Protective Action of Ostreococcus Tauri and Phaeodactylum Tricornutum Extracts towards Benzo[a]Pyrene-Induced Cytotoxicity in Endothelial Cells


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Carotenoid and Fatty Acid Contents in P. tricornutum and O. tauri Extracts
2.2. Cytotoxic Effects of B[a]P and Microalgal Extracts on Endothelial HMEC-1 Cells
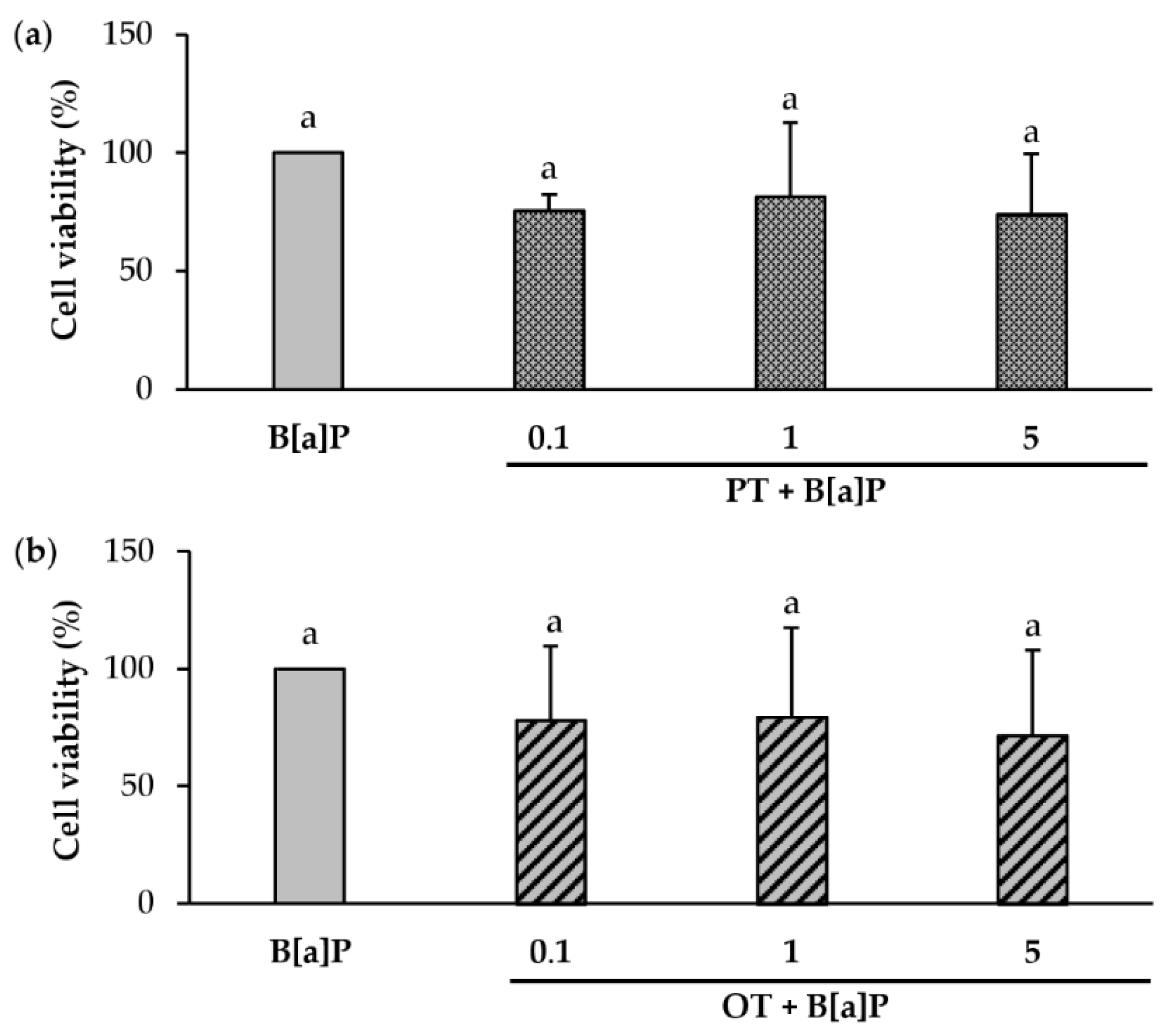
2.3. Effect of Co-Exposure to B[a]P and Extracts of P. tricornutum or O. tauri on the Viability of Endothelial HMEC-1 Cells
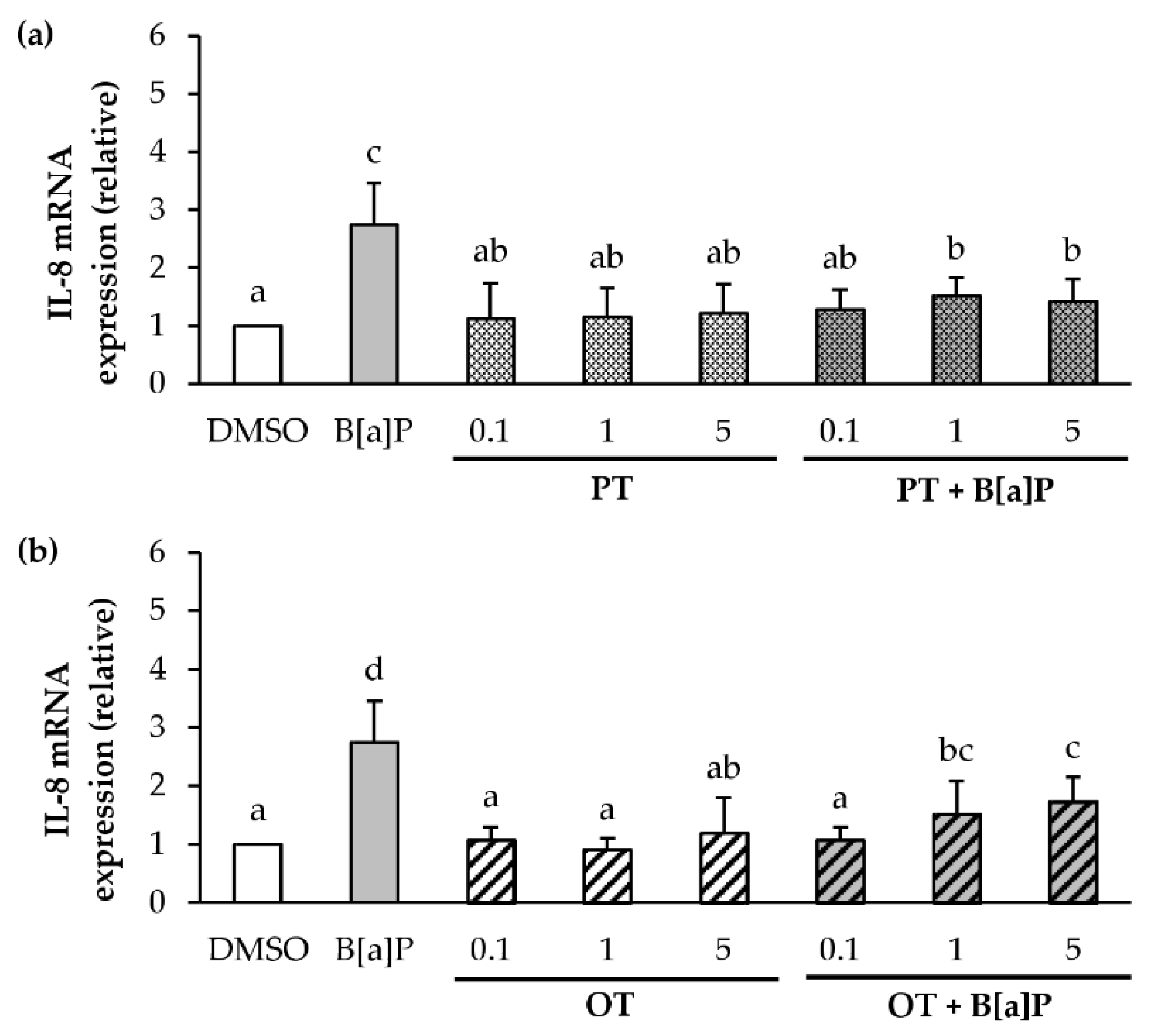
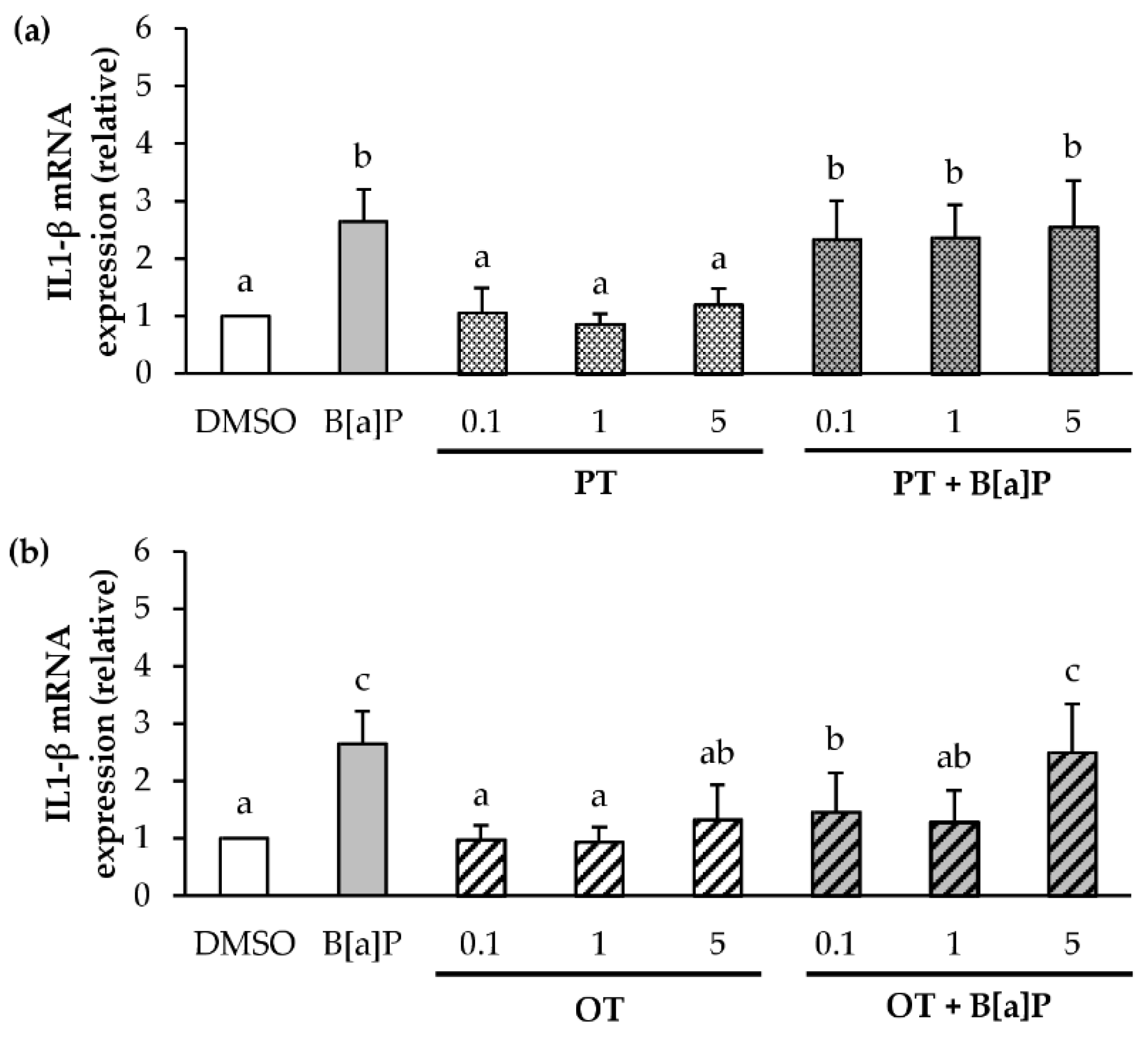
2.4. Effect of P. tricornutum or O. tauri Extracts on Gene Expression of Pro-Inflammatory Cytokines Induced by B[a]P
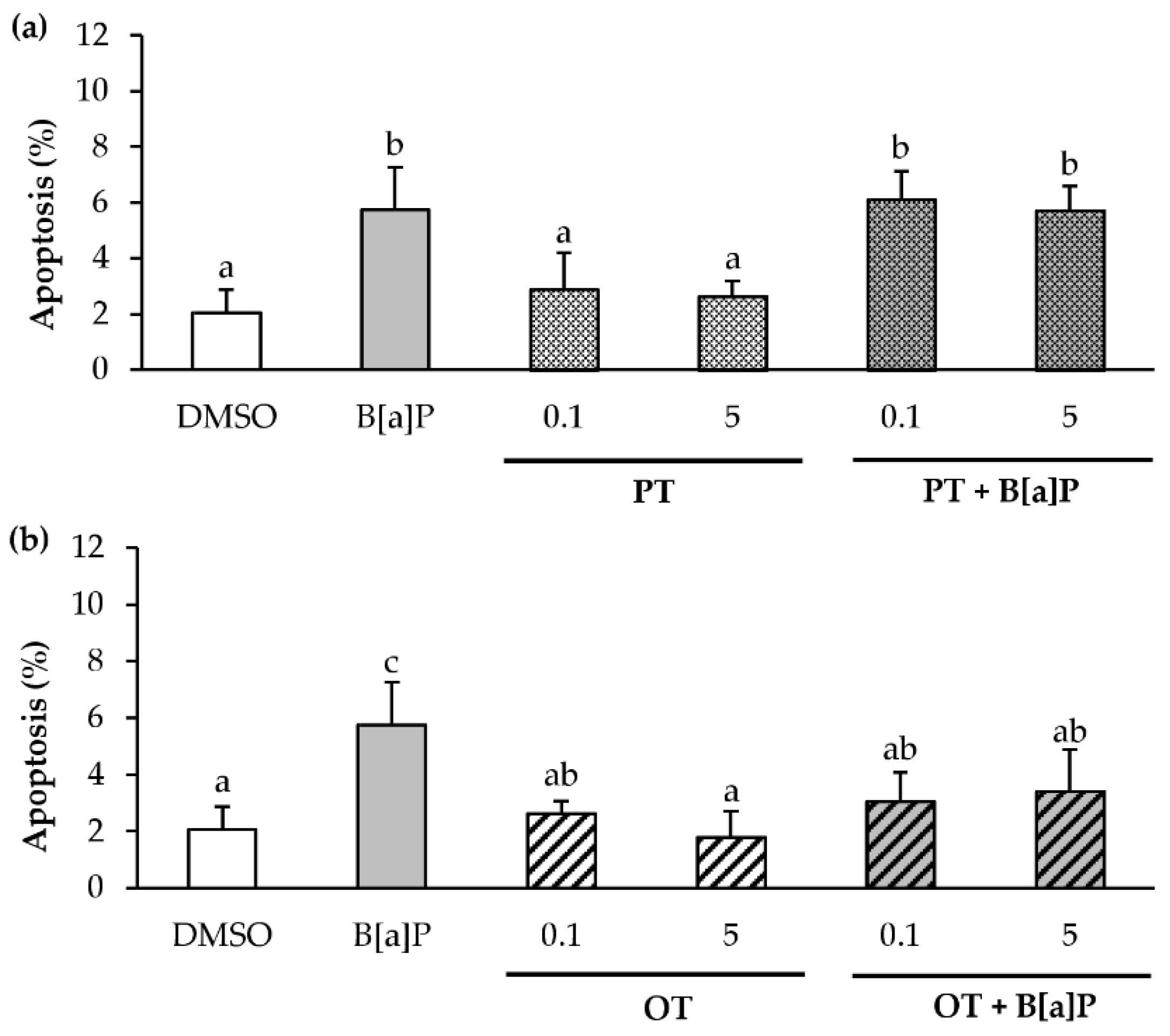
2.5. Only the O. tauri Extract Inhibits the B[a]P-Induced Apoptosis
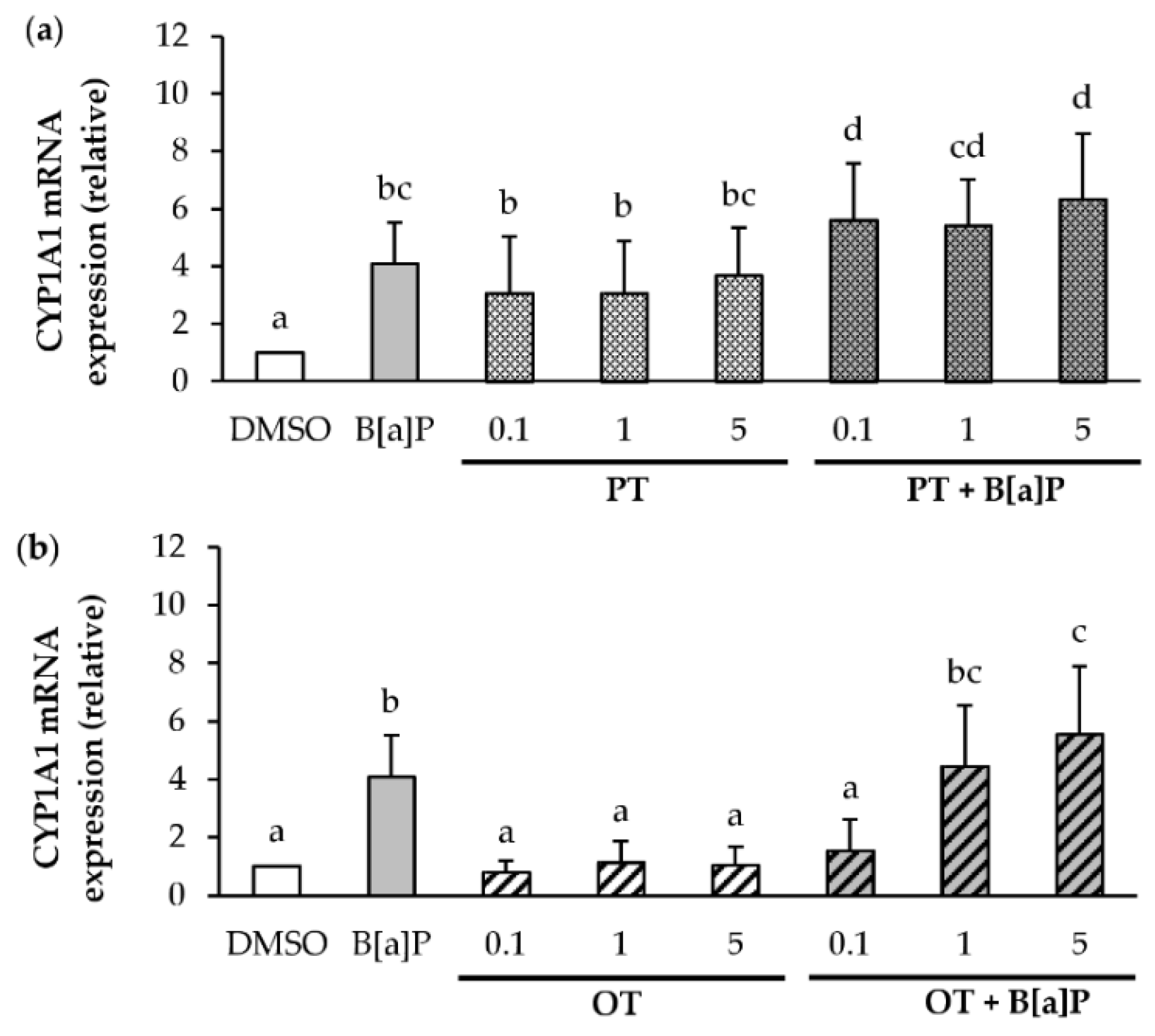
2.6. Effect of P. tricornutum or O. tauri Extracts on the Induction of CYP1A1 by B[a]P
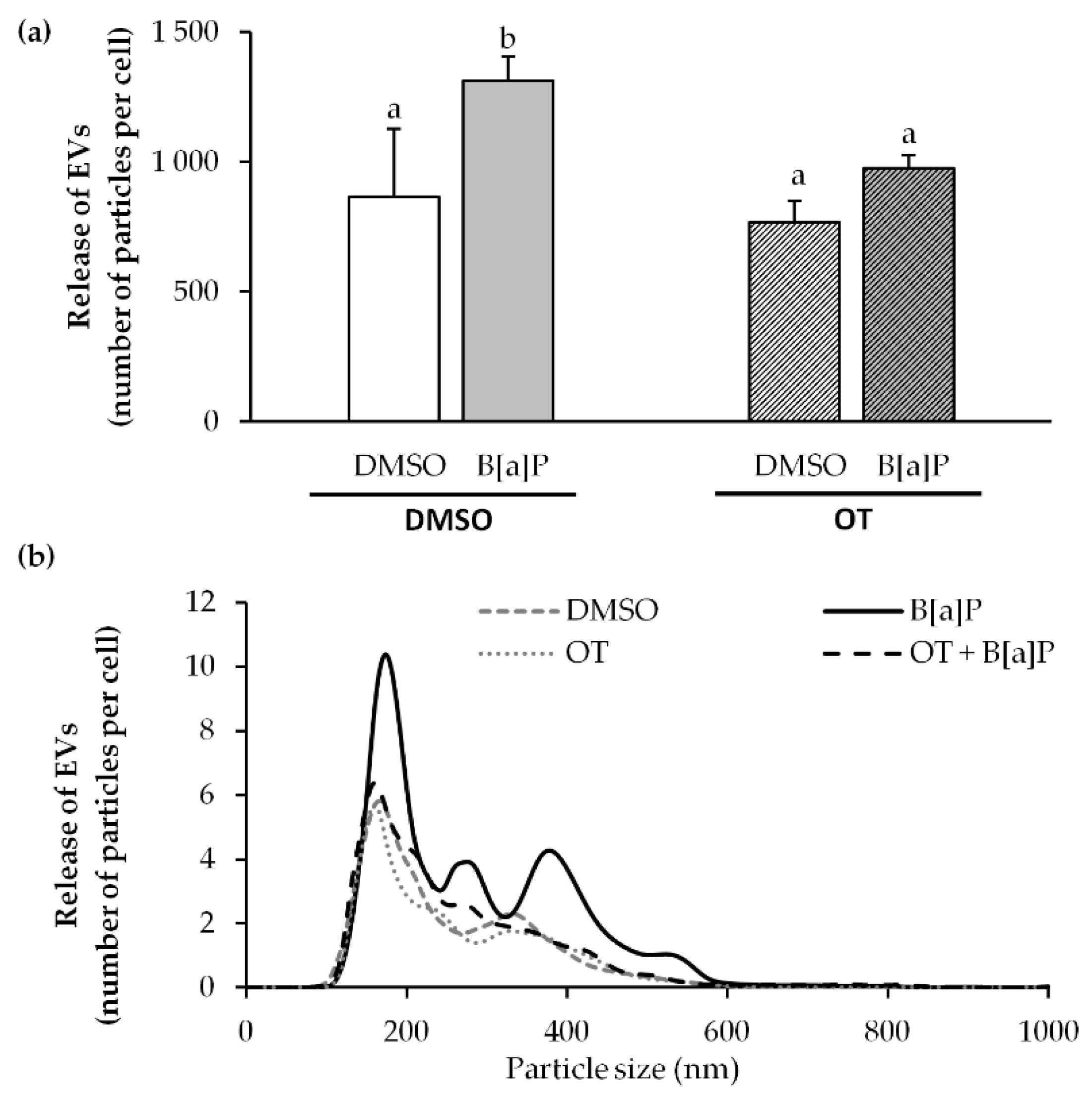
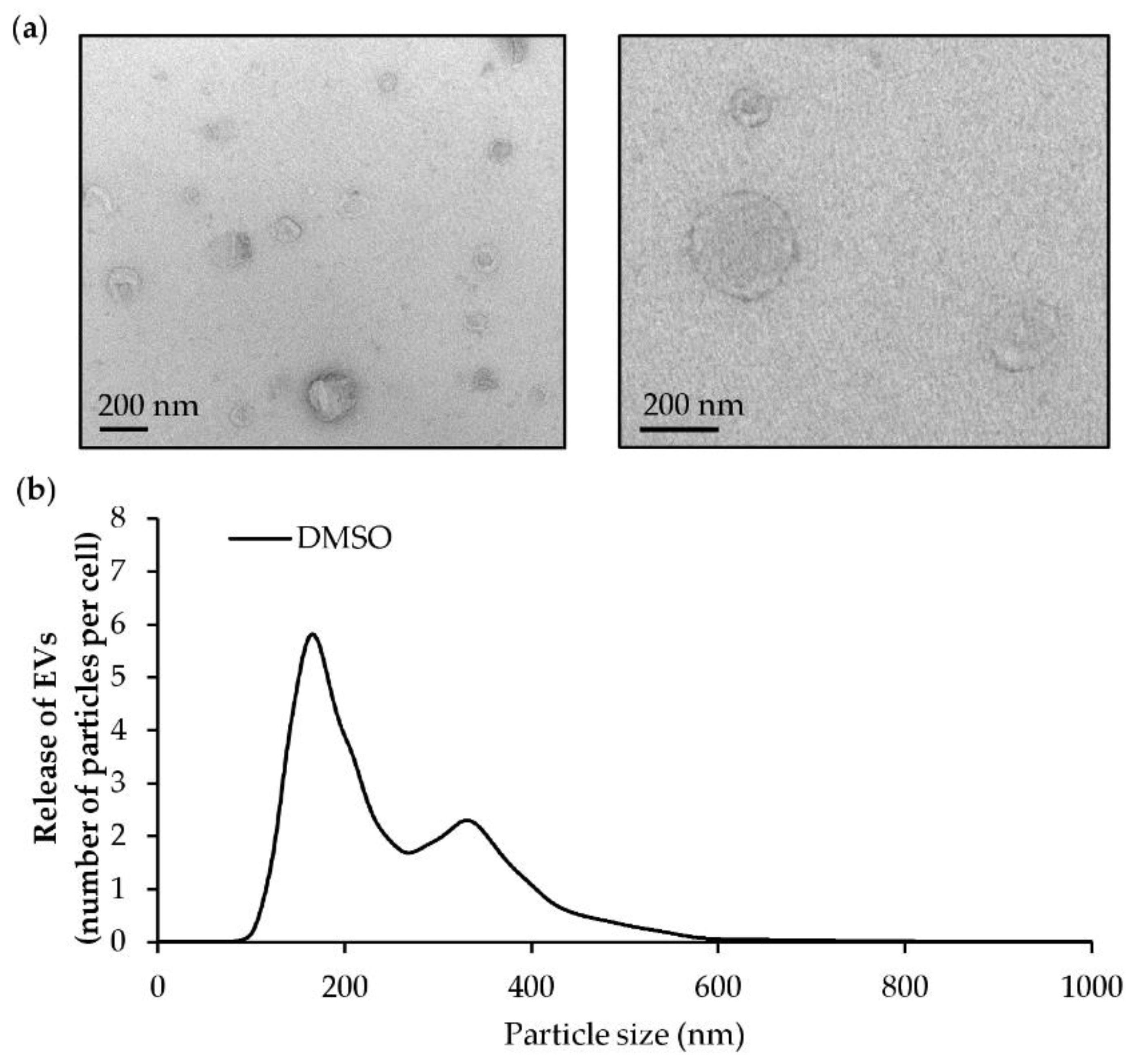
2.7. O. tauri Extract Inhibits the Release of Extracellular Vesicles
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Microalgal Cultivation
4.3. Preparation of Ethanolic Extracts
4.4. Fatty Acid Composition Analyses
4.5. Pigment Composition
4.6. Cell Culture
4.7. Cell Viability Assay
4.8. RNA Isolation and Analysis
4.9. Apoptosis
4.10. Isolation of Extracellular Vesicles
4.11. Nanoparticle Tracking Analysis
4.12. Electron Microscopy
4.13. Characterization of Extracellular Vesicles
4.14. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Lokeshkumar, B.; Rajendran, P.; Nishigaki, I. Exposure to polycyclic aromatic hydrocarbons with special focus on cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Poursafa, P.; Moosazadeh, M.; Abedini, E.; Hajizadeh, Y.; Mansourian, M.; Pourzamani, H.; Amin, M.-M. A systematic review on the effects of polycyclic aromatic hydrocarbons on cardiometabolic impairment. Int. J. Prev. Med. 2017, 8, 19. [Google Scholar]
- Holme, J.A.; Brinchmann, B.C.; Refsnes, M.; Låg, M.; Øvrevik, J. Potential role of polycyclic aromatic hydrocarbons as mediators of cardiovascular effects from combustion particles. Environ. Health 2019, 18, 74. [Google Scholar] [CrossRef] [Green Version]
- Bruzzoniti, M.C.; Fungi, M.; Sarzanini, C. Determination of EPA’s priority pollutant polycyclic aromatic hydrocarbons in drinking waters by solid phase extraction-HPLC. Anal. Methods 2010, 2, 739. [Google Scholar] [CrossRef]
- IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Volume 92, Some non-Heterocyclic Polycyclic Aromatic Hydrocarbons and Some Related Exposures: This Publication Represents the Views and Expert Opinions of an IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, Which Met in Lyon, 11–18 October 2005; International Agency for Research on Cancer (Ed.) WHO: Lyon, France, 2010; ISBN 978-92-832-1292-8. [Google Scholar]
- Das, D.N.; Bhutia, S.K. Inevitable dietary exposure of Benzo[a]pyrene: Carcinogenic risk assessment an emerging issues and concerns. Curr. Opin. Food Sci. 2018, 24, 16–25. [Google Scholar] [CrossRef]
- Hardonnière, K.; Huc, L.; Sergent, O.; Holme, J.A.; Lagadic-Gossmann, D. Environmental carcinogenesis and pH homeostasis: Not only a matter of dysregulated metabolism. Semin. Cancer Biol. 2017, 43, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Yang, C.S. Influences of Dietary and Other Factors on Xenobiotic Metabolism and Carcinogenesis—A Review Article in Memory of Dr. Allan H. Conney (1930–2013). Nutr. Cancer 2015, 67, 1209–1215. [Google Scholar] [CrossRef]
