Quantitative and Qualitative HPLC Analysis of Mycosporine-Like Amino Acids Extracted in Distilled Water for Cosmetical Uses in Four Rhodophyta


Abstract
:1. Introduction
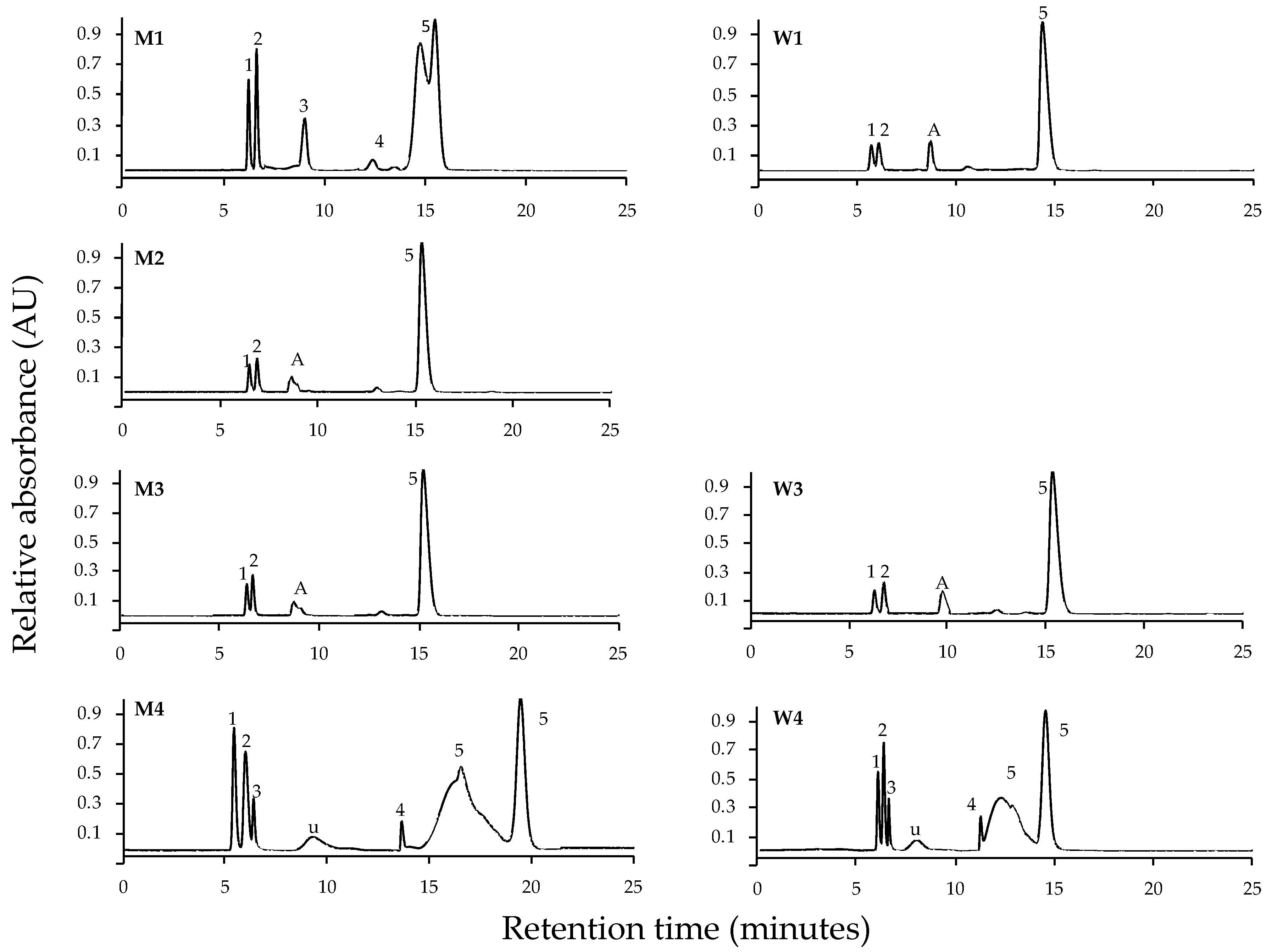
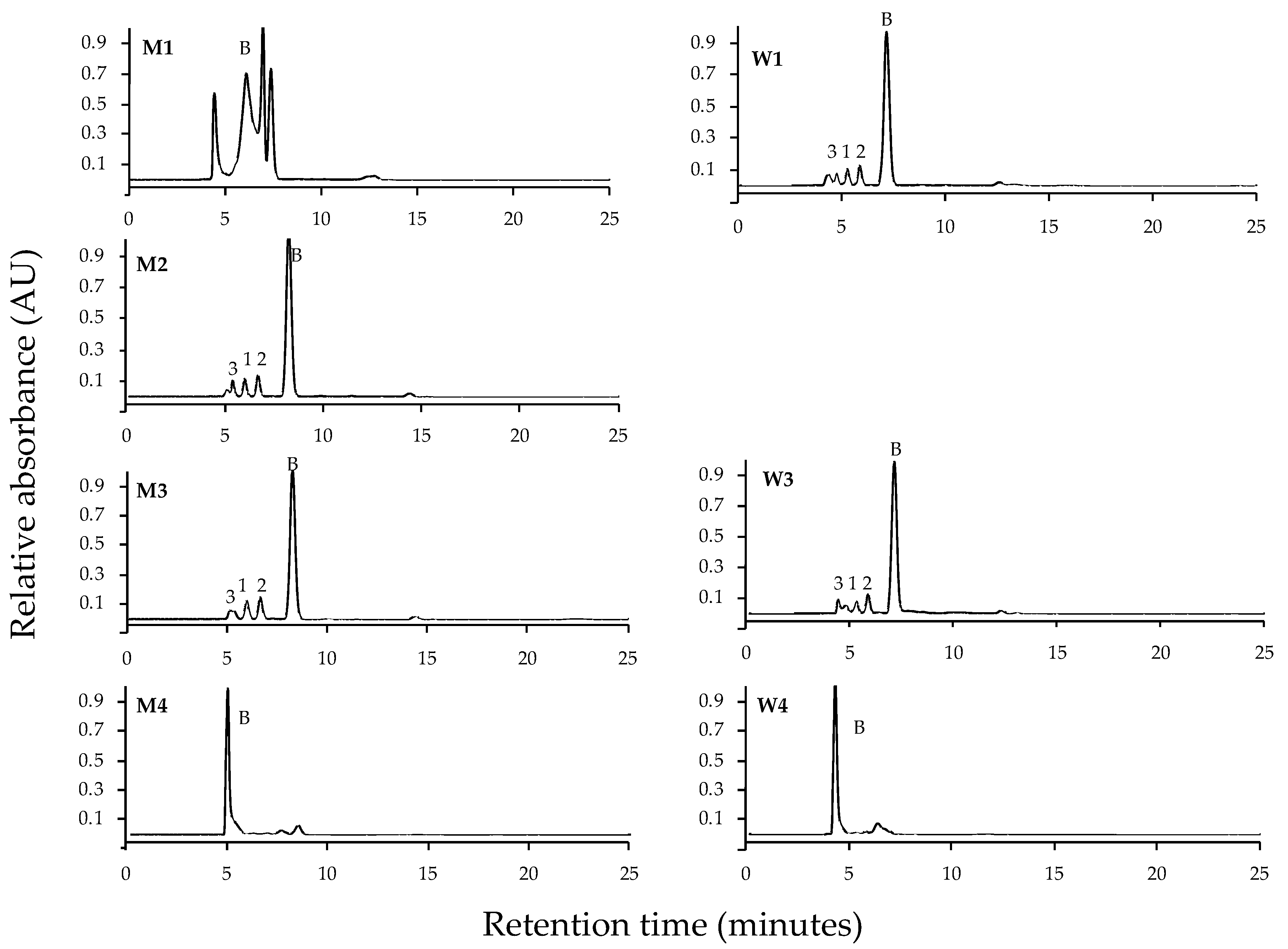
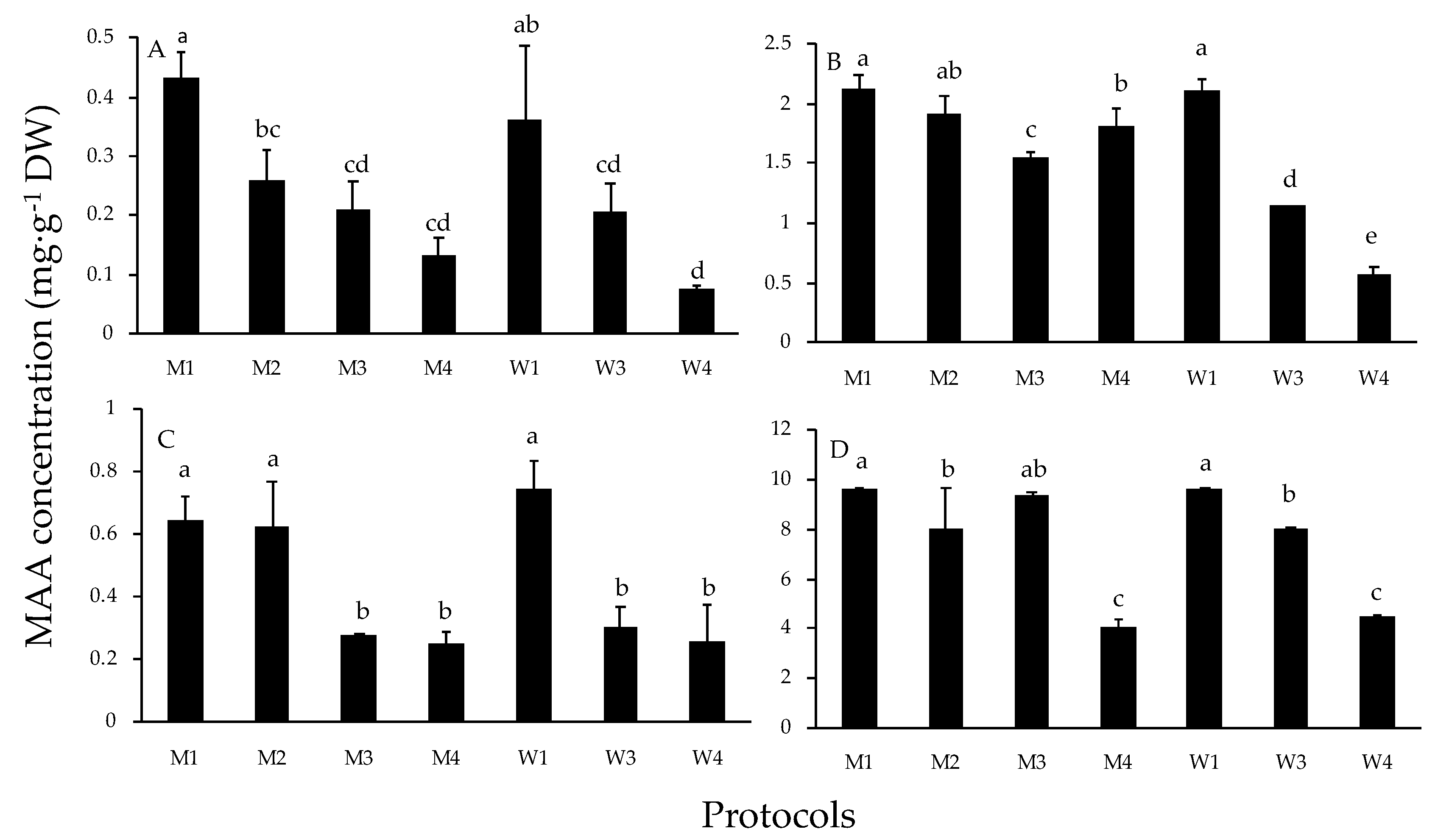
2. Results
3. Discussion and Conclusions
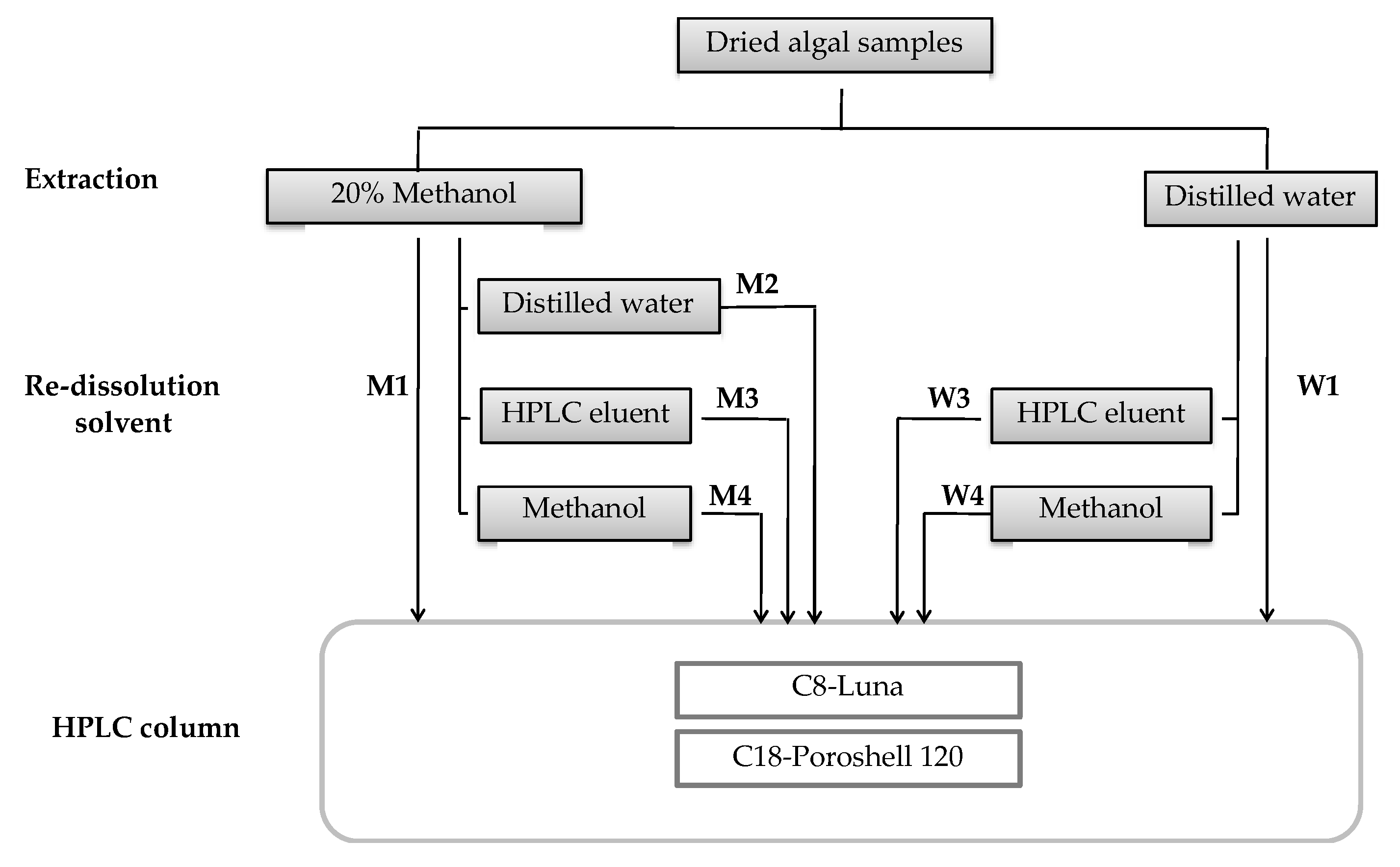
4. Materials and Methods
4.1. Biological Material
4.2. Extraction and Identification of MAAs
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Franklin, L.A.; Forster, R.M. The changing irradiance environment: Consequences for marine macrophyte physiology, productivity and ecology. Eur. J. Phycol. 1997, 32, 207–232. [Google Scholar] [CrossRef] [Green Version]
- Misonou, T.; Saitoh, J.; Oshiba, S.; Tokitomo, Y.; Maegawa, M.; Inoue, Y.; Hori, H.; Sakurai, T. UV-absorbing substance in the red alga Porphyra Yezoensis (Bangiales, Rhodophyta) block thymine photodimer production. Mar. Biotechnol. 2003, 5, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537–558. [Google Scholar] [CrossRef] [PubMed]
- Buma, A.G.J.; Engelen, A.H.; Gieskes, W.W.C. Wavelength-dependent induction of thymine dimers and growth rate reduction in the marine diatom Cyclotellasp exposed to ultraviolet radiation. Mar. Ecol. Prog. Ser. 1997, 153, 91–97. [Google Scholar] [CrossRef]
- Helbling, E.W.; Villafañe, V.E.; Buma, A.G.; Andrade, M.; Zaratti, F. DNA damage and photosynthetic inhibition induced by solar UVR in tropical phytoplankton. Eur. J. Phycol. 2001, 36, 157–166. [Google Scholar] [CrossRef]
- Vicent, W.F.; Roy, S. Solar ultraviolet-B radiation and aquatic primary production: Damage, protection, and recovery. Environ. Rev. 1993, 1, 1–12. [Google Scholar] [CrossRef]
- Häder, D.P.; Lebert, M.; Sinha, R.O.; Barbieri, E.; Herbling, E.W. Role of protective and repair mechanisms in the inhibition of photosynthesis in marine macroalgae. Photochem. Photobiol. Sci. 2002, 1, 809–814. [Google Scholar] [CrossRef]
- Karentz, D. Chemical defenses of marine organisms against solar radiation exposure: UV-absorbing mycosporine-like amino acids and scytonemin. In Marine Chemical Ecology; McClinctock, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 481–520. [Google Scholar]
- Cockell, C.S.; Knowland, J. Ultraviolet radiation screening compounds. Biol. Rev. 1999, 74, 311–345. [Google Scholar] [CrossRef]
