN-PACGPKYS--C* | |
| RgIA | --GCCSDPRCRYR----CR | |
| Vc1.1 | --GCCSDPRCNYSHPEIC* | |
| MII | --GCCSNPVCHLEHSNLC* | |
| PIA | RDPCCSNPVCTVHNPQIC* | |
| LsIA | -SGCCSNPACRVNNPNIC* |
| IC50 | α1β1δε (nM) | α1β1γδ (nM) | α2β4 (nM) | α4β2 (nM) | α4β4 (nM) | α7 (nM) | α9α10 (nM) |
|---|---|---|---|---|---|---|---|
| MilIA | 11,180 ± 78,891 | 13,130 ± 1125 | >104 | >104 | >104 | >104 | >104 |
| MilIA [M9G] | >105 | 577 ± 79 | >104 | >104 | >104 | >104 | >104 |
| MilIA [N10K] | >105 | 3842 ± 148 | >104 | >104 | >104 | >104 | >104 |
| MilIA [∆1,M2R] | 19,550 ± 50 | 18,012 ± 1430 | >104 | >104 | >104 | >104 | >104 |
| MilIA [∆1,M2R, M9G, N10K, H11K] | 25 ± 1 | 16 ± 1 | >104 | >104 | >104 | >104 | 511 ± 195 |
| MilIA [M9G, N10K] | 149 ± 39 | 178 ± 57 | >104 | >104 | >104 | >104 | >104 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peigneur, S.; Devi, P.; Seldeslachts, A.; Ravichandran, S.; Quinton, L.; Tytgat, J. Structure-Function Elucidation of a New α-Conotoxin, MilIA, from Conus milneedwardsi. Mar. Drugs 2019, 17, 535. https://doi.org/10.3390/md17090535
Peigneur S, Devi P, Seldeslachts A, Ravichandran S, Quinton L, Tytgat J. Structure-Function Elucidation of a New α-Conotoxin, MilIA, from Conus milneedwardsi. Marine Drugs. 2019; 17(9):535. https://doi.org/10.3390/md17090535
Chicago/Turabian StylePeigneur, Steve, Prabha Devi, Andrea Seldeslachts, Samuthirapandian Ravichandran, Loïc Quinton, and Jan Tytgat. 2019. "Structure-Function Elucidation of a New α-Conotoxin, MilIA, from Conus milneedwardsi" Marine Drugs 17, no. 9: 535. https://doi.org/10.3390/md17090535
APA StylePeigneur, S., Devi, P., Seldeslachts, A., Ravichandran, S., Quinton, L., & Tytgat, J. (2019). Structure-Function Elucidation of a New α-Conotoxin, MilIA, from Conus milneedwardsi. Marine Drugs, 17(9), 535. https://doi.org/10.3390/md17090535