- Mandlekar, S.; Hong, J.-L.; Kong, A.-N.T. Modulation of metabolic enzymes by dietary phytochemicals: A review of mechanisms underlying beneficial versus unfavorable effects. Curr. Drug Metab. 2006, 7, 661–675. [Google Scholar] [CrossRef]
- Azrad, M.; Turgeon, C.; Demark-Wahnefried, W. Current Evidence Linking Polyunsaturated Fatty Acids with Cancer Risk and Progression. Front. Oncol. 2013, 3, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccioni, G.; D’Orazio, N.; Franceschelli, S.; Speranza, L. Marine Carotenoids and Cardiovascular Risk Markers. Mar. Drugs 2011, 9, 1166–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Wu, J.-C.; Lai, C.-S.; Tsai, M.-L.; Ho, C.-T.; Wang, Y.-J.; Pan, M.-H. Chemopreventive effect of natural dietary compounds on xenobiotic-induced toxicity. J. Food Drug Anal. 2017, 25, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tylichová, Z.; Neča, J.; Topinka, J.; Milcová, A.; Hofmanová, J.; Kozubík, A.; Machala, M.; Vondráček, J. n-3 Polyunsaturated fatty acids alter benzo[a]pyrene metabolism and genotoxicity in human colon epithelial cell models. Food Chem. Toxicol. 2019, 124, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Bhagavathy, S.; Sumathi, P. Evaluation of antigenotoxic effects of carotenoids from green algae Chlorococcum humicola using human lymphocytes. Asian Pac. J. Trop. Biomed. 2012, 2, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.B.; Hennig, B. Protective influence of healthful nutrition on mechanisms of environmental pollutant toxicity and disease risks: Nutritional protection and environmental insults. Ann. N. Y. Acad. Sci. 2017, 1398, 99–107. [Google Scholar] [CrossRef]
- Hennig, B.; Petriello, M.C.; Gamble, M.V.; Surh, Y.-J.; Kresty, L.A.; Frank, N.; Rangkadilok, N.; Ruchirawat, M.; Suk, W.A. The role of nutrition in influencing mechanisms involved in environmentally mediated diseases. Rev. Environ. Health 2018, 33, 87–97. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Odjadjare, E.C.; Mutanda, T.; Olaniran, A.O. Potential biotechnological application of microalgae: A critical review. Crit. Rev. Biotechnol. 2017, 37, 37–52. [Google Scholar] [CrossRef]
- Kothari, R.; Pandey, A.; Ahmad, S.; Kumar, A.; Pathak, V.V.; Tyagi, V.V. Microalgal cultivation for value-added products: A critical enviro-economical assessment. 3 Biotech 2017, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.-P.; Morant-Manceau, A.; Schoefs, B. The Potential of Microalgae for the Production of Bioactive Molecules of Pharmaceutical Interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef] [PubMed]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal Derivatives as Potential Nutraceutical and Food Supplements for Human Health: A Focus on Cancer Prevention and Interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef] [Green Version]
- Nazih, H.; Bard, J.-M. Microalgae in Human Health. In Microalgae in Health and Disease Prevention; An Imprint of Elsevier; Academic Press: New York, NY, USA, 2018; pp. 211–226. ISBN 978-0-12-811405-6. [Google Scholar]
- Marin, B.; Melkonian, M. Molecular Phylogeny and Classification of the Mamiellophyceae class. nov. (Chlorophyta) based on Sequence Comparisons of the Nuclear- and Plastid-encoded rRNA Operons. Protist 2010, 161, 304–336. [Google Scholar] [CrossRef]