- Yoshiki, M.; Tsuge, K.; Tsuruta, Y.; Yoshimura, T.; Koganemaru, K.; Sumi, T.; Matsui, T.; Matsumoto, K. Production of new antioxidant compound from mycosporine-like amino acid, porphyra-334 by heat treatment. Food Chem. 2009, 113, 1127–1132. [Google Scholar] [CrossRef]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L.; De Gálvez, M.V.; Herrera, E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J. Appl. Phycol. 2009, 21, 161–169. [Google Scholar] [CrossRef]
- Karsten, U.; Franklin, L.A.; Lüning, K.; Wiencke, C. Natural ultraviolet and photosynthetic active radiation induce formation of mycosporine-like amino acids in the marine macroalga Chondrus crispus (Rhodophyta). Planta 1998, 205, 257–262. [Google Scholar] [CrossRef]
- Hoyer, K.; Karsten, U.; Wiencke, C. Induction of sunscreen compounds in Antarctic macroalgae by different radiation conditions. Mar. Biol. 2002, 141, 619–627. [Google Scholar]
- Lalegerie, F.; Lajili, S.; Bedoux, G.; Taupin, L.; Stiger-Pouvreau, V.; Connan, S. Photo-protective compounds in red macroalgae from Brittany: Considerable diversity in mycosporine-like amino acids (MAAs). Mar. Enviorn. Res. 2019, 147, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, H.; Kobayashi, J.; Hirata, Y. Separation of mycosporine-like amino acids in marine organisms using reverse-phase high-performance liquid chromatography. J. Chromatogr. 1982, 250, 113–118. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids a related Gadusols: Biosynthesis, accumulation and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [Green Version]
- Korbee-Peinado, N.; Abdala Díaz, R.T.; Figueroa, F.L.; Helbling, E.W. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J. Phycol. 2004, 40, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like amino acids (MAAs): Chemical structure, biosynthesis and significance as UV absorbing/screening compounds. Indian J. Exp. Biol. 2008, 146, 7–17. [Google Scholar]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The photoprotector mechanism of mycosporine-like amino acids. Excited-state properties and photostability of porphyra-334 in aqueous solution. J. Photochem. Photobiol. B Biol. 2000, 56, 139–144. [Google Scholar] [CrossRef]
- De la Coba, F.; Aguilera, J.; Korbee, N.; De Gálvez, M.V.; Herrera, E.; Álvarez-Gómez, F.; Figueroa, F.L. UVA and UVB photoprotective capabilities of topical formulations containing mycosporine-like amino acids (MAAs) through different biological effective protection factors (BEPFs). Mar. Drugs. 2019, 17, 55. [Google Scholar] [CrossRef] [Green Version]
