Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Screening for AHL Degrading Bacteria
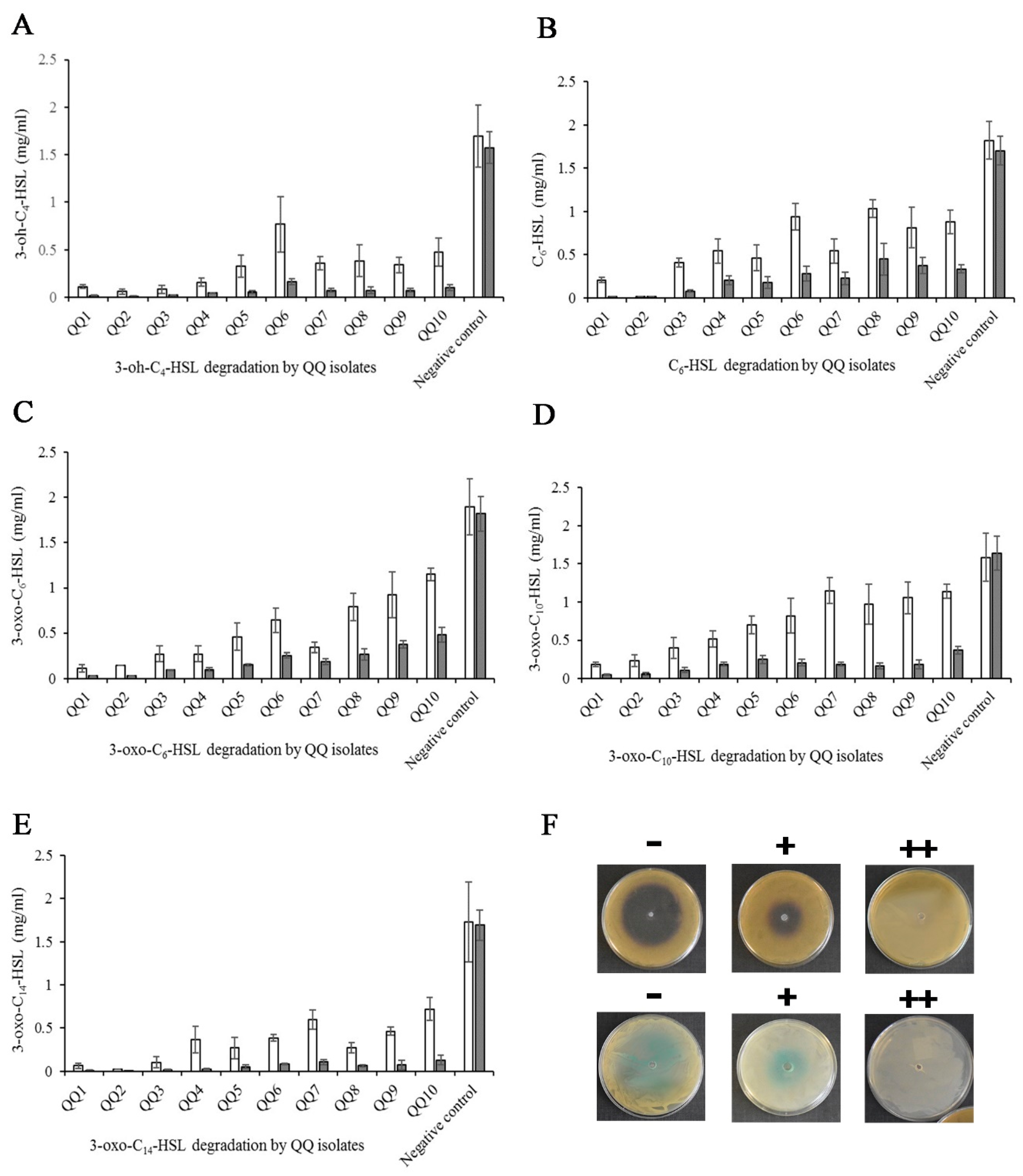
2.2. Calibration Curve and Kinetics of AHL Degradation
2.3. Identification of QQ Bacteria
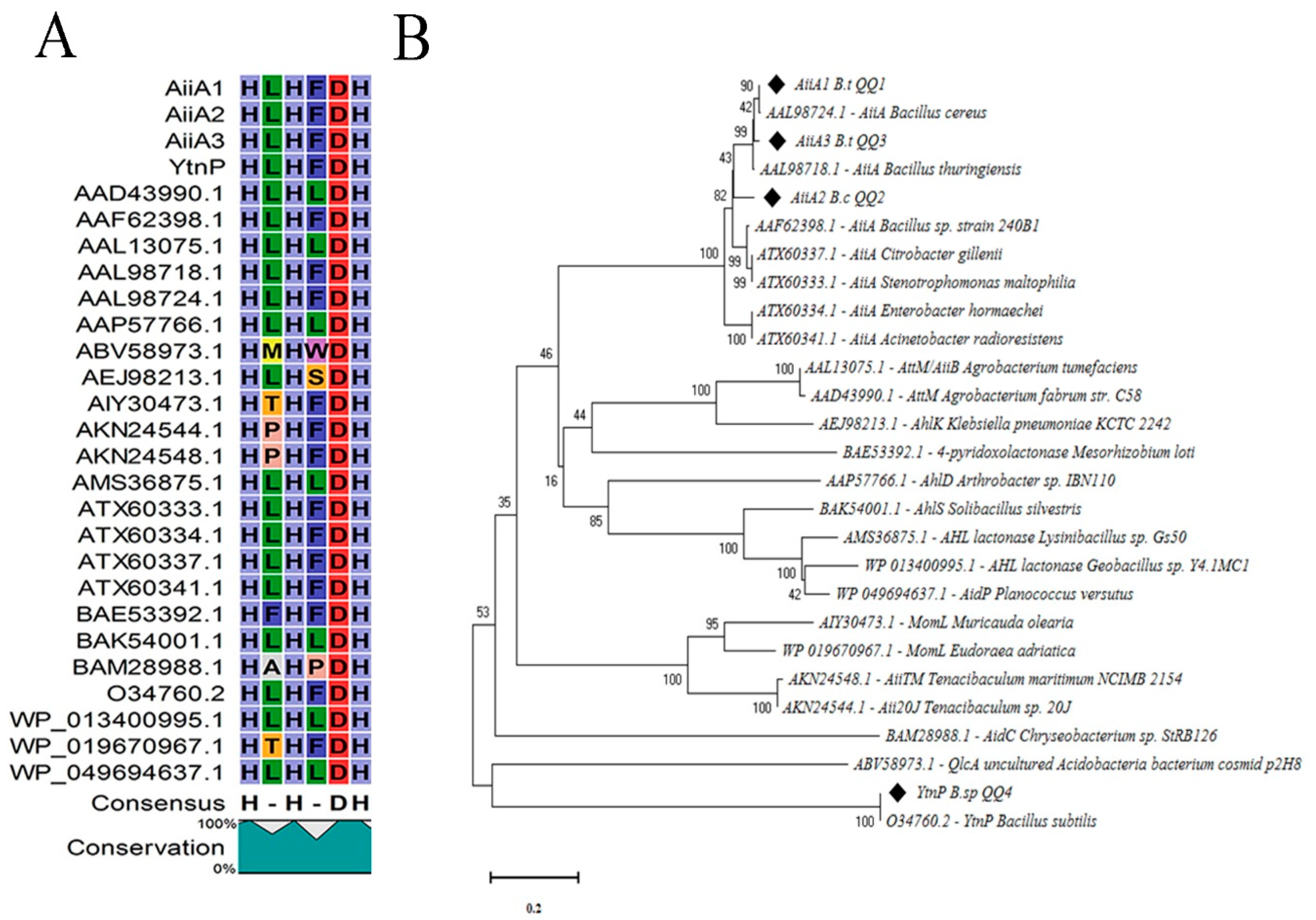
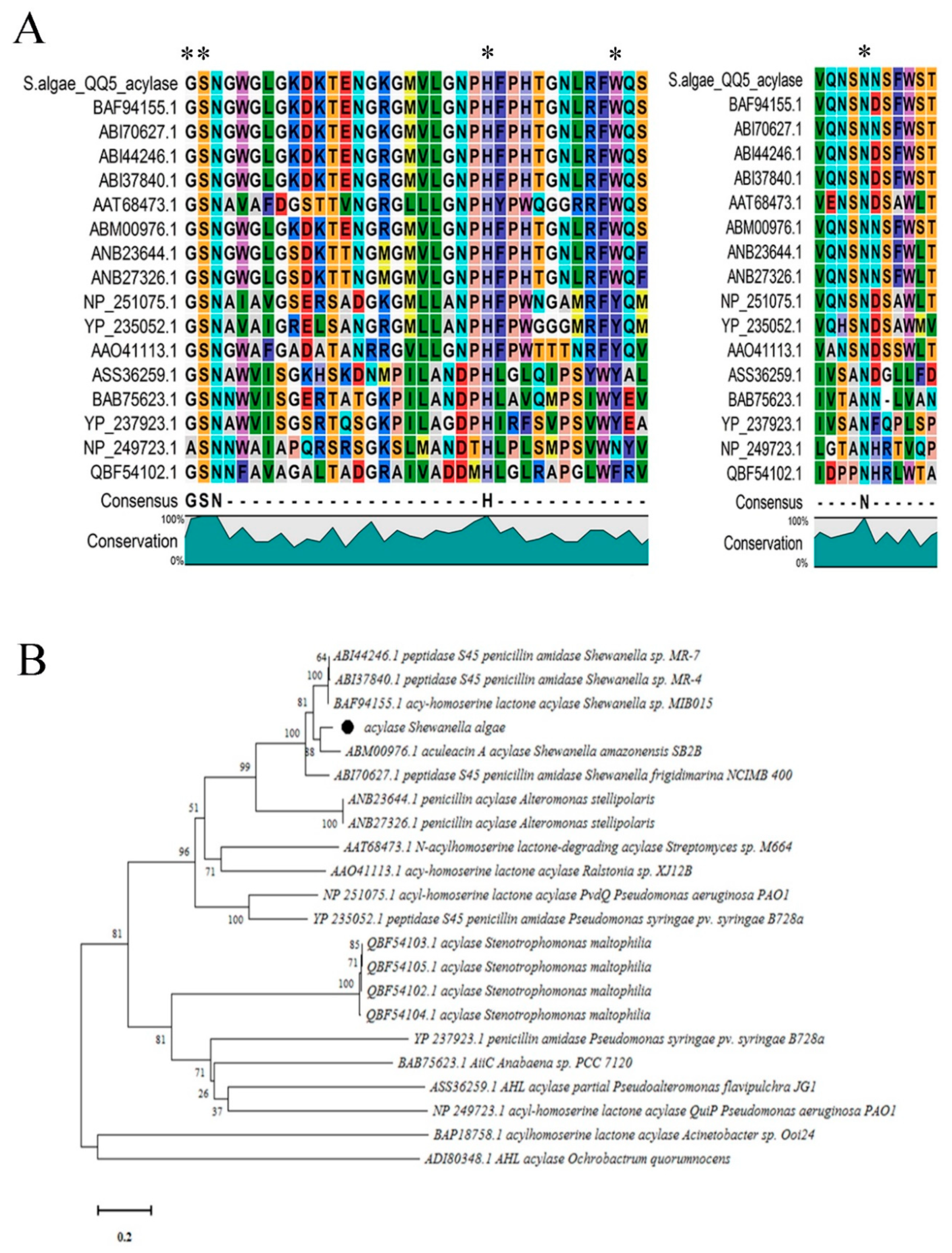
2.4. Sequence Alignment and Phylogenetic Analyses
2.5. Antimicrobial Activity of QQIs
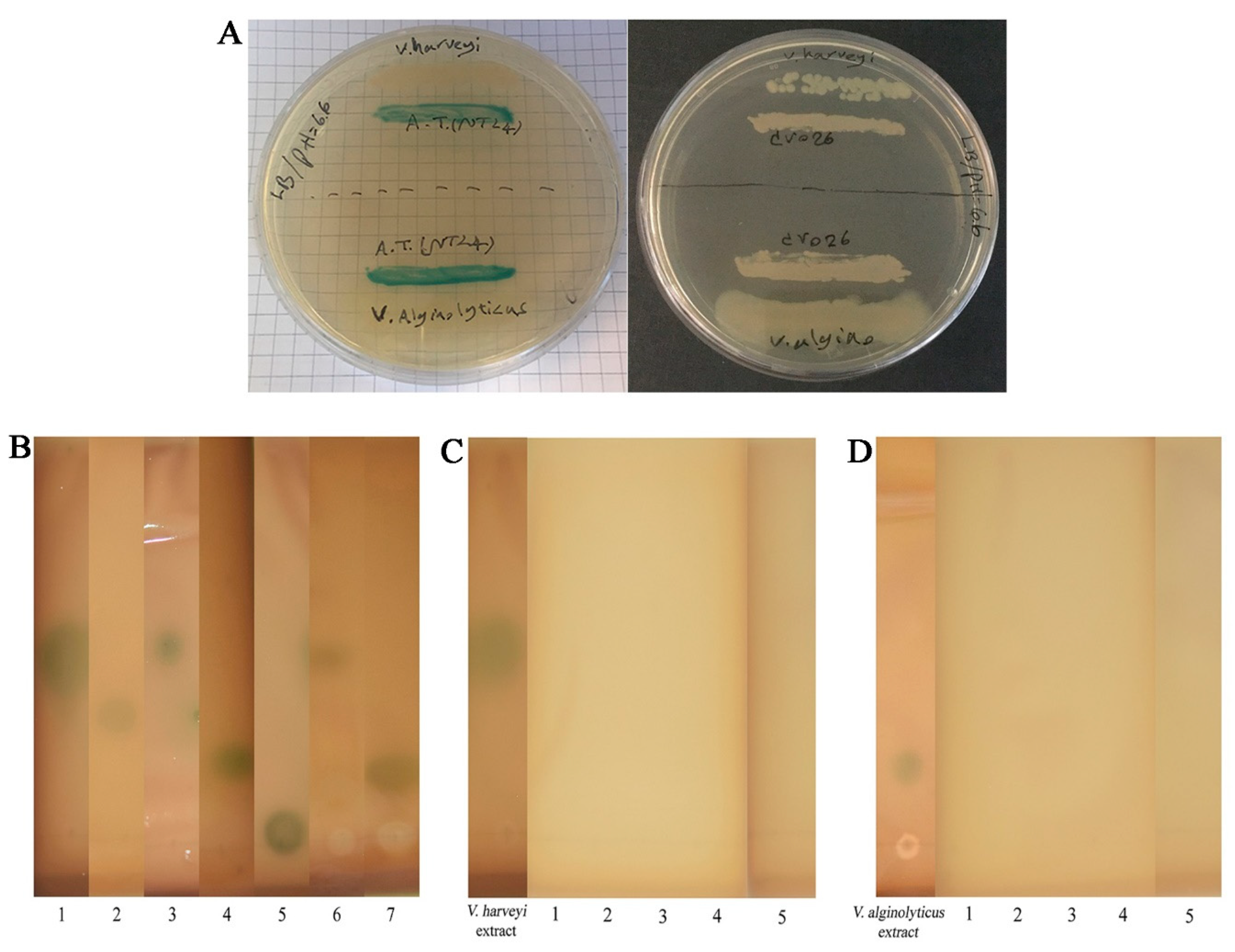
2.6. AHL Profiling and Degradation of V. harveyi and V. alginolyticus
2.7. Localization of the QQ Enzymes
2.8. Extracellular Enzymes and Sporulation
2.9. Resistance to Bile Salts and pH
2.10. Tolerance to Sodium Chloride Salt
2.11. Cell Surface Hydrophobicity
2.12. Aggregation Ability
2.13. Potential for Growth on Intestinal Mucus
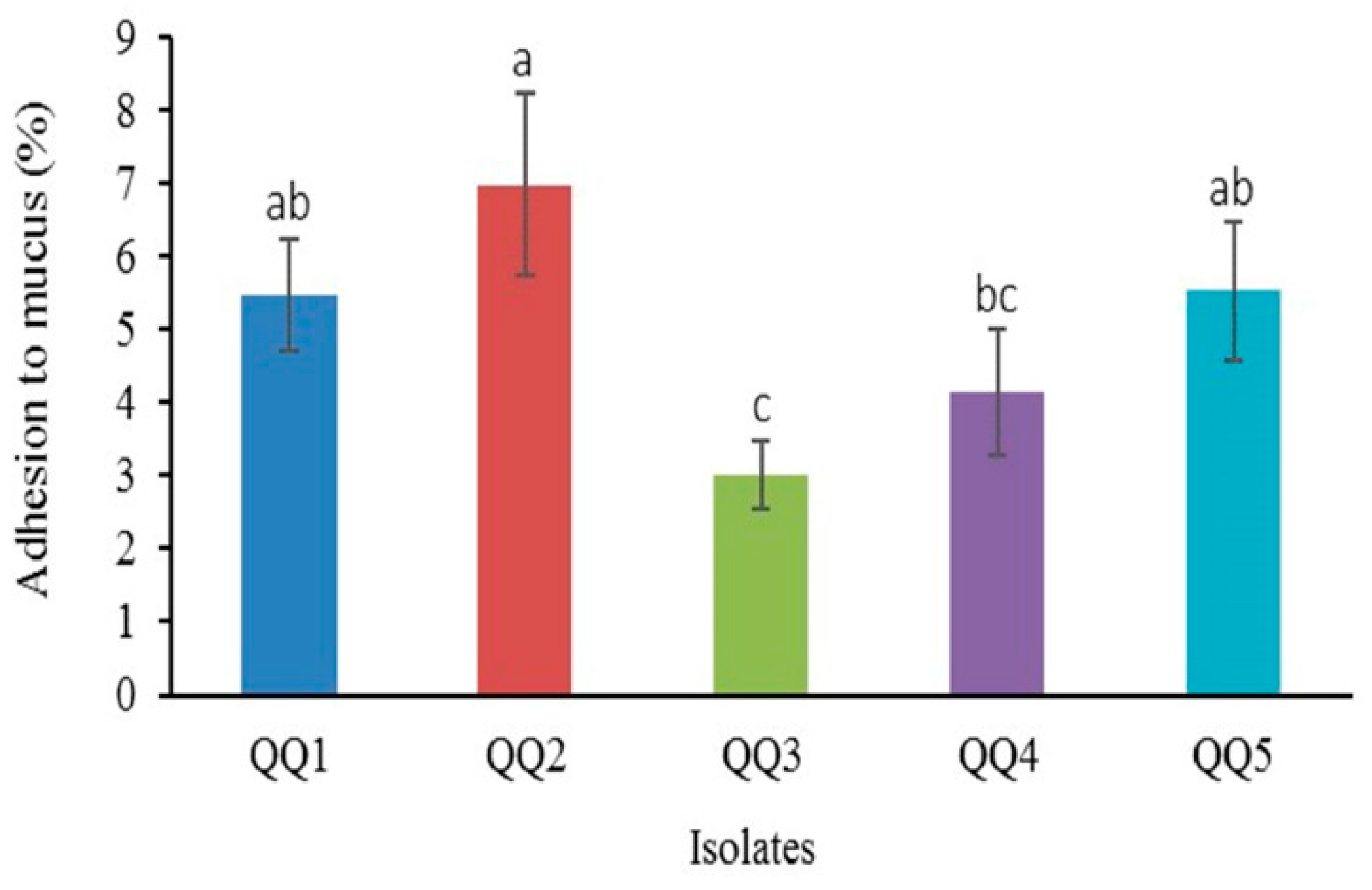
2.14. Adhesion to Intestinal Mucus
2.15. Antibiotic Susceptibility
2.16. Adverse Effects of QQIs in Barramundi Fish
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Compounds
4.2. Isolation of Bacteria from the Intestines
4.3. Screening for AHL Degrading Bacteria
4.4. Evaluation of AHL Degradation Potential using Synthetic AHLs
4.5. Identification of Signal Degrading Isolates
4.6. Sequencing of QQ-Related Genes and Phylogenetic Analysis
4.7. Antagonistic Activity in AHL Degrading Bacteria
4.8. Detection of AHL in Tested Vibrio harveyi and Vibrio alginolyticus
4.9. In vitro Assay of QQ Activity on Vibrio harveyi and Vibrio alginolyticus’ AHLs
4.10. Location of AHL degrading Enzymes in Selected Bacteria
4.11. In vitro Tests of Candidate Probiotic Bacteria
4.11.1. Enzyme Production and Spore Formation
4.11.2. Resistance to Bile Salts and pH
4.11.3. Growth at Different Salinities
4.11.4. Hydrophobicity
4.11.5. Auto-Aggregation and Co-Aggregation
4.11.6. Adherence to Intestinal Mucus
4.11.7. Growth on Intestinal Mucus
4.11.8. Antibiotic Resistance Test
4.11.9. Safety of the Selected Bacteria for Fish
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ina-Salwany, M.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in fish: A review on disease development and prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; El Basuini, M.F.; Hossain, M.S.; Nhu, T.H.; Dossou, S.; Moss, A.S. Effects of dietary supplementation of Lactobacillus rhamnosus or/and Lactococcus lactis on the growth, gut microbiota and immune responses of red sea bream, Pagrus major. Fish Shellfish. Immunol. 2016, 49, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Novriadi, R. Vibriosis in aquaculture. Omni-Akuatika 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- Vezzulli, L.; Colwell, R.R.; Pruzzo, C. Ocean warming and spread of pathogenic vibrios in the aquatic environment. Microb. Ecol. 2013, 65, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4, 1–19. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Assefa, A.; Abunna, F. Maintenance of fish health in aquaculture: Review of epidemiological approaches for prevention and control of infectious disease of fish. Vet. Med. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Fuente, M.D.L.; Miranda, C.D.; Jopia, P.; González-Rocha, G.; Guiliani, N.; Sossa, K.; Urrutia, H. Growth inhibition of bacterial fish pathogens and quorum-sensing blocking by bacteria recovered from Chilean salmonid farms. J. Aquat. Anim. Health 2015, 27, 112–122. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Saurav, K.; Costantino, V.; Venturi, V.; Steindler, L. Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef]