- Vaezi, R.; Napier, J.; Sayanova, O. Identification and Functional Characterization of Genes Encoding Omega-3 Polyunsaturated Fatty Acid Biosynthetic Activities from Unicellular Microalgae. Mar. Drugs 2013, 11, 5116–5129. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Kirsch, H.; Domergue, F.; Abbadi, A.; Sperling, P.; Bauer, J.; Cirpus, P.; Zank, T.K.; Moreau, H.; Roscoe, T.J.; et al. Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis. J. Lipid Res. 2004, 45, 1899–1909. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.; Hoppe, K.; Czabany, T.; Heilmann, M.; Daum, G.; Feussner, I.; Fulda, M. Identification and characterization of an acyl-CoA:diacylglycerol acyltransferase 2 (DGAT2) gene from the microalga O. tauri. Plant Physiol. Biochem. 2010, 48, 407–416. [Google Scholar] [CrossRef]
- Six, C.; Worden, A.Z.; Rodríguez, F.; Moreau, H.; Partensky, F. New Insights into the Nature and Phylogeny of Prasinophyte Antenna Proteins: Ostreococcus tauri, a Case Study. Mol. Biol. Evol. 2005, 22, 2217–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, F.; Derelle, E.; Guillou, L.; Le Gall, F.; Vaulot, D.; Moreau, H. Ecotype diversity in the marine picoeukaryote Ostreococcus (Chlorophyta, Prasinophyceae). Environ. Microbiol. 2005, 7, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Kotake-Nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids Affect Proliferation of Human Prostate Cancer Cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef] [PubMed]
- Honold, P.J.; Jacobsen, C.; Jónsdóttir, R.; Kristinsson, H.G.; Hermund, D.B. Potential seaweed-based food ingredients to inhibit lipid oxidation in fish-oil-enriched mayonnaise. Eur. Food Res. Technol. 2016, 242, 571–584. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Du, L.; Hosokawa, M.; Miyashita, K.; Kokubun, Y.; Arai, H.; Taroda, H. Fatty Acid and Lipid Class Composition of the Microalga Phaeodactylum tricornutum. J. Oleo Sci. 2017, 66, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.L.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab. Eng. 2014, 22, 3–9. [Google Scholar] [CrossRef]
- Sayanova, O.; Mimouni, V.; Ulmann, L.; Morant-Manceau, A.; Pasquet, V.; Schoefs, B.; Napier, J.A. Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160407. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Jung, Y.-J.; Kwon, O.-N.; Cha, K.H.; Um, B.-H.; Chung, D.; Pan, C.-H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Zarekarizi, A.; Hoffmann, L.; Burritt, D. Approaches for the sustainable production of fucoxanthin, a xanthophyll with potential health benefits. J. Appl. Phycol. 2019, 31, 281–299. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flaiz Flister, V.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S. Fucoxanthin, A Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities In Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Gateau, H.; Solymosi, K.; Marchand, J.; Schoefs, B. Carotenoids of Microalgae Used in Food Industry and Medicine. Mini-Rev. Med. Chem. 2017, 17, 1140–1172. [Google Scholar] [CrossRef] [PubMed]
- Michiels, C. Endothelial cell functions. J. Cell. Physiol. 2003, 196, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Hou, J.; Zhou, Y.; Sun, H.; Yin, W.; Zhang, Y.; Wang, X.; Wang, G.; Chen, W.; Yuan, J. Association of polycyclic aromatic hydrocarbons exposure with atherosclerotic cardiovascular disease risk: A role of mean platelet volume or club cell secretory protein. Environ. Pollut. 2018, 233, 45–53. [Google Scholar] [CrossRef]
- He, X.; Chen, Y.; Zhang, C.; Gong, W.; Zhang, X.; Nie, S. Polycyclic Aromatic Hydrocarbons from Particulate Matter 2.5 (PM2.5) in Polluted Air Changes miRNA Profile Related to Cardiovascular Disease. Med. Sci. Monit. 2018, 24, 5925–5934. [Google Scholar] [CrossRef]