- Schmid, D.; Schürch, C.; Zülli, F.; Nissen, H.-P.; Prieur, H. Mycosporine-like amino acids: Natural UV-screening compounds from red algae to protect the skin against photoaging. SOFW J. 2003, 129, 38–42. [Google Scholar]
- Ryu, J.; Park, S.-J.; Kim, I.-H.; Choi, Y.; Nam, T.-J. Protective effect of porphyra-334 on UVA-induced photoaging in human skin fibroblasts. Int. J. Mol. Med. 2014, 34, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.-S.; Hwang, J.; Park, M.; Seo, H.H.; Kim, H.-S.; Lee, J.H.; Moh, S.H.; Lee, T.-K. Anti-inflammation activities of mycosporine-like amino acids (MAAs) in response to UV radiation suggest potential anti-skin aging activity. Mar. Drugs. 2014, 12, 5174–5187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morliere, P.; Annie, M.; Isabelle, T. Action spectrum for UV-inducen lipid peroxidation in cultured human skin fibroblast. Free Radic. Biol. Med. 1995, 19, 365–371. [Google Scholar] [CrossRef]
- de la Coba, F.; Aguilera, J.; De Galvez, M.V.; Alvarez, M.; Gallego, E.; Figueroa, F.L.; Herrera, E. Prevention of the ultraviolet effects on clinical and histopathological changes, as well as the heat shock protein-70 expression in mouse skin by topical application of algal UV-absorbing compounds. J. Dermatol. Sci. 2009, 55, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Tartarotti, B.; Sommaruga, R. The effect of different methanol concentrations and temperatures on the extraction of mycosporine-like amino acids (MAAs) in algae and zooplankton. Arch. Hydrobiol. 2002, 154, 691–703. [Google Scholar] [CrossRef]
- Volkmann, M.; Gorbushina, A.A. A broadly applicable method for extraction and characterization of mycosporines and mycosporine-like amino acids of terrestrial, marine and fresh distilled water origin. FEMS Microbiol. Lett. 2006, 255, 286–295. [Google Scholar] [CrossRef]
- Karsten, U.; Escoubeyrou, K.; Charles, F. The effect of re-dissolution solvents and HPLC columns on the analysis of mycosporine-like amino acids in the eulittoral macroalgae Prasiola crispa and Porphyra umbilicalis. Helgol. Mar. Res. 2009, 63, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, W.C.; Chalker, B.E. Identification and quantification of near-UV absorbing compounds (S-320) in a hermatypics cleractinian. Coral Reefs 1986, 5, 155–159. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O.; Montoya, N.G. Comparative studies on mycosporine-like amino acids, paralytic shellfish toxins and pigment profiles of the toxic dinoflagellates Alexandrium tamarense, A-catenella and A-minutum. Mar. Ecol. Prog. Ser. 2001, 223, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Gomez, F.; Korbee, N.; Figueroa, F.L. Analysis of antioxidant capacity and bioactive compounds in marine macroalgal and lichenic extracts using different solvents and evaluation methods. Cienc. Mar. 2016, 42, 271–288. [Google Scholar] [CrossRef] [Green Version]
- Barceló-Villalobos, M.; Figueroa, F.L.; Korbee, N.; Álvarez-Gómez, F.; Abreu, M.H. Production of mycosporine-like amino acids from Gracilaria vermiculophylla (Rhodophyta) cultured through one year in an integrated multi-trophic aquaculture (IMTA) system. Mar. Biotechnol. 2017, 19, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Murauer, A.; Ganzera, M. Quantitative analysis of mycosporine-like amino acids in marine algae by capillary electrophoresis with diode-array detection. J. Pharm. Biomed. Anal. 2017, 138, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.N.; Braun, C.; Kvaskoff, D. Extraction and analysis of mycosporine-like amino acids in marine algae. Methods Mol. Biol. 2015, 1308, 119–129. [Google Scholar] [PubMed]
- Bedoux, G.; Hardouin, K.; Marty, C.; Taupin, L.; Vandanjon, L.; Bourgougnon, N. Chemical characterization and photoprotective activity measurement of extracts from the red macroalga Solieria chordalis. Bot. Mar. 2014, 57, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Whitchcad, K.; Hedges, J.I. Analysis of mycosporine-like amino acids in plankton by liquid chromatography electrospray ionization mass spectrometry. Mar. Chem. 2002, 80, 27–39. [Google Scholar]
- Carreto, J.L.; Carignan, M.O.; Montoya, N.G. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar. Biol. 2005, 146, 237–252. [Google Scholar] [CrossRef]
- Orfanoudaki, M.; Hartmann, A.; Karsten, U.; Ganzera, M. Chemical profiling of mycosporine-like amino acids in twenty-three red algal species. J. Physol. 2019, 55, 393–403. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs. 2011, 9, 387–446. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Nyberg, C.D.; Wulff, A. UVR defense mechanisms in eurytopic and invasive Gracilaria vermiculophylla (Gracilariales, Rhodophyta). Physiol. Plantarum. 2012, 146, 205–216. [Google Scholar] [CrossRef]