- Torres, M.; Reina, J.C.; Fuentes-Monteverde, J.C.; Fernández, G.; Rodríguez, J.; Jiménez, C.; Llamas, I. AHL-lactonase expression in three marine emerging pathogenic Vibrio spp. reduces virulence and mortality in brine shrimp (Artemia salina) and Manila clam (Venerupis philippinarum). PLoS ONE 2018, 13, e0195176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Shi, Y.; Zeng, G.; Gu, Y.; Chen, G.; Shi, L.; Hu, Y.; Tang, B.; Zhou, J. Acyl-homoserine lactone-based quorum sensing and quorum quenching hold promise to determine the performance of biological wastewater treatments: An overview. Chemosphere 2016, 157, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fu, K.; Wu, C.; Qin, K.; Li, F.; Zhou, L. “In-Group” Communication in Marine Vibrio: A Review of N-Acyl Homoserine Lactones-Driven Quorum Sensing. Front. Cell. Infect. Microbiol. 2018, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Purohit, A.; Johansen, J.; Hansen, H.; Leiros, H.K.; Kashulin, A.; Karlsen, C.; Smalås, A.; Haugen, P.; Willassen, N. Presence of acyl-homoserine lactones in 57 members of the Vibrionaceae family. J. Appl. Microbiol. 2013, 115, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.; Zhang, X.-H. Quorum quenching agents: Resources for antivirulence therapy. Mar. Drugs 2014, 12, 3245–3282. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Peng, M.; Tong, W.; Zhang, Q.; Song, Z. The Quorum Quenching Bacterium Bacillus licheniformis T-1 Protects Zebrafish against Aeromonas hydrophila Infection. Probiotics Antimicrob. Proteins 2019, 1–12. [Google Scholar] [CrossRef]
- Defoirdt, T.; Thanh, L.D.; Van Delsen, B.; De Schryver, P.; Sorgeloos, P.; Boon, N.; Bossier, P. N-acylhomoserine lactone-degrading Bacillus strains isolated from aquaculture animals. Aquaculture 2011, 311, 258–260. [Google Scholar] [CrossRef]
- Morohoshi, T.; Nakazawa, S.; Ebata, A.; Kato, N.; Ikeda, T. Identification and characterization of N-acylhomoserine lactone-acylase from the fish intestinal Shewanella sp. strain MIB015. Biosci. Biotechnol. Biochem. 2008, 72, 1887–1893. [Google Scholar] [CrossRef] [Green Version]
- Tinh, N.T.N.; Yen, V.H.N.; Dierckens, K.; Sorgeloos, P.; Bossier, P. An acyl homoserine lactone-degrading microbial community improves the survival of first-feeding turbot larvae (Scophthalmus maximus L.). Aquaculture 2008, 285, 56–62. [Google Scholar] [CrossRef]
