Differential Expression of Nicotine Acetylcholine Receptors Associates with Human Breast Cancer and Mediates Antitumor Activity of αO-Conotoxin GeXIVA

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. The LBD cDNA Cloning of Each nAChR Subunit in Various Human Breast Cell Lines
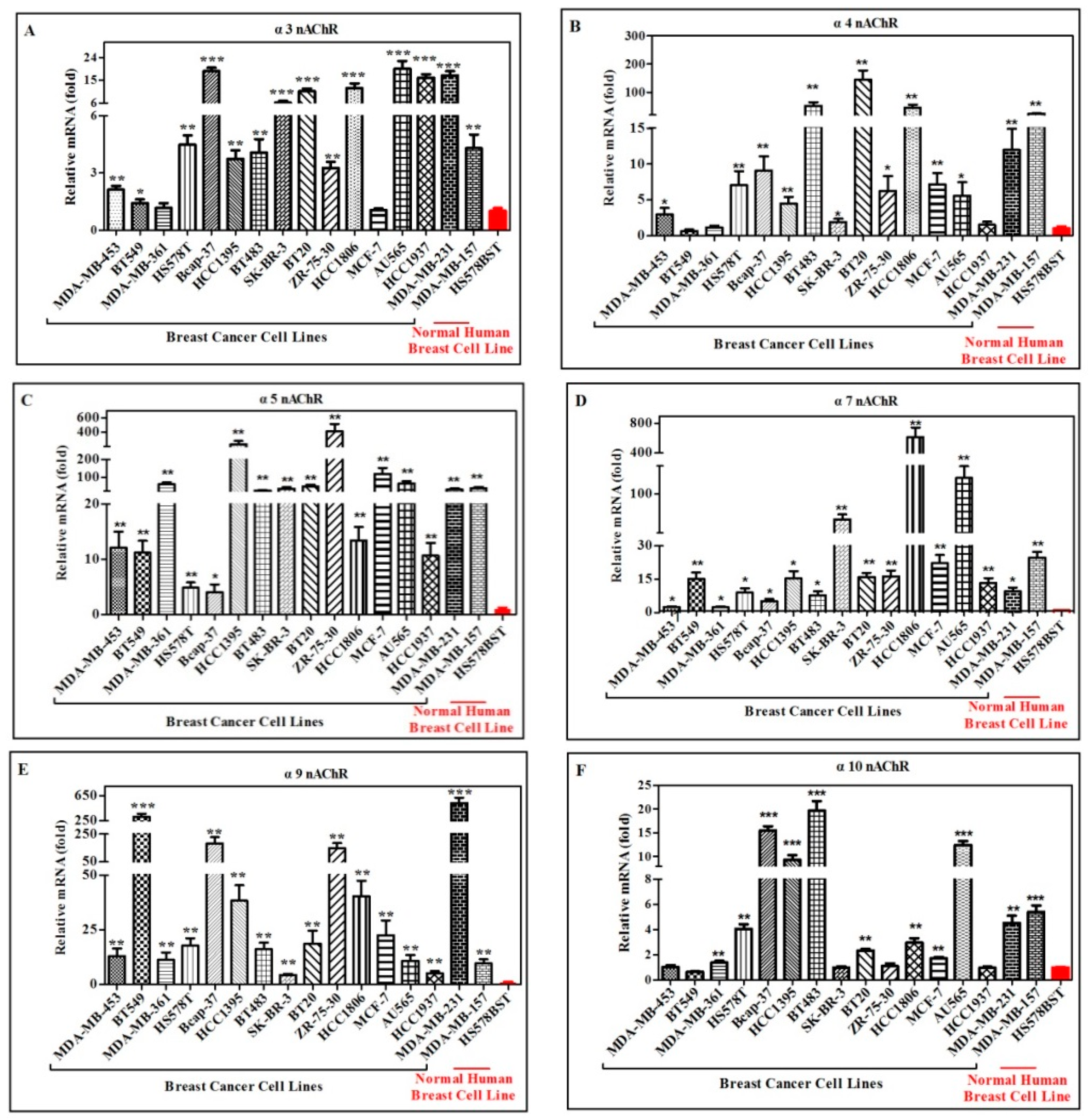
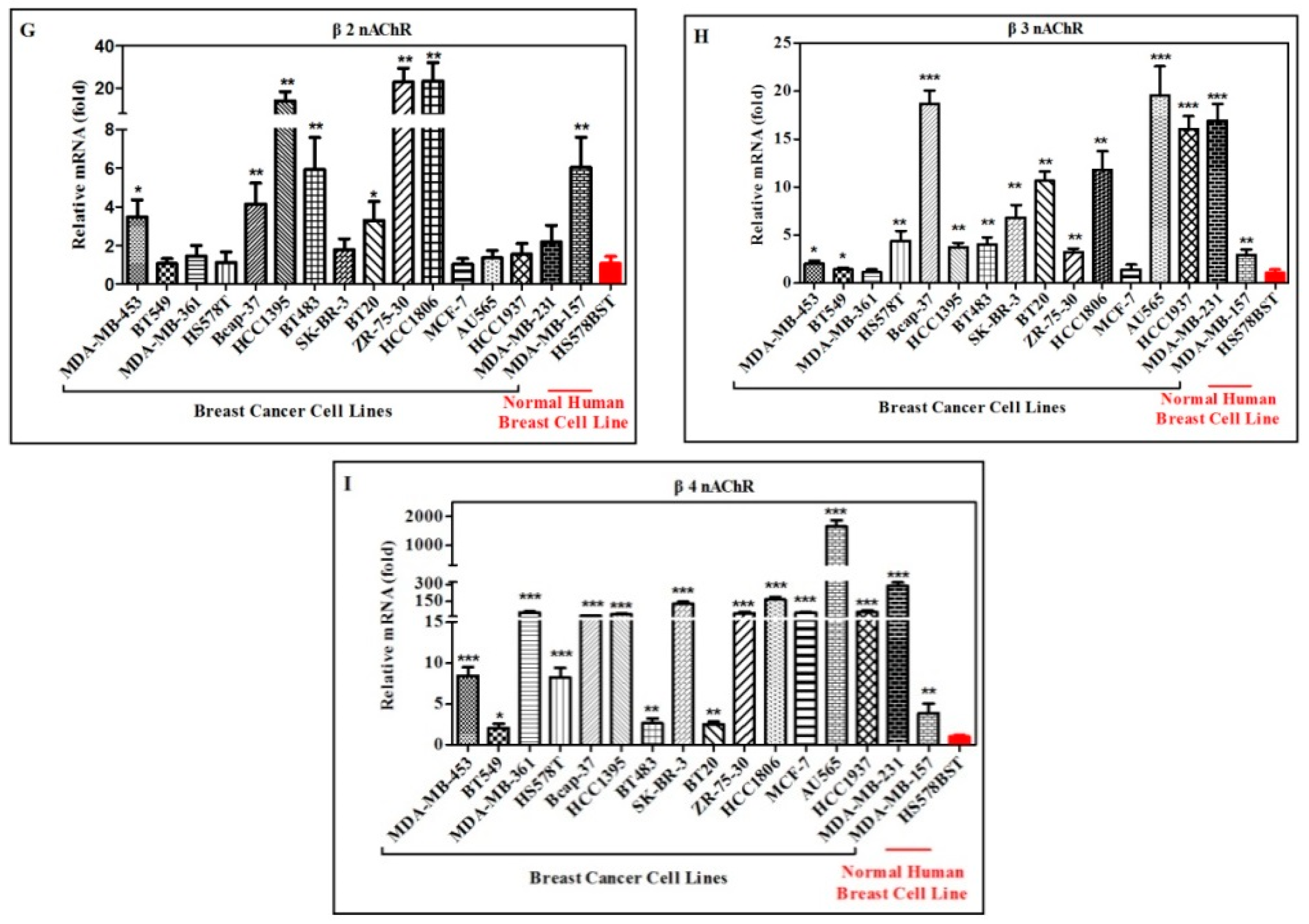
2.2. Quantification and Comparison of Different nAChR Subunits mRNA in Different Human Breast Cancer and Normal Cell Lines
2.3. The Functional nAChRs Expression on the Surface of Breast Cancer Cells and Mammary Gland Epithelial Cells
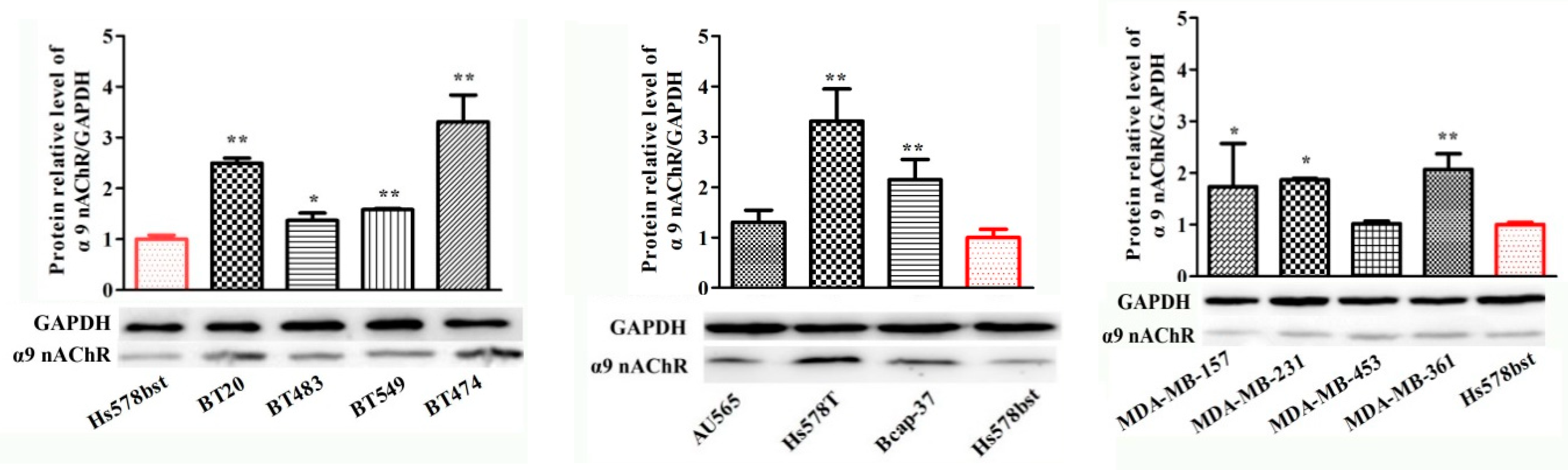
2.4. Protein Expression of α9-Containing Subunit nAChRs in Different Human Breast Cancer Cell Lines
2.5. αO-Conotoxin GeXIVA Affects Breast Cancer Cell Proliferation through the Inhibition of α9-nAChR-Mediated Signals
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Isolation and RT-PCR Amplification of Ligand Binding Domain (LBD) of Different Human nAChR Subunits
4.3. Quantitative Real-Time PCR
4.4. Protein Extraction and Western Blot Analysis
4.5. Flow Cytometry Analysis of Functional nAChRs Expression on the Cell Surface
4.6. Cell Viability
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [PubMed] [Green Version]
- WHO. Cancer What Is Cancer? Available online: http://www.who.int/cancer/en/ (accessed on 15 January 2020).
- Wanqing, C.; Rongshou, Z.; Siwei, Z.; Tingting, Z.; Hongmei, Z.; Jie, H. Report of cancer incidence and mortality in China, 2012. China Cancer 2016, 24, 1–7. [Google Scholar]
- Schuller, H.M. Is cancer triggered by altered signalling of nicotinic acetylcholine receptors? Nat. Rev. Cancer 2009, 9, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Dang, N.; Meng, X.; Song, H. Nicotinic acetylcholine receptors and cancer. Biomed. Rep. 2016, 4, 515–518. [Google Scholar] [PubMed] [Green Version]
- Cascio, M. Structure and function of the glycine receptor and related nicotinicoid receptors. J. Biol. Chem. 2004, 279, 19383–19386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, E.X.; Pereira, E.F.R.; Manickavasagom, A.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [PubMed] [Green Version]
- Bychkov, M.; Shenkarev, Z.; Shulepko, M.; Shlepova, O.; Kirpichnikov, M.; Lyukmanova, E. Water-Soluble variant of human Lynx1 induces cell cycle arrest and apoptosis in lung cancer cells via modulation of alpha7 nicotinic acetylcholine receptors. PLoS ONE 2019, 14, e0217339. [Google Scholar] [CrossRef]
- Grando, S.A. Connections of nicotine to cancer. Nat. Rev. Cancer 2014, 14, 419–429. [Google Scholar] [CrossRef]