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.P. Mycosporine-like amino acids in the marine red algae Gracilaria cornea—Effects of UV and heat. Environ. Exp. Bot. 2000, 43, 33–43. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Korbee, N.; Abdala, R.; Jerez, C.G.; López-de la Torre, M.; Güenaga, L.; Gómez-Pinchetii, J.L. Biofiltration of fishpond effluents and accumulation of N-compounds (phycobiliproteins and mycosporine-like amino acids) versus C-compounds (polysaccharides) in Hydropuntia cornea (Rhodophyta). Mar. Pollut. Bull. 2012, 64, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Gómez, F.; Korbee, N.; Casas-Arrojo, V.; Abdala-Díaz, R.T.; Figueroa, F.L. UV photoprotection, cytotoxicity and inmunology capacity of red algae extracts. Molecules 2019, 24, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Gómez, F.; Korbee, N.; Figueroa, F.L. Effects of UV radiation on photosynthesis, antioxidant capacity and the accumulation of bioactive compounds in Gracilariopsis longissima, Hydropuntia cornea and Halopithys incurve (Rhodophyta). J. Phycol. 2019, 55, 1258–1273. [Google Scholar] [CrossRef]
- Torres, P.B.; Chow, F.; Ferreira, M.J.P.; dos Santos, D.Y.A.C. Mycosporine-like amino acids from Gracilariopsis tenuifrons (Gracilariales, Rhodophyta) and its variation under high light. J. Appl. Phycol. 2016, 28, 2035–2040. [Google Scholar] [CrossRef]
- Diehl, N.; Michalik, D.; Zuccarello, G.C.; Karsten, U. Stress metabolite pattern in the eulittoral red alga Pyropiaplicata (Bangiales) in New Zealand—Mycosporine-like amino acids and heterosides. J. Exp. Mar. Biol. 2019, 510, 23–30. [Google Scholar] [CrossRef]
- De Ramos, B.; da Costa, G.B.; Ramlov, F.; Maraschin, M.; Horta, P.A.; Figueroa, F.L.; Korbee, N.; Bonomi-Barufi, J. Ecophysiological implications of UV radiation in the interspecific interaction of Pyropia acanthopora and Grateloupia turuturu (Rhodophyta). Mar. Environ. Res. 2019, 144, 36–45. [Google Scholar] [CrossRef]
- Ashwini, S.; Manjula, S.; Suresh, B. Comparative HPLC analysis of camptothecin and quercetin contents in ethanolic extracts of Gracilariac corticata (J. Agardh). IJPSR. 2017, 8, 4710–4715. [Google Scholar]
- Jung, S.K.; Lee, K.W.; Kim, H.Y.; Oh, M.H.; Byun, S.; Lim, S.H.; Heo, Y.S.; Kang, N.J.; Bode, A.M.; Dong, Z. Myricetin suppresses UVB-induced wrinkle formation and MMP-9 expression by inhibiting Raf. Biochem. Pharm. 2010, 79, 1455–1461. [Google Scholar] [CrossRef] [Green Version]