- Torres, M.; Rubio-Portillo, E.; Antón, J.; Ramos-Esplá, A.A.; Quesada, E.; Llamas, I. Selection of the N-acylhomoserine lactone-degrading bacterium Alteromonas stellipolaris PQQ-42 and of its potential for biocontrol in aquaculture. Front. Microbiol. 2016, 7, 646. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, A.; Yin, H.; Chu, W. Bacillus sp. QSI-1 modulate quorum sensing signals reduce Aeromonas hydrophila level and alter gut microbial community structure in fish. Front. Cell. Infect. Microbiol. 2016, 6, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, Z.U.; Leiknes, T. Quorum Quenching Bacteria Isolated from Red Sea Sediments Reduces Biofilm Formation by Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 1354. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Lu, F.; Zhu, W.; Kang, C. Isolation and characterization of new potential probiotic bacteria based on quorum-sensing system. J. Appl. Microbiol. 2011, 110, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xi, B.; Qin, T.; Chen, K.; Xie, J. Isolation and characterization of AHL-degrading bacteria from fish and pond sediment. J. Oceanol. Limnol. 2019, 37, 1460–1467. [Google Scholar] [CrossRef]
- Torabi Delshad, S.; Soltanian, S.; Sharifiyazdi, H.; Haghkhah, M.; Bossier, P. Identification of N-acyl homoserine lactone-degrading bacteria isolated from rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2018, 125, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, K.; Shi, X.; Zhang, X.-H. Flaviramulus ichthyoenteri sp. nov., an N-acylhomoserine lactone-degrading bacterium isolated from the intestine of a flounder (Paralichthys olivaceus), and emended descriptions of the genus Flaviramulus and Flaviramulus basaltis. Int. J. Syst. Evol. Microbiol. 2013, 63, 4477–4483. [Google Scholar] [CrossRef] [Green Version]
- Sharifuzzaman, S.; Al-Harbi, A.; Austin, B. Characteristics of growth, digestive system functionality, and stress factors of rainbow trout fed probiotics Kocuria SM1 and Rhodococcus SM2. Aquaculture 2014, 418, 55–61. [Google Scholar] [CrossRef]
- Bezkorovainy, A. Probiotics: Determinants of survival and growth in the gut. Am. J. Clin. Nutr. 2001, 73, 399s–405s. [Google Scholar] [CrossRef]
- Czajkowski, R.; Jafra, S. Quenching of acyl-homoserine lactone-dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochim. Pol. 2009, 56, 1–16. [Google Scholar] [CrossRef]
- Ma, Z.-P.; Song, Y.; Cai, Z.-H.; Lin, Z.-J.; Lin, G.-H.; Wang, Y.; Zhou, J. Anti-quorum sensing activities of selected coral symbiotic bacterial extracts from the South China Sea. Front. Cell. Infect. Microbiol. 2018, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. Probiotics in marine larviculture. FEMS Microbiol. Rev. 2006, 30, 404–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Xia, Y.; Zhu, C.; Chu, W. Isolation of marine Bacillus sp. with antagonistic and organic-substances-degrading activities and its potential application as a fish probiotic. Mar. Drugs 2018, 16, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morohoshi, T.; Ebata, A.; Nakazawa, S.; Kato, N.; Ikeda, T. N-acyl homoserine lactone-producing or-degrading bacteria isolated from the intestinal microbial flora of ayu fish (Plecoglossus altivelis). Microbes Environ. 