- Franses, J.W.; Drosu, N.C.; Gibson, W.J.; Chitalia, V.C.; Edelman, E.R. Dysfunctional endothelial cells directly stimulate cancer inflammation and metastasis: Dysfunctional endothelium stimulates metastasis. Int. J. Cancer 2013, 133, 1334–1344. [Google Scholar] [CrossRef] [Green Version]
- Ba, Q.; Li, J.; Huang, C.; Qiu, H.; Li, J.; Chu, R.; Zhang, W.; Xie, D.; Wu, Y.; Wang, H. Effects of Benzo[a]pyrene Exposure on Human Hepatocellular Carcinoma Cell Angiogenesis, Metastasis, and NF- κ B Signaling. Environ. Health Perspect. 2015, 123, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W.; Mantoura, R.F.C.; Wright, S.W. (Eds.) Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Monographs on Oceanographic Methodology; UNESCO Pub: Paris, France, 1997; ISBN 978-92-3-103275-2. [Google Scholar]
- Latasa, M.; Scharek, R.; Gall, F.L.; Guillou, L. Pigments suites and taxonomic groups in prasinophyceae. J. Phycol. 2004, 40, 1149–1155. [Google Scholar] [CrossRef]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. (Eds.) Carotenoids Handbook; Birkhäuser Verlag: Basel, Switzerland; Boston, MA, USA, 2004; ISBN 978-3-7643-6180-8. [Google Scholar]
- Ajayi, B.O.; Adedara, I.A.; Farombi, E.O. Benzo(a)pyrene induces oxidative stress, pro-inflammatory cytokines, expression of nuclear factor-kappa B and deregulation of wnt/beta-catenin signaling in colons of BALB/c mice. Food Chem. Toxicol. 2016, 95, 42–51. [Google Scholar] [CrossRef]
- Ji, K.; Xing, C.; Jiang, F.; Wang, X.; Guo, H.; Nan, J.; Qian, L.; Yang, P.; Lin, J.; Li, M.; et al. Benzo[a]pyrene induces oxidative stress and endothelial progenitor cell dysfunction via the activation of the NF-κB pathway. Int. J. Mol. Med. 2013, 31, 922–930. [Google Scholar] [CrossRef]
- Podechard, N.; Lecureur, V.; Le Ferrec, E.; Guenon, I.; Sparfel, L.; Gilot, D.; Gordon, J.R.; Lagente, V.; Fardel, O. Interleukin-8 induction by the environmental contaminant benzo(a)pyrene is aryl hydrocarbon receptor-dependent and leads to lung inflammation. Toxicol. Lett. 2008, 177, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Brinchmann, B.C.; Skuland, T.; Rambøl, M.H.; Szoke, K.; Brinchmann, J.E.; Gutleb, A.C.; Moschini, E.; Kubátová, A.; Kukowski, K.; Le Ferrec, E.; et al. Lipophilic components of diesel exhaust particles induce pro-inflammatory responses in human endothelial cells through AhR dependent pathway(s). Part. Fibre Toxicol. 2018, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, U.; Louis, S.; Gille, A.; Derwenskus, F.; Schmid-Staiger, U.; Briviba, K.; Bischoff, S.C. Anti-inflammatory effects of Phaeodactylum tricornutum extracts on human blood mononuclear cells and murine macrophages. J. Appl. Phycol. 2018, 30, 2837–2846. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta BBA—Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Ren, A.; Wang, L.; Huang, Y.; Wang, Y.; Wang, C.; Greene, N.D. Oxidative Stress and Apoptosis in Benzo[a]pyrene-Induced Neural Tube Defects. Free Radic. Biol. Med. 2018, 116, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Tekpli, X.; Rissel, M.; Huc, L.; Catheline, D.; Sergent, O.; Rioux, V.; Legrand, P.; Holme, J.A.; Dimanche-Boitrel, M.-T.; Lagadic-Gossmann, D. Membrane remodeling, an early event in benzo[α]pyrene-induced apoptosis. Toxicol. Appl. Pharmacol. 2010, 243, 68–76. [Google Scholar] [CrossRef]