- Sim, G.S.; Lee, B.C.; Cho, H.S.; Lee, J.W.; Kim, J.H.; Lee, D.H.; Kim, J.H.; Pyo, H.B.; Moon, D.C.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Durr, P. Plant extracts and natural compounds used against UVB-induced photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanna, B.; Brahmbhatt, H.R.; Mishra, A. Phenolic, flavonoid, and amino acid compositions reveal that selected tropical seaweeds have the potential to be functional food ingredients. J. Food Process. Preserv. 2019, 43, e14266. [Google Scholar] [CrossRef]
- Lin, M.C.; Tsai, M.J.; Wen, K.C. Supercritical fluid extraction of flavonoids from Scutellariae Radix. J. Chromatogr. A 1999, 830, 387–395. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. IFRJ 2010, 17, 205–220. [Google Scholar]
- Navarro, N.P.; Figueroa, F.L.; Korbee, N.; Bonomi, J.; Álvarez Gómez, F.; de la Coba, F. Mycosporine-like amino acids form red algae to develop natural UV sunscreens. In Sunscreens: Source, Formulation, Efficacy and Recommendations; Ragesh, P.R., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2018; pp. 99–129. ISBN 978-1-53613-294-6. [Google Scholar]
- Torres, P.; Pires Santos, J.; Chow, F.; dos Santos, D. A comprehensive review of traditional uses, bioactivity potential, and chemical diversity of the genus Gracilaria (Gracilariales, Rhodophyta). Algal Res. 2019, 37, 288–306. [Google Scholar] [CrossRef]
- Takano, S.; Uemura, D.; Hirata, Y. Isolation and structure of a new amino acid, palythine, from the zoanthid Palythoa tuberculosa. Tetrahedron Lett. 1978, 26, 2299–2300. [Google Scholar] [CrossRef]
- Tsujino, I.; Yabe, K.; Sekekawa, I. Isolation and structure of a new amino acid, shinorine, from the red alga Chondrusyendoi. Bot. Mar. 1980, 23, 65–68. [Google Scholar]
- Pliego-Cortés, H.; Bedoux, G.; Boulho, R.; Taupin, L.; Freile-pelegrín, Y.; Bourgougnon, N.; Robledo, D. Stress tolerance and photoadaptation to solar radiation in Rhodymenia pseudopalmat a (Rhodophyta) through mycosporine-like amino acids, phenolic compounds and pigments in an Integrated Muti-Trophic Aquaculture system. Algal Res. 2019, 41, 101542. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | MAAs | % Relative | Mol. Formula | λmax (nm) | Exact (ppm) | Calculated (m/z [M + H]+) | Observed (m/z [M + H]+) |