- Scherl, C.; SchãFer, R.; Schlabrakowski, A.; Tziridis, K.; Iro, H.; Wendler, O. Nicotinic acetylcholine receptors in head and neck cancer and their correlation to tumor site and progression. ORL J. Otorhinolaryngol. Relat. Spec. 2016, 78, 151–158. [Google Scholar] [CrossRef]
- Lyukmanova, E.N.; Bychkov, M.L.; Sharonov, G.V.; Efremenko, A.V.; Shulepko, M.A.; Kulbatskii, D.S.; Shenkarev, Z.O.; Feofanov, A.V.; Dolgikh, D.A.; Kirpichnikov, M.P. Human secreted proteins SLURP-1 and SLURP-2 control the growth of epithelial cancer cells via interactions with nicotinic acetylcholine receptors. Br. J. Pharmacol. 2018, 175, 1973–1986. [Google Scholar]
- Improgo, M.R.; Soll, L.G.; Tapper, A.R.; Gardner, P.D. Nicotinic acetylcholine receptors mediate lung cancer growth. Front. Physiol. 2013, 4, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, Z.; Małgorzata, K.; Dominika, U.; Aneta, L.; Sławomir, L. Role of α7 nicotinic receptor in the immune system and intracellular signaling pathways. Cent. Eur. J. Immunol. 2015, 40, 373–379. [Google Scholar]
- Sun, H.; Ma, X. α5-nAChR modulates nicotine-induced cell migration and invasion in A549 lung cancer cells. Exp. Toxicol. Pathol. 2015, 67, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Lee, C.H.; Wu, C.H. The Alpha 9-Nicotinic Acetylcholine Receptor Serves as a Molecular Target for Breast Cancer Therapy. J. Exp. Clin. Med. 2011, 3, 246–251. [Google Scholar] [CrossRef]
- Calleja-Macias, I.E.; Kalantari, M.; Bernard, H.U. Cholinergic signaling through nicotinic acetylcholine receptors stimulates the proliferation of cervical cancer cells: An explanation for the molecular role of tobacco smoking in cervical carcinogenesis? Int. J. Cancer 2009, 124, 1090–1096. [Google Scholar] [CrossRef]
- Grozio, A.; Paleari, L.; Catassi, A.; Servent, D.; Cilli, M.; Piccardi, F.; Paganuzzi, M.; Cesario, A.; Granone, P.; Mourier, G. Natural agents targeting the alpha7-nicotinic-receptor in NSCLC: A promising prospective in anti-cancer drug development. Int. J. Cancer 2010, 122, 1911–1915. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, X.; Zhao, Q.; Yang, X.; An, S.; Wang, H.; Xu, L.; Zhu, L.; Chen, H. Role of α7-nicotinic acetylcholine receptor in nicotine-induced invasion and epithelial-to-mesenchymal transition in human non-small cell lung cancer cells. Oncotarget 2016, 7, 59199–59208. [Google Scholar]
- Lee, C.H.; Huang, C.S.; Chen, C.S.; Tu, S.H.; Wang, Y.J.; Chang, Y.J.; Tam, K.W.; Wei, P.L.; Cheng, T.C.; Chu, J.S. Overexpression and activation of the α9-nicotinic receptor during tumorigenesis in human breast epithelial cells. J. Natl. Cancer Inst. 2010, 102, 1322–1335. [Google Scholar] [CrossRef]