- Lee, J.; Park, A.; Kim, M.; Lim, H.-J.; Rha, Y.-A.; Kang, H.-G. Spirulina Extract Enhanced a Protective Effect in Type 1 Diabetes by Anti-Apoptosis and Anti-ROS Production. Nutrients 2017, 9, 1363. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.; Arlt, V.M.; Phillips, D.H. The role of cytochrome P450 enzymes in carcinogen activation and detoxication: An in vivo–in vitro paradox. Carcinogenesis 2018, 39, 851–859. [Google Scholar] [CrossRef]
- Shimada, T. Xenobiotic-metabolizing enzymes involved in activation and detoxification of carcinogenic polycyclic aromatic hydrocarbons. Drug Metab. Pharmacokinet. 2006, 21, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Oesterling, E.; Toborek, M.; Hennig, B. Benzo[a]pyrene induces intercellular adhesion molecule-1 through a caveolae and aryl hydrocarbon receptor mediated pathway. Toxicol. Appl. Pharmacol. 2008, 232, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Nickenig, G.; Werner, N. Extracellular Vesicles in Cardiovascular Disease: Potential Applications in Diagnosis, Prognosis, and Epidemiology. Circ. Res. 2017, 120, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Sturk, A.; Nieuwland, R. Extracellular vesicles in physiological and pathological conditions. Blood Rev. 2013, 27, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Goff, M.; Lagadic-Gossmann, D.; Latour, R.; Podechard, N.; Grova, N.; Gauffre, F.; Chevance, S.; Burel, A.; Appenzeller, B.M.R.; Ulmann, L.; et al. PAHs increase the production of extracellular vesicles both in vitro in endothelial cells and in vivo in urines from rats. Environ. Pollut. 2019, 255, 113171. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive Compounds Isolated from Microalgae in Chronic Inflammation and Cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Montero-Lobato, Z.; Vázquez, M.; Navarro, F.; Fuentes, J.; Bermejo, E.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae. Mar. Drugs 2018, 16, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Raposo, M.F.; de Morais, A.M. Microalgae for the prevention of cardiovascular disease and stroke. Life Sci. 2015, 125, 32–41. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Abdelnour, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Mahgoub, S.A.; Elnesr, S.S.; Gebriel, M.G. Microalgae in modern cancer therapy: Current knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef]
- Degraeve-Guilbault, C.; Bréhélin, C.; Haslam, R.; Sayanova, O.; Marie-Luce, G.; Jouhet, J.; Corellou, F. Glycerolipid Characterization and Nutrient Deprivation-Associated Changes in the Green Picoalga Ostreococcus tauri. Plant Physiol. 2017, 173, 2060–2080. [Google Scholar] [CrossRef] [Green Version]
- Di Nunzio, M.; Valli, V.; Tomás-Cobos, L.; Tomás-Chisbert, T.; Murgui-Bosch, L.; Danesi, F.; Bordoni, A. Is cytotoxicity a determinant of the different in vitro and in vivo effects of bioactives? BMC Complement. Altern. Med. 2017, 17, 453. [Google Scholar] [CrossRef] [Green Version]
- De Sousa Andrade, L.N.; de Lima, T.M.; Curi, R.; de Lauro Castrucci, A.M. Toxicity of fatty acids on murine and human melanoma cell lines. Toxicol. In Vitro 2005, 19, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.A.; Walker, C.L.; Pavlina, T.M.; Xu, Z.; Zaloga, G.P.; Siddiqui, R.A. Long-chain saturated fatty acids induce pro-inflammatory responses and impact endothelial cell growth. Clin. Nutr. 2010, 29, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Ucci, M.; Di Tomo, P.; Tritschler, F.; Cordone, V.G.P.; Lanuti, P.; Bologna, G.; Di Silvestre, S.; Di Pietro, N.; Pipino, C.; Mandatori, D.; et al. Anti-inflammatory Role of Carotenoids in Endothelial Cells Derived from Umbilical Cord of Women Affected by Gestational Diabetes Mellitus. Oxid. Med. Cell. Longev. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, A.; Asha, V.V.; Nair, S.A.; Sasidharan, S.P.; Sureshkumar, P.K.; Rajendran, K.N.; Karunagaran, D.; Ramalingam, K. Chlorophyll Revisited: Anti-inflammatory Activities of Chlorophyll a and Inhibition of Expression of TNF-α Gene by the Same. Inflammation 2012, 35, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Heo, S.-J.; Yoon, W.-J.; Kang, S.-M.; Ahn, G.; Yi, T.-H.; Jeon, Y.-J. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-inflammatory Effects of Violaxanthin Isolated from Microalga Chlorella ellipsoidea in RAW 264.7 Macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Caterina, R.; Cybulsky, M.I.; Clinton, S.K.; Gimbrone, M.A.; Libby, P. The omega-3 fatty acid docosahexaenoate reduces cytokine-induced expression of proatherogenic and proinflammatory proteins in human endothelial cells. Arterioscler. Thromb. J. Vasc. Biol. 1994, 14, 1829–1836. [Google Scholar] [CrossRef] [Green Version]
- Solanki, P.; Aminoshariae, A.; Jin, G.; Montagnese, T.A.; Mickel, A. The effect of docosahexaenoic acid (DHA) on expression of IL-1ß, IL-6, IL-8, and TNF-α in normal and lipopolysaccharide (LPS)-stimulated macrophages. Quintessence Int. 2013, 44, 393. [Google Scholar]
- De Souza, C.O.; Valenzuela, C.A.; Baker, E.J.; Miles, E.A.; Rosa Neto, J.C.; Calder, P.C. Palmitoleic Acid has Stronger Anti-Inflammatory Potential in Human Endothelial Cells Compared to Oleic and Palmitic Acids. Mol. Nutr. Food Res. 2018, 62, 1800322. [Google Scholar] [CrossRef]
- Erdinest, N.; Shmueli, O.; Grossman, Y.; Ovadia, H.; Solomon, A. Anti-Inflammatory Effects of Alpha Linolenic Acid on Human Corneal Epithelial Cells. Investig. Opthalmol. Vis. Sci. 2012, 53, 4396. [Google Scholar] [CrossRef]
- Liu, M.-H.; Lin, A.-H.; Lu, S.-H.; Peng, R.-Y.; Lee, T.-S.; Kou, Y.R. Eicosapentaenoic acid attenuates cigarette smoke-induced lung inflammation by inhibiting ROS-sensitive inflammatory signaling. Front. Physiol. 2014, 5, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, X.-H.; Nakanishi, Y.; Inoue, H.; Takayama, K.; Bai, F.; Hara, N. Polycyclic aromatic hydrocarbons induce IL-8 expression through nuclear factor κB activation in A549 cell line. Cytokine 2002, 19, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Badal, S.; Delgoda, R. Role of the modulation of CYP1A1 expression and activity in chemoprevention: Modulation of CYP1A1 expression and activity in chemoprevention. J. Appl. Toxicol. 2014, 34, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Wang, J.; Zhu, K.; Tang, Y.; Huang, S.; Shui, X.; Ding, Y.; Chen, C.; Lei, W. Aryl Hydrocarbon Receptor: A New Player of Pathogenesis and Therapy in Cardiovascular Diseases. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Satomi, Y.; Nishino, H. Inhibition of the enzyme activity of cytochrome P450 1A1, 1A2 and 3A4 by fucoxanthin, a marine carotenoid. Oncol. Lett. 2013, 6, 860–864. [Google Scholar] [CrossRef] [Green Version]
- Dendelé, B.; Tekpli, X.; Hardonnière, K.; Holme, J.A.; Debure, L.; Catheline, D.; Arlt, V.M.; Nagy, E.; Phillips, D.H.; Øvrebø, S.; et al. Protective action of n-3 fatty acids on benzo[a]pyrene-induced apoptosis through the plasma membrane remodeling-dependent NHE1 pathway. Chem. Biol. Interact. 2014, 207, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Gdula-Argasińska, J.; Czepiel, J.; Totoń-Żurańska, J.; Jurczyszyn, A.; Perucki, W.; Wołkow, P. Docosahexaenoic acid regulates gene expression in HUVEC cells treated with polycyclic aromatic hydrocarbons. Toxicol. Lett. 2015, 236, 75–81. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; E Baart, G.J.; De Cooman, L. Detection of flavonoids in microalgae from different evolutionary lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Denison, M.S.; Nagy, S.R. Activation of the Aryl Hydrocarbon Receptor by Structural Diverse Exogenous and Endogenous Chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef]