2005, 20, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.; Yepes, A.; Garcia-Betancur, J.C.; Westedt, I.; Mielich, B.; López, D. Streptomycin-induced expression in Bacillus subtilis of YtnP, a lactonase-homologous protein that inhibits development and streptomycin production in Streptomyces griseus. Appl. Environ. Microbiol. 2012, 78, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Barbeyron, T.; Brillet-Guéguen, L.; Carré, W.; Carrière, C.; Caron, C.; Czjzek, M.; Hoebeke, M.; Michel, G. Matching the diversity of sulfated biomolecules: Creation of a classification database for sulfatases reflecting their substrate specificity. PLoS ONE 2016, 11, e0164846. [Google Scholar] [CrossRef]
- Bebrone, C. Metallo-β-lactamases (classification, activity, genetic organization, structure, zinc coordination) and their superfamily. Biochem. Pharmacol. 2007, 74, 1686–1701. [Google Scholar] [CrossRef]
- Seo, M.-J.; Lee, B.-S.; Pyun, Y.-R.; Park, H. Isolation and characterization of N-acylhomoserine lactonase from the thermophilic bacterium, Geobacillus caldoxylosilyticus YS-8. Biosci. Biotechnol. Biochem. 2011, 1108022598. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.W.; Stone, E.M.; Costello, A.L.; Tierney, D.L.; Fast, W. The quorum-quenching lactonase from Bacillus thuringiensis is a metalloprotein. Biochemistry 2005, 44, 7559–7569. [Google Scholar] [CrossRef]
- See-Too, W.S.; Ee, R.; Lim, Y.-L.; Convey, P.; Pearce, D.A.; Yin, W.-F.; Chan, K.-G. AidP, a novel N-Acyl homoserine lactonase gene from Antarctic Planococcus sp. Sci. Rep. 2017, 7, 42968. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Kang, H.-O.; Jang, H.-S.; Lee, J.-K.; Koo, B.-T.; Yum, D.-Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Fu, K.; Wang, Y.; Wu, C.; Li, F.; Shi, L.; Ge, Y.; Zhou, L. Detection of diverse N-acyl-homoserine lactones in Vibrio alginolyticus and regulation of biofilm formation by N-(3-oxodecanoyl) homoserine lactone in vitro. Front. Microbiol. 2017, 8, 1097. [Google Scholar] [CrossRef] [PubMed]
- Tinh, N.T.N.; Asanka Gunasekara, R.; Boon, N.; Dierckens, K.; Sorgeloos, P.; Bossier, P. N-acyl homoserine lactone-degrading microbial enrichment cultures isolated from Penaeus vannamei shrimp gut and their probiotic properties in Brachionus plicatilis cultures. FEMS Microbial. Ecol. 2007, 62, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, C.R.; Almeida, E.M.; Guerreiro, I.; Santos, R.; Merrifield, D.L.; Tavares, F.; Oliva-Teles, A.; Enes, P. Selection of carbohydrate-active probiotics from the gut of carnivorous fish fed plant-based diets. Sci. Rep. 2019, 9, 6384. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Ray, A.; Ghosh, K.; Ringø, E. Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.; Askarian, F.; Ringø, E. Characterisation and identification of enzyme-producing autochthonous bacteria from the gastrointestinal tract of two Indian air-breathing fish. Benef. Microbes 2013, 4, 277–284. [Google Scholar] [CrossRef]
- Ghosh, K.; Sen, S.K.; Ray, A.K. Characterization of Bacilli isolated from the gut of rohu, Labeo rohita, fingerlings and its significance in digestion. J. Appl. Aquac. 2002, 12, 33–42. [Google Scholar] [CrossRef]