- Huang, L.C.; Lin, C.L.; Qiu, J.Z.; Lin, C.Y.; Hsu, K.W.; Tam, K.W.; Lee, J.Y.; Yang, J.M.; Lee, C.H. Nicotinic acetylcholine receptor subtype alpha-9 mediates triple-negative breast cancers based on a spontaneous pulmonary metastasis mouse model. Front. Cell. Neurosci. 2017, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2017, 679, 24–34. [Google Scholar] [CrossRef]
- Hone, A.J.; Mcintosh, J.M.; Azam, L.; Lindstrom, J.; Lucero, L.; Whiteaker, P.; Passas, J.; Blã, Z.J.; Albillos, A. α-conotoxins identify the α3β4* subtype as the predominant nicotinic acetylcholine receptor expressed in human adrenal chromaffin cells. Mol. Pharmacol. 2015, 88, 881–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannon, H.; Atchison, W. Omega-conotoxins as experimental tools and therapeutics in pain management. Mar. Drugs 2013, 11, 680–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essack, M.; Bajic, V.B.; Archer, J.A.C. Conotoxins that confer therapeutic possibilities. Mar. Drugs 2012, 10, 1244–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, L.; McIntosh, J. Conotoxins: Therapeutic potential and application. Mar. Drugs 2006, 4, 119–142. [Google Scholar]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kaas, Q.; Wu, Y.; Zhu, X.; Hu, Y.; Li, X.; Tsetlin, V.I.; Christensen, S.; et al. Cloning, synthesis and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, E4026–E4035. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, X.D.; Zhangsun, D.; Yu, G.; Su, R.; Luo, S. The alpha9alpha10 nicotinic acetylcholine receptor antagonist alpha O-conotoxin GeXIVA[1,2] alleviates and reverses chemotherapy-induced neuropathic pain. Mar. Drugs 2019, 17, 265. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Hu, Y.; Wu, Y.; Huang, Y.; Yu, S.; Ding, Q.; Zhangsun, D.; Luo, S. Anti-hypersensitive effect of intramuscular administration of αO-conotoxin GeXIVA[1,2] and GeXIVA[1,4] in rats of neuropathic pain. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 66, 112–119. [Google Scholar] [CrossRef]
- Liu, Y.; Qian, J.; Sun, Z.; Zhangsun, D.; Luo, S. Cervical cancer correlates with the differential expression of nicotinic acetylcholine receptors and reveals therapeutic targets. Mar. Drugs 2019, 17, 256. [Google Scholar] [CrossRef] [Green Version]
- Li, S.X.; Huang, S.; Bren, N.; Noridomi, K.; Dellisanti, C.D.; Sine, S.M.; Chen, L. Ligand-binding domain of an α7-nicotinic receptor chimera and its complex with agonist. Nat. Neurosci. 2011, 14, 1253–1259. [Google Scholar] [CrossRef] [Green Version]