- Holme, J.A.; Gorria, M.; Arlt, V.M.; Øvrebø, S.; Solhaug, A.; Tekpli, X.; Landvik, N.E.; Huc, L.; Fardel, O.; Lagadic-Gossmann, D. Different mechanisms involved in apoptosis following exposure to benzo[a]pyrene in F258 and Hepa1c1c7 cells. Chem. Biol. Interact. 2007, 167, 41–55. [Google Scholar] [CrossRef]
- Chung, J.; Kim, J.; Kim, W.; Lee, S.; Kim, Y.; Park, J.; Hong, Y.; Chun, Y.; Park, Y.; Oh, S. Abundance of aryl hydrocarbon receptor potentiates benzo[a]pyrene-induced apoptosis in Hepa1c1c7 cells via CYP1A1 activation. Toxicology 2007, 235, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Tian, L.; Shen, T.; Sun, H.; Liu, P. Alpha-Lipoic Acid Protects Human Aortic Endothelial Cells Against H2O2-Induced Injury and Inhibits Atherosclerosis in Ovariectomized Low Density Lipoprotein Receptor Knock-Out Mice. Cell. Physiol. Biochem. 2018, 47, 2261–2277. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.A.; Walker, C.L.; Xu, Z.; Whitley, P.; Pavlina, T.M.; Hise, M.; Zaloga, G.P.; Siddiqui, R.A. Oleic acid inhibits stearic acid-induced inhibition of cell growth and pro-inflammatory responses in human aortic endothelial cells. J. Lipid Res. 2010, 51, 3470–3480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, F.; Li, Q.; Pfeifer, A.; Werner, N. Endothelial- and Immune Cell-Derived Extracellular Vesicles in the Regulation of Cardiovascular Health and Disease. JACC Basic Transl. Sci. 2017, 2, 790–807. [Google Scholar] [CrossRef]
- Van Meteren, N.; Lagadic-Gossmann, D.; Chevanne, M.; Gallais, I.; Gobart, D.; Burel, A.; Bucher, S.; Grova, N.; Fromenty, B.; Appenzeller, B.M.R.; et al. Polycyclic Aromatic Hydrocarbons Can Trigger Hepatocyte Release of Extracellular Vesicles by Various Mechanisms of Action Depending on Their Affinity for the Aryl Hydrocarbon Receptor. Toxicol. Sci. 2019, 171, 443–462. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Mayneris-Perxachs, J.; Lovegrove, J.A.; Todd, S.; Yaqoob, P. Fish-oil supplementation alters numbers of circulating endothelial progenitor cells and microparticles independently of eNOS genotype. Am. J. Clin. Nutr. 2014, 100, 1232–1243. [Google Scholar] [CrossRef] [Green Version]
- Muller-Feuga, A.; Lemar, M.; Vermel, E.; Pradelles, R.; Rimbaud, L.; Valiorgue, P. Appraisal of a horizontal two-phase flow photobioreactor for industrial production of delicate microalgae species. J. Appl. Phycol. 2012, 24, 349–355. [Google Scholar] [CrossRef]
- Muller-Feuga, A. Réacteur Photosynthétique Pour la Culture de Microorganismes et Procédé de Culture de Microorganismes. Patent No. WO/2010/109108, 30 September 2010. [Google Scholar]
- Heydarizadeh, P.; Boureba, W.; Zahedi, M.; Huang, B.; Moreau, B.; Lukomska, E.; Couzinet-Mossion, A.; Wielgosz-Collin, G.; Martin-Jézéquel, V.; Bougaran, G.; et al. Response of CO2-starved diatom Phaeodactylum tricornutum to light intensity transition. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160396. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. ISBN 978-0-12-182048-0. [Google Scholar]
- Schoefs, B. Determination of pigments in vegetables. J. Chromatogr. A 2004, 1054, 217–226. [Google Scholar] [CrossRef]