- Bairagi, A.; Ghosh, K.S.; Sen, S.K.; Ray, A.K. Enzyme producing bacterial flora isolated from fish digestive tracts. Aquac. Int. 2002, 10, 109–121. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbial. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Mohammadian, T.; Alishahi, M.; Tabandeh, M.R.; Nejad, A.J.; Karami, E.; Zarea, M. Effects of autochthonous probiotics, isolated from Tor grypus (Karaman, 1971) intestine and Lactobacillus casei (PTCC 1608) on expression of immune-related genes. Aquac. Int. 2019, 27, 239–260. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Xiao, P.; Li, G.; Yue, S.; Huang, J.; Zhu, W.; Mo, Z. Isolation and characterization of Bacillus spp. M 001 for potential application in turbot (Scophthalmus maximus L.) against Vibrio anguillarum. Aquac. Nutr. 2016, 22, 374–381. [Google Scholar] [CrossRef]
- Kavitha, M.; Raja, M.; Perumal, P. Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822). Aquac. Rep. 2018, 11, 59–69. [Google Scholar] [CrossRef]
- Sansawat, A.; Thirabunyanon, M. Anti-Aeromonas hydrophila activity and characterisation of novel probiotic strains of Bacillus subtilis isolated from the gastrointestinal tract of giant freshwater prawns. Maejo Int. J. Sci. Technol. 2009, 3, 77–87. [Google Scholar]
- Sorroza, L.; Padilla, D.; Acosta, F.; Román, L.; Grasso, V.; Vega, J.; Real, F. Characterization of the probiotic strain Vagococcus fluvialis in the protection of European sea bass (Dicentrarchus labrax) against vibriosis by Vibrio anguillarum. Vet. Microbiol. 2012, 155, 369–373. [Google Scholar] [CrossRef]
- Sánchez-Ortiz, A.C.; Luna-González, A.; Campa-Córdova, Á.I.; Escamilla-Montes, R.; del Carmen Flores-Miranda, M.; Mazón-Suástegu, J.M. Isolation and characterization of potential probiotic bacteria from pustulose ark (Anadara tuberculosa) suitable for shrimp farming. Lat. Am. J. Aquat. Res. 2015, 43, 123–136. [Google Scholar] [CrossRef]
- Meidong, R.; Doolgindachbaporn, S.; Jamjan, W.; Sakai, K.; Tashiro, Y.; Okugawa, Y.; Tongpim, S. A novel probiotic Bacillus siamensis B44v isolated from Thai pickled vegetables (Phak-dong) for potential use as a feed supplement in aquaculture. J. Gen. Appl. Microbiol. 2017, 63, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Mizan, M.F.R.; Jahid, I.K.; Kim, M.; Lee, K.-H.; Kim, T.J.; Ha, S.-D. Variability in biofilm formation correlates with hydrophobicity and quorum sensing among Vibrio parahaemolyticus isolates from food contact surfaces and the distribution of the genes involved in biofilm formation. Biofouling 2016, 32, 497–509. [Google Scholar] [CrossRef]
- Wang, L.-Q.; Meng, X.-C.; Zhang, B.-R.; Wang, Y.; Shang, Y.-L. Influence of cell surface properties on adhesion ability of bifidobacteria. World J. Microbiol. Biotechnol. 2010, 26, 1999–2007. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Janković, T.; Frece, J.; Abram, M.; Gobin, I. Aggregation ability of potential probiotic Lactobacillus plantarum strains. Int. J. Sanit. En.g Res. 2012, 6, 19–24. [Google Scholar]