- Morales-Perez, C.L.; Noviello, C.M.; Hibbs, R.E. X-ray structure of the human alpha4beta2 nicotinic receptor. Nature 2016, 538, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y. The oncogenic functions of nicotinic acetylcholine receptors. J. Oncol. 2016, 2016, 9650481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Chang, Y.C.; Chen, C.S.; Tu, S.H.; Wang, Y.J.; Chen, L.C.; Chang, Y.J.; Wei, P.L.; Chang, H.W.; Chang, C.H. Crosstalk between nicotine and estrogen-induced estrogen receptor activation induces α9-nicotinic acetylcholine receptor expression in human breast cancer cells. Breast Cancer Res. Treat. 2011, 129, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Kalantari-Dehaghi, M.; Parnell, E.A.; Armand, T.; Bernard, H.-U.; Grando, S.A. The nicotinic acetylcholine receptor-mediated reciprocal effects of the tobacco nitrosamine NNK and SLURP-1 on human mammary epithelial cells. Int. Immunopharmacol. 2015, 29, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Lee, C.H.; Hsieh, C.D.; Ho, C.T.; Pan, M.H.; Huang, C.S.; Tu, S.H.; Wang, Y.J.; Chen, L.C.; Chang, Y.J. Nicotine-induced human breast cancer cell proliferation attenuated by garcinol through down-regulation of the nicotinic receptor and cyclin D3 proteins. Breast Cancer Res. Treat. 2011, 125, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Hirata, N.; Sekino, Y.; Kanda, Y. Role of α7-nicotinic acetylcholine receptor in normal and cancer stem cells. Curr. Drug Targets 2012, 13, 656–665. [Google Scholar] [PubMed]
- Tait, L.; Soule, H.D.; Russo, J. Ultrastructural and immunocytochemical characterization of an immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6087–6094. [Google Scholar] [PubMed]
- Gerzanich, V.; Wang, F.; Kuryatov, A.; Lindstrom, J. alpha 5 subunit alters desensitization, pharmacology, Ca++ permeability and Ca++ modulation of human neuronal alpha 3 nicotinic receptors. J. Pharmacol. Exp. Ther. 1998, 286, 311–320. [Google Scholar]
- Wojaskrawczyk, K.; Krawczyk, P.; Biernacka, B.; Grzybek, M.; Kołodziej, P.; Kucharczyk, T.; Mlak, R.; Milanowski, J. The polymorphism of the CHRNA5 gene and the strength of nicotine addiction in lung cancer and COPD patients. Eur. J. Cancer Prev. 2012, 21, 111–117. [Google Scholar] [CrossRef]
- Ma, X.; Jia, Y.; Zu, S.; Li, R.; Jia, Y.; Zhao, Y.; Xiao, D.; Dang, N.; Wang, Y. α5 nicotinic acetylcholine receptor mediates nicotine-induced HIF-1α and VEGF expression in non-small cell lung cancer. Toxicol. Appl. Pharmacol. 2014, 278, 172–179. [Google Scholar] [CrossRef]
- Sandeep, S.; Smitha, P.; Srikumar, C. Nicotinic acetylcholine receptor signaling in tumor growth and metastasis. J. Oncol. 2011, 2011, 456743. [Google Scholar]
- Gankhuyag, N.; Lee, K.-H.; Cho, J.-Y. The role of nitrosamine (NNk) in breast cancer carcinogenesis. J. Mammary Gland Biol. Neoplasia 2017, 22, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Thapa, P.; Espiritu, M.J.; Cabalteja, C.; Bingham, J.P. The emergence of cyclic peptides: The potential of bioengineered peptide drugs. Int. J. Pept. Res. Ther. 2014, 20, 545–551. [Google Scholar] [CrossRef]