|---|---|---|---|---|---|---|---|
| Porphyra-334 | 40.19 ± 1.33 | C14H22N2O8 | 334 | 0.8 | 347.14489 | 347.14365 | |
| Shinorine | 10.63 ± 2.71 | C13H20N2O8 | 334 | 1.4 | 333.12924 | 333.12799 | |
| AV | Palythine | 42.61 ± 3.09 | C10H16N2O5 | 320 | 2.9 | 245.11320 | 245.11247 |
| Asterina-330 | 4.97 ± 0.57 | C12H20N2O6 | 330 | 1.8 | 289.13941 | 289.13840 | |
| Porphyra-334 | 18.04 ± 3.85 | C14H22N2O8 | 334 | 0.8 | 347.14489 | 347.14316 | |
| Shinorine | 49.20 ± 2.80 | C13H20N2O8 | 334 | 1.4 | 333.12924 | 333.12781 | |
| CC | Palythine-serine | 6.12 ± 1.42 | C11H18N2O6 | 320 | 3.8 | 275.12376 | 275. 12271 |
| Asterina-330 | 1.27 ± 0.41 | C12H20N2O6 | 330 | 1.8 | 289.13941 | 289.13840 | |
| Palythinol | 30.28 ± 1.74 | C13H22N2O6 | 332 | 0.9 | 303.15506 | 303.15399 | |
| Porphyra-334 | 5.36 ± 3.18 | C14H22N2O8 | 334 | 0.8 | 347.14489 | 347.14371 | |
| Shinorine | 37.05 ± 9.79 | C13H20N2O8 | 334 | 1.4 | 333.12924 | 333.12805 | |
| GL | Palythine | 1.70 ± 0.20 | C10H16N2O5 | 320 | 1.2 | 245.11320 | 245.11290 |
| Asterina-330 | 1.63 ± 0.45 | C12H20N2O6 | 330 | 1.8 | 289.13941 | 289.13849 | |
| Palythinol | 59.28 ± 9.52 | C13H22N2O6 | 332 | 0.9 | 303.15506 | 303.15421 | |
| Porphyra-334 | 79.83 ± 1.98 | C14H22N2O8 | 334 | 0.8 | 347.14489 | 347.14343 | |
| Shinorine | 3.77 ± 0.99 | C13H20N2O8 | 334 | 1.4 | 333.12924 | 333.12799 | |
| PL | Palythine | 6.26 ± 0.95 | C10H16N2O5 | 320 | 1.2 | 245.11320 | 245.11227 |
| Asterina-330 | 7.69 ± 0.80 | C12H20N2O6 | 330 | 1.8 | 289.13941 | 289.13821 | |
| Palythinol | 2.44 ± 0.16 | C13H22N2O6 | 332 | 0.9 | 303.15506 | 303.15411 |
| Agarophyton vermiculophyllum | |||||||
|---|---|---|---|---|---|---|---|
| MAAs | Protocols | ||||||
| M1 | M2 | M3 | M4 | W1 | W3 | W4 | |
| Porphyra-334 | • | • | • | • | • | • | • |
| Shinorine | • | • | • | • | • | • | • |
| Palythine | • | • | • | • | • | • | • |
| Asterina-330 | • | • | • | • | • | • | • |
| Crassiphycus corneus | |||||||
| M1 | M2 | M3 | M4 | W1 | W3 | W4 | |
| Porphyra-334 | • | • | • | • | • | • | • |
| Shinorine | • | • | |||||
| Palythine-serine | • | • | • | • | • | • | • |
| Asterina-330 | • | • | • | • | • | • | • |
| Palythinol | • | • | |||||
| Gracilariopsis longissima | |||||||
| M1 | M2 | M3 | M4 | W1 | W3 | W4 | |
| Porphyra-334 | • | • | • | • | • | • | • |
| Shinorine | • | • | |||||
| Palythine | • | • | • | • | • | • | • |
| Asterina-330 | • | • | • | • | • | • | • |