- Geraylou, Z.; Vanhove, M.P.; Souffreau, C.; Rurangwa, E.; Buyse, J.; Ollevier, F. In vitro selection and characterization of putative probiotics isolated from the gut of Acipenser baerii (Brandt, 1869). Aquac. Res. 2014, 45, 341–352. [Google Scholar] [CrossRef]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. In vitro growth characteristics of five candidate aquaculture probiotics and two fish pathogens grown in fish intestinal mucus. FEMS Microbiol. Lett. 2004, 231, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Kusada, H.; Zhang, Y.; Tamaki, H.; Kimura, N.; Kamagata, Y. Novel N-acyl homoserine lactone-degrading bacteria isolated from penicillin-contaminated environments and their quorum-quenching activities. Front. Microbiol. 2019, 10, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, D.; Chen, B.; Bai, R.; Song, P.; Lin, H. Contamination of sulfonamide antibiotics and sulfamethazine-resistant bacteria in the downstream and estuarine areas of Jiulong River in Southeast China. Environ. Sci. Pollut. Res. Int. 2015, 22, 12104–12113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steindler, L.; Venturi, V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Defoirdt, T.; Verstraete, W.; Bossier, P. Luminescence, virulence and quorum sensing signal production by pathogenic Vibrio campbellii and Vibrio harveyi isolates. J. Appl. Microbiol. 2008, 104, 1480–1487. [Google Scholar] [CrossRef] [Green Version]
- Milton, D.L. Quorum sensing in vibrios: Complexity for diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef]
- Bjelland, A.M.; Sørum, H.; Tegegne, D.A.; Winther-Larsen, H.C.; Willassen, N.P.; Hansen, H. LitR of Vibrio salmonicida is a salinity-sensitive quorum-sensing regulator of phenotypes involved in host interactions and virulence. Infect. Immun. 2012, 80, 1681–1689. [Google Scholar] [CrossRef] [Green Version]
- Olsson, J.C.; Westerdahl, A.; Conway, P.L.; Kjelleberg, S. Intestinal colonization potential of turbot (Scophthalmus maximus)-and dab (Limanda limanda)-associated bacteria with inhibitory effects against Vibrio anguillarum. Appl. Environ. Microbiol. 1992, 58, 551–556. [Google Scholar]
- Lee, S.J.; Park, S.-Y.; Lee, J.-J.; Yum, D.-Y.; Koo, B.-T.; Lee, J.-K. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl. Environ. Microbiol. 2002, 68, 3919–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greisen, K.; Loeffelholz, M.; Purohit, A.; Leong, D. PCR primers and probes for the 16S rRNA gene of most species of pathogenic bacteria, including bacteria found in cerebrospinal fluid. J. Clin. Microbiol. 1994, 32, 335–351. [Google Scholar] [PubMed]
- Iwamoto, T.; Tani, K.; Nakamura, K.; Suzuki, Y.; Kitagawa, M.; Eguchi, M.; Nasu, M. Monitoring impact of in situ biostimulation treatment on groundwater bacterial community by DGGE. FEMS Microbiol. Ecol. 2000, 32, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Butcher, B.G.; Helmann, J.D. Identification of Bacillus subtilis σW-dependent genes that provide intrinsic resistance to antimicrobial compounds produced by Bacilli. Mol. Microbiol. 2006, 60, 765–782. [Google Scholar] [CrossRef]