- Luetje, C.W.; Wada, K.; Rogers, S.; Abramson, S.N.; Tsuji, K.; Heinemann, S.; Patrick, J. Neurotoxins distinguish between different neuronal nicotinic acetylcholine receptor subunit combinations. J. Neurochem. 1990, 55, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.S.; Peng, Y.J.; Wei, P.L.; Lee, C.H.; Su, H.Y.; Ho, Y.S.; Lin, S.Y.; Wu, C.H.; Chang, Y.J. The alpha9 nicotinic acetylcholine receptor is the key mediator in nicotine-enhanced cancer metastasis in breast cancer cells. J. Exp. Clin. Med. 2011, 3, 283–292. [Google Scholar] [CrossRef]
- Eline, L.; Steve, P.; Isuru, W.; Jan, T. Conotoxins targeting nicotinic acetylcholine receptors: An overview. Mar. Drugs 2014, 12, 2970–3004. [Google Scholar]
- Azam, L.; Mcintosh, J.M. α-Conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; Mcintosh, J.M. Characterization of a novel alpha-conotoxin TxID from conus textile that potently blocks rat alpha3beta4 nicotinic acetylcholine receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhangsun, D.; Zhu, X.; Kaas, Q.; Zhangsun, M.; Harvey, P.J.; Craik, D.J.; Mcintosh, J.M.; Luo, S. α-Conotoxin [S9A]TxID potently discriminates between α3β4 and α6/α3β4 nicotinic acetylcholine receptors. J. Med. Chem. 2017, 60, 5826–5833. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhu, X.; Harvey, P.J.; Kaas, Q.; Zhangsun, D.; Craik, D.J.; Luo, S. Single amino acid substitution in alpha-conotoxin TxID reveals a specific alpha3beta4 nicotinic acetylcholine receptor antagonist. J. Med. Chem. 2018, 61, 9256–9265. [Google Scholar] [CrossRef]
- Qian, J.; Liu, Y.-Q.; Sun, Z.-H.; Zhangsun, D.-T.; Luo, S.-L. Identification of nicotinic acetylcholine receptor subunits in different lung cancer cell lines and the inhibitory effect of alpha-conotoxin TxID on lung cancer cell growth. Eur. J. Pharmacol. 2019, 865, 172674. [Google Scholar] [CrossRef]
- Peng, C.; Tang, S.; Pi, C.; Liu, J.; Wang, F.; Wang, L.; Zhou, W.; Xu, A. Discovery of a novel class of conotoxin from conus litteratus, lt14a, with a unique cysteine pattern. Peptides 2006, 27, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Vincler, M.; Wittenauer, S.; Parker, R.; Ellison, M.; Olivera, B.M.; McIntosh, J.M. Molecular mechanism for analgesia involving specific antagonism of α9α10 nicotinic acetylcholine receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 17880–17884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannelli, L.D.C.; Cinci, L.; Micheli, L.; Zanardelli, M.; Pacini, A.; Mcintosh, J.M.; Ghelardini, C. α-Conotoxin