| Palythinol | • | • | |||||
| Pyropia leucosticta | |||||||
| M1 | M2 | M3 | M4 | W1 | W3 | W4 | |
| Porphyra-334 | • | • | • | • | • | • | • |
| Shinorine | • | • | • | ||||
| Palythine | • | • | • | • | • | • | • |
| Asterina-330 | • | • | • | • | • | • | • |
| Palythinol | • | • | • | ||||
| Extraction Solvent | Re-dissolution Solvent | Abbreviation | Strengths | Weaknesses |
|---|---|---|---|---|
| 20% Methanol | Direct HPLC analysis without re-dissolution | M1 | High MAA concentration | Accumulation of impurities in the column; Toxic for cosmetic use |
| Distilled water | M2 | High MAA concentration | Some unidentified MAAs; Toxic for cosmetic use | |
| HPLC eluent | M3 | Intermediate MAA concentration | Some unidentified MAAs; Toxic for cosmetic use | |
| 100% Methanol | M4 | Improved MAA separation | Low MAA concentration; Toxic for cosmetic use | |
| Distilled water | Direct HPLC analysis without re-dissolution | W1 | High MAA concentration; Suitable for cosmetic | Accumulation of impurities in the column |
| HPLC eluent | W3 | Intermediate MAA concentration;Suitable for cosmetic | Some unidentified MAAs | |
| 100% Methanol | W4 | Improved MAA separation; Suitable for cosmetic | Low MAA concentration |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves-Peña, P.; de la Coba, F.; Figueroa, F.L.; Korbee, N. Quantitative and Qualitative HPLC Analysis of Mycosporine-Like Amino Acids Extracted in Distilled Water for Cosmetical Uses in Four Rhodophyta. Mar. Drugs 2020, 18, 27. https://doi.org/10.3390/md18010027
Chaves-Peña P, de la Coba F, Figueroa FL, Korbee N. Quantitative and Qualitative HPLC Analysis of Mycosporine-Like Amino Acids Extracted in Distilled Water for Cosmetical Uses in Four Rhodophyta. Marine Drugs. 2020; 18(1):27. https://doi.org/10.3390/md18010027
Chicago/Turabian StyleChaves-Peña, Patricia, Francisca de la Coba, Felix L. Figueroa, and Nathalie Korbee. 2020. "Quantitative and Qualitative HPLC Analysis of Mycosporine-Like Amino Acids Extracted in Distilled Water for Cosmetical Uses in Four Rhodophyta" Marine Drugs 18, no. 1: 27. https://doi.org/10.3390/md18010027
APA StyleChaves-Peña, P., de la Coba, F., Figueroa, F. L., & Korbee, N. (2020). Quantitative and Qualitative HPLC Analysis of Mycosporine-Like Amino Acids Extracted in Distilled Water for Cosmetical Uses in Four Rhodophyta. Marine Drugs, 18(1), 27. https://doi.org/10.3390/md18010027