- Nakayama, T.; Lu, H.; Nomura, N. Inhibitory effects of Bacillus probionts on growth and toxin production of Vibrio harveyi pathogens of shrimp. Lett. Appl. Microbiol. 2009, 49, 679–684. [Google Scholar] [CrossRef]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan, J.E.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudi, E.; Ahmadi, A.; Sayed-Tabatabaei, B.; Ghobadi, C.; Akhavan, A.; Hasanzadeh, N.; Venturi, V. A novel AHL-degrading rhizobacterium quenches the virulence of Pectobacterium atrosepticum on potato plant. J. Plant Pathol. 2011, 93, 587–594. [Google Scholar]
- Ramesh, D.; Vinothkanna, A.; Rai, A.K.; Vignesh, V.S. Isolation of potential probiotic Bacillus spp. and assessment of their subcellular components to induce immune responses in Labeo rohita against Aeromonas hydrophila. Fish Shellfish. Immunol. 2015, 45, 268–276. [Google Scholar] [CrossRef]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Reda, R.M.; Selim, K.M.; El-Sayed, H.M.; El-Hady, M. In Vitro Selection and Identification of Potential Probiotics Isolated from the Gastrointestinal Tract of Nile Tilapia, Oreochromis niloticus. Probiotics Antimicrob. Proteins 2018, 10, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Arpana, M.; Gulab, S.; Varsha, G.; Anita, Y.; Aneja, K.R.; Gautam, S.K.; Aggarwal, N.K. Isolation and biochemical characterization of acido-thermophilic extracellular phytase producing bacterial strain for potential application in poultry feed. Adv. Appl. Res. 2011, 4, 26–34. [Google Scholar]
- Ran, C.; Carrias, A.; Williams, M.A.; Capps, N.; Dan, B.C.; Newton, J.C.; Kloepper, J.W.; Ooi, E.L.; Browdy, C.L.; Terhune, J.S. Identification of Bacillus strains for biological control of catfish pathogens. PLoS ONE 2012, 7, e45793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifuzzaman, S.; Rahman, H.; Austin, D.A.; Austin, B. Properties of probiotics Kocuria SM1 and Rhodococcus SM2 isolated from fish guts. Probiotics Antimicrob. Proteins 2018, 10, 534–542. [Google Scholar] [CrossRef]
- Muñoz-Atienza, E.; Araújo, C.; Magadán, S.; Hernández, P.E.; Herranz, C.; Santos, Y.; Cintas, L.M. In vitro and in vivo evaluation of lactic acid bacteria of aquatic origin as probiotics for turbot (Scophthalmus maximus L.) farming. Fish Shellfish. Immunol. 2014, 41, 570–580. [Google Scholar] [CrossRef]
- Handley, P.S.; Harty, D.W.; Wyatt, J.E.; Brown, C.R.; Doran, J.P.; Gibbs, A.C. A comparison of the adhesion, coaggregation and cell-surface hydrophobicity properties of fibrillar and fimbriate strains of Streptococcus salivarius. Microbiology 1987, 133, 3207–3217. [Google Scholar] [CrossRef] [Green Version]