RgIA protects against the development of nerve injury-induced chronic pain and prevents both neuronal and glial derangement. Pain 2014, 155, 1986–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhangsun, D.; Zhu, X.; Kaas, Q.; Wu, Y.; Craik, D.J.; McIntosh, J.M.; Luo, S. αO-Conotoxin GeXIVA disulfide bond isomers exhibit differential sensitivity for various nicotinic acetylcholine receptors but retain potency and selectivity for the human α9α10 subtype. Neuropharmacology 2017, 127, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Yungleun, S.; Liu, H.C.; Chen, C.S.; Chunghuei, H.; Pan, M.H.; Chang, H.W.; Chang, C.H.; Chen, F.C.; Chitang, H.; Yang, Y.Y. Combination treatment with luteolin and quercetin enhances antiproliferative effects in nicotine-treated MDA-MB-231 cells by down-regulating nicotinic acetylcholine receptors. J. Agric. Food Chem. 2010, 58, 235–241. [Google Scholar]
- Tu, S.H.; Ku, C.Y.; Ho, C.T.; Chen, C.S.; Huang, C.S.; Lee, C.H.; Chen, L.C.; Pan, M.H.; Chang, H.W.; Chang, C.H. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits nicotine- and estrogen-induced α9-nicotinic acetylcholine receptor upregulation in human breast cancer cells. Mol. Nutr. Food Res. 2015, 55, 455–466. [Google Scholar] [CrossRef]
- Bubner, B.; Baldwin, I.T. Use of real-time PCR for determining copy number and zygosity in transgenic plants. Plant Cell Rep. 2004, 23, 263–271. [Google Scholar] [CrossRef]
- Katchman, B.A.; Ocal, I.T.; Cunliffe, H.E.; Chang, Y.H.; Hostetter, G.; Watanabe, A.; Lobello, J.; Lake, D.F. Expression of quiescin sulfhydryl oxidase 1 is associated with a highly invasive phenotype and correlates with a poor prognosis in Luminal B breast cancer. Breast Cancer Res. 2013, 15, R28. [Google Scholar] [CrossRef] [Green Version]
- Yousef, E.M.; Laperrière, D.; Ramzan-Tahir, M.; Mader, S.; Gaboury, L.A. Deregulated expression of ANXA1 in human high-grade breast cancers. J. Mol. Biomark. Diagn. 2003, 4, 2. [Google Scholar]
- Cailleau, R.; Young, R.; Olivé, M.; Reeves, J.R.W. Breast tumor cell lines from pleural effusions. J. Natl. Cancer Inst. 1974, 53, 661–674. [Google Scholar] [CrossRef]
- Hackett, A.J.; Smith, H.S.; Springer, E.L.; Owens, R.B.; Nelsonrees, W.A.; Riggs, J.L.; Gardner, M.B. Two syngeneic cell lines from human breast tissue: The aneuploid mammary epithelial (Hs578t) and the diploid myoepithelial (Hs578bst) cell lines. J. Natl. Cancer Inst. 1977, 58, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Zhang, J.Q.; Wang, S.X. Establishment and investigation biological functions of breast cancer cell lines Bcap-37. J. Peking Univ. (Med. Ed.) 1983, 15, 161–164. [Google Scholar]