- Schoefs, B.; Franck, F. Chlorophyll Synthesis in Dark-Grown Pine Primary Needles. Plant Physiol. 1998, 118, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W. (Ed.) Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods, 2nd ed.; Monographs on Oceanographic Methodology; Unesco Publ: Paris, France, 2005; ISBN 978-92-3-103275-2. [Google Scholar]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chlorophylls | Pigment Concentration (g/L) | |
|---|---|---|
| PT | OT | |
| Chlorophyll a | 5.08 | 2.51 |
| Chorophyll b * | - | 1.64 |
| Chorophyll c * | 0.47 | - |
| Carotenoids | Absorbance Maxima (nm) | Absorbance Maxima in Literature (nm) | Pigment Concentration (g/L) | |
|---|---|---|---|---|
| PT | OT | |||
| Total carotenoids | 2.47 | 1.00 | ||
| β-carotene | 424,454,484 | (429),454,480 | 0.03 | 0.05 |
| Fucoxanthin | (415),443,472 | (420),447,468 | 2.13 | - |
| Other carotenoids | ||||
| Dihydrolutein | (−),432,462 | 408,428,453 | - | 0.08 |
| Micromonal | 422,451,477 | (423),449,(472) | - | 0.43 |
| Prasinoxanthin | 420,449,(−) | (426),459,(473) | - | 0.10 |
| Neochrome | (−),414,442 | 398,421,448 | - | 0.10 |
| Fatty Acids (mol %) | P. tricornutum | O. tauri | ||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| C14:0 | 1.18 | 0.43 | 4.20 | 2.80 |
| C16:0 | 7.68 | 0.20 | 14.64 | 0.53 |
| C16:1n-7 | 18.27 | 0.96 | 8.68 | 0.91 |
| C16:2 | 5.28 | 0.29 | ND | - |
| C16:4 | 11.23 | 1.38 | ND | - |
| C18:0 | 1.88 | 0.32 | 10.91 | 2.69 |
| C18:1 | 1.87 | 0.35 | 23.06 | 3.01 |
| C18:2n-6 | 2.97 | 0.11 | 4.40 | 0.43 |
| C18:3n-3 | ND | - | 8.56 | 0.86 |
| C18:4n-3 | ND | - | 11.56 | 0.82 |
| C20:1 | ND | - | 0.83 | 0.07 |
| C20:2 | 2.08 | 0.26 | ND | - |
| C20:4 | 3.34 | 0.16 | 0.97 | 0.26 |
| C20:5n-3 | 39.91 | 2.91 | 1.16 | 0.18 |
| C22:6n-3 | 1.72 | 0.11 | 7.69 | 1.26 |
| Gene Symbol | Forward Sequence | Reverse Sequence |
|---|---|---|
| CYP1A1 | CCCACAGCACAACAAGAGACA | CATCAGGGGTGAGAAACCGT |
| IL-8 | ACTCCAAACCTTTCCACCCC | TCTCAGCCCTCTTCAAAAACTTC |
| IL1-β | CTCTGGGATTCTCTTCAGCCA | AGGAGCACTTCATCTGTTTAGGG |
| 18S | CGCCGCTAGAGGTGAAATTC | TTGGCAAATGCTTTCGCTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Goff, M.; Delbrut, A.; Quinton, M.; Pradelles, R.; Bescher, M.; Burel, A.; Schoefs, B.; Sergent, O.; Lagadic-Gossmann, D.; Le Ferrec, E.; et al. Protective Action of Ostreococcus Tauri and Phaeodactylum Tricornutum Extracts towards Benzo[a]Pyrene-Induced Cytotoxicity in Endothelial Cells. Mar. Drugs 2020, 18, 3. https://doi.org/10.3390/md18010003
Le Goff M, Delbrut A, Quinton M, Pradelles R, Bescher M, Burel A, Schoefs B, Sergent O, Lagadic-Gossmann D, Le Ferrec E, et al. Protective Action of Ostreococcus Tauri and Phaeodactylum Tricornutum Extracts towards Benzo[a]Pyrene-Induced Cytotoxicity in Endothelial Cells. Marine Drugs. 2020; 18(1):3. https://doi.org/10.3390/md18010003
Chicago/Turabian StyleLe Goff, Manon, Antoine Delbrut, Marie Quinton, Rémi Pradelles, Maelle Bescher, Agnès Burel, Benoît Schoefs, Odile Sergent, Dominique Lagadic-Gossmann, Eric Le Ferrec, and et al. 2020. "Protective Action of Ostreococcus Tauri and Phaeodactylum Tricornutum Extracts towards Benzo[a]Pyrene-Induced Cytotoxicity in Endothelial Cells" Marine Drugs 18, no. 1: 3. https://doi.org/10.3390/md18010003
APA StyleLe Goff, M., Delbrut, A., Quinton, M., Pradelles, R., Bescher, M., Burel, A., Schoefs, B., Sergent, O., Lagadic-Gossmann, D., Le Ferrec, E., & Ulmann, L. (2020). Protective Action of Ostreococcus Tauri and Phaeodactylum Tricornutum Extracts towards Benzo[a]Pyrene-Induced Cytotoxicity in Endothelial Cells. Marine Drugs, 18(1), 3. https://doi.org/10.3390/md18010003