- Mohammadian, T.; Alishahi, M.; Tabandeh, M.R.; Ghorbanpoor, M.; Gharibi, D.; Tollabi, M.; Rohanizade, S. Probiotic effects of Lactobacillus plantarum and L. delbrueckii ssp. bulguricus on some immune-related parameters in Barbus grypus. Aquac. Int. 2016, 24, 225–242. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. Available online: https://www.ncbi.nlm.nih.gov/books/NBK54050 (accessed on 25 December 2019).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Number | Closest Known Species (16S rDNA Sequence) | Identity (%) | Location-Culture System | Accession Number |
|---|---|---|---|---|
| QQ1 | Bacillus thuringiensis | 99.76 | Bushehr-cage culture | SAMN13108109 |
| QQ2 | Bacillus cereus | 99.72 | Hormozgan-cage culture | SAMN13108110 |
| QQ3 | Bacillus thuringiensis | 100 | Hormozgan-cage culture | SAMN13108111 |
| QQ4 | Bacillus sp. | 99.29 | Bushehr-cage culture | SAMN13108112 |
| QQ5 | Shewanella algae | 99.08 | Khuzestan-earthen pond | SAMN13108113 |
| QQ6 | Carnobacterium maltaromaticum | 97.41 | Bushehr-cage culture | SAMN13108114 |
| QQ7 | Bacillus sp. | 96.86 | Bushehr-cage culture | SAMN13108115 |
| QQ8 | Bacillus sp. | 96.63 | Bushehr-cage culture | SAMN13108116 |
| QQ9 | Bacillus sp. | 95.35 | Hormozgan-cage culture | SAMN13108117 |
| QQ10 | Bacillus sp. | 96.35 | Hormozgan-cage culture | SAMN13108118 |
| Isolates | Enzyme Production | Spore Formation | Total Score | |||
|---|---|---|---|---|---|---|
| Protease | Lipase | Amylase | Phytase | |||
| QQ1 | ++ | ++ | ++ | − | + | 7 |
| QQ2 | +++ | + | - | − | + | 5 |
| QQ3 | + | − | ++ | + | + | 5 |
| QQ4 | +++ | + | − | + | + | 6 |
| QQ5 | +++ | ++ | − | − | − | 5 |
| Signals (AHLs) | Pathogens | AHL Detector | References |
|---|---|---|---|
| 3-oh-C4-HSL | Vibrio harveyi | A. tumefaciens NTL4 | [68] |
| Vibrio campbelli | |||
| Vibrio parahaemolyticus | |||
| 3-oh-C4-HSL | Vibrio alginolyticus | A. tumefaciens NTL4 | [42] |
| 3-oxo-C10-HSL | A. tumefaciens NTL4 | ||
| 3-oxo-C14-HSL | A. tumefaciens NTL4 | ||
| C6-HSL | Vibrio anguillarum | C. violaceum CV026 | [69] |
| 3-oxo-C10-HSL | A. tumefaciens NTL4 | ||
| C6-HSL | Vibrio salmonicida | C. violaceum CV026 | [70] |
| 3-oxo-C6-HSL | C. violaceum CV026 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghanei-Motlagh, R.; Mohammadian, T.; Gharibi, D.; Menanteau-Ledouble, S.; Mahmoudi, E.; Khosravi, M.; Zarea, M.; El-Matbouli, M. Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer. Mar. Drugs 2020, 18, 23. https://doi.org/10.3390/md18010023
Ghanei-Motlagh R, Mohammadian T, Gharibi D, Menanteau-Ledouble S, Mahmoudi E, Khosravi M, Zarea M, El-Matbouli M. Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer. Marine Drugs. 2020; 18(1):23. https://doi.org/10.3390/md18010023
Chicago/Turabian StyleGhanei-Motlagh, Reza, Takavar Mohammadian, Darioush Gharibi, Simon Menanteau-Ledouble, Esmaeil Mahmoudi, Mohammad Khosravi, Mojtaba Zarea, and Mansour El-Matbouli. 2020. "Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer" Marine Drugs 18, no. 1: 23. https://doi.org/10.3390/md18010023
APA StyleGhanei-Motlagh, R., Mohammadian, T., Gharibi, D., Menanteau-Ledouble, S., Mahmoudi, E., Khosravi, M., Zarea, M., & El-Matbouli, M. (2020). Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer. Marine Drugs, 18(1), 23. https://doi.org/10.3390/md18010023