- Gazdar, A.; Kurvari, V.; Virmani, A.; Gollahon, L.; Sakaguchi, M.; Westerfield, M.; Kodagoda, D.; Stasny, V.; Cunningham, H.; Wistuba, I. Characterization of paired tumor and non-tumor cell lines established from patients with breast cancer. Int. J. Cancer 2015, 78, 766–774. [Google Scholar] [CrossRef]
- Lasfargues, E.Y.; Coutinho, W.G.; Redfield, E.S. Isolation of two human tumor epithelial cell lines from solid breast carcinomas. J. Natl. Cancer Inst. 1978, 61, 967–978. [Google Scholar]
- Subik, K.; Lee, J.F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.C.; Bonfiglio, T.; Hicks, D.G. The expression patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by immunohistochemical analysis in breast cancer cell lines. Breast Cancer Basic Clin. Res. 2010, 4, 35–41. [Google Scholar] [CrossRef]
- Trempe, G.L. Human breast cancer in culture. Recent Results Cancer Res. 1976, 57, 33–41. [Google Scholar]
- Lasfargues, E.Y.; Ozzello, L. Cultivation of human breast carcinomas. J. Natl. Cancer Inst. 1958, 21, 1131–1147. [Google Scholar]
- Engel, L.W.; Young, N.A.; Tralka, T.S.; Lippman, M.E.; O’Brien, S.J.; Joyce, M.J. Establishment and characterization of three new continuous cell lines derived from human breast carcinomas. Cancer Res. 1978, 38, 3352–3364. [Google Scholar]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Tomlinson, G.E.; Chen, T.T.; Stastny, V.A.; Virmani, A.K.; Spillman, M.A.; Tonk, V.; Blum, J.L.; Schneider, N.R.; Wistuba, I.I.; Shay, J.W.; et al. Characterization of a breast cancer cell line derived from a germ-line BRCA1 mutation carrier. Cancer Res. 1998, 58, 3237–3242. [Google Scholar]
- Young, R.K.; Cailleau, R.M.; Mackay, B.; Reeves, W.J. Establishment of epithelial cell line MDA-MB-157 from metastatic pleural effusion of human breast carcinoma. In Vitro 1974, 9, 239–245. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | nAChR Subunits | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Name | α3 | α4 | α5 | α7 | α9 | α10 | β2 | β3 | β4 |
| MDA-MB-453 | ++ | + | ++ | + | ++ | ns | ++ | + | +++ |
| BT-549 | + | ns | ++ | ++ | +++ | ns | ns | + | + |
| MDA-MB-361 | ns | ns | ++ | + | ++ | ++ | ns | ns | +++ |
| HS578T | ++ | ++ | + | + | ++ | ++ | ns | ++ | +++ |
| Bcap-37 | ++ | ++ | + | + | ++ | +++ | ++ | +++ | +++ |
| HCC1395 | ++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | +++ |
| BT-483 | ++ | ++ | ++ | + | ++ | +++ | ++ | ++ | ++ |
| SK-BR-3 | +++ | + | ++ | ++ | ++ | ns | ns | ++ | +++ |
| BT-20 | +++ | ++ | ++ | ++ | ++ | ++ | + | ++ | ++ |
| ZR-75-30 | ++ | + | ++ | ++ | ++ | ns | ++ | ++ | +++ |
| HCC1806 | +++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | +++ |
| MCF-7 | ns | ++ | ++ | ++ | ++ | ++ | ns | ns | +++ |
| AU565 | +++ | + | ++ | ++ | ++ | +++ | ns | +++ | +++ |
| HCC1937 | +++ | ns | ++ | ++ | + | ns | ns | +++ | +++ |
| MDA-MB-231 | +++ | ++ | ++ | + | +++ | ++ | + | +++ | +++ |
| MDA-MB-157 | ++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | ++ |
| HS578BST | - | - | - | - | - | - | - | - | - |
| Cell Lines | nAChR Subunits (Protein Level) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Name | α3 | α4 | α5 | α6 | α7 | α9 | α10 | β2 | β3 | β4 |
| MDA-MB-453 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| BT-549 | ns | ns | ns | ns | ns | +++ | ns | ns | ns | ns |
| MDA-MB-361 | ns | + | + | ns | ns | +++ | ns | ns | ns | ns |
| HS578T | ns | ns | ns | ns | ns | +++ | ns | ns | +++ | ns |
| Bcap-37 | ns | ns | ns | ns | ns | +++ | ns | ns | ++ | ns |
| HCC1395 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| BT-483 | ns | ns | +++ | ++ | + | +++ | ns | ns | +++ | ns |
| SK-BR-3 | ns | + | + | ns | ns | +++ | ns | ns | ns | + |
| BT-20 | ns | ++ | + | + | ++ | +++ | ns | ns | +++ | ns |
| ZR-75-30 | ++ | + | ++ | + | + | +++ | +++ | ++ | ns | ++ |
| HCC1806 | +++ | +++ | +++ | +++ | ns | +++ | ++ | ns | ++ | ns |
| MCF-7 | ns | ns | ns | ns | ns | +++ | ++ | ns | ns | ns |
| AU565 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ++ | ns | +++ |
| HCC1937 | +++ | ns | ns | ns | ns | +++ | ns | ns | ++ | ns |
| MDA-MB-231 | ++ | ++ | ns | ns | ++ | +++ | ns | ns | ns | ++ |
| MDA-MB-157 | +++ | +++ | ++ | +++ | +++ | +++ | +++ | ns | +++ | +++ |
| BT474 | +++ | ns | ++ | ns | + | +++ | ns | ns | ++ | + |
| HS578BST | − | − | − | − | − | − | − | − | − | |
| Treatment | IC50 (uM) | Control | 11.25 μM | 22.5 μM | 45 μM | 90 μM | 180 μM |
|---|---|---|---|---|---|---|---|
| Cells | Mean ± SD | ||||||
| MDA-MB-157 | 35.18 | 100 ± 2.82 d | 72.19 ± 3.53 c | 68.14 ± 2.44 c | 54.83 ± 3.27 b | 51.33 ± 3.23 b | 45.37 ± 2.91 a |
| MDA-MB-231 | 125.1 | 100 ± 5.61 e | 88.07 ± 1.69 d | 85.88 ± 3.41 d | 78.92 ± 1.41 c | 65.65 ± 2.63 b | 39.93 ± 1.87 a |
| MDA-MB-361 | 77.03 | 100 ± 5.35 c | 78.58 ± 6.30 b | 75.48 ± 6.82 b | 70.67 ± 6.32 b | 64.57 ± 6.77 b | 33.34 ± 1.84 a |
| MDA-MB-453 | 65.96 | 100 ± 4.58 d | 93.14 ± 1.51 c | 91.72 ± 0.58 c | 81.52 ± 3.30 b | 75.41 ± 4.02 b | 65.47 ± 9.63 a |
| HCC1395 | 73.50 | 100 ± 0.80 f | 77.79 ± 0.46 e | 73.15 ± 0.59 d | 68.21 ± 1.32 c | 57.47 ± 1.75 b | 49.93 ± 4.03 a |
| HCC1937 | 127.8 | 100 ± 1.89 d | 94.11 ± 1.71 d | 93.92 ± 1.43 d | 80.53 ± 7.43 c | 67.17 ± 1.53 b | 28.66 ± 0.48 a |
| HCC1806 | 110.7 | 100 ± 2.74 f | 88.83 ± 0.97 e | 78.62 ± 1.04 d | 60.32 ± 0.91 c | 27.76 ± 1.18 b | 9.86 ± 0.46 a |
| BT20 | 113.1 | 100 ± 9.86 c | 87.15 ± 5.51 c | 72.38 ± 4.73 b | 69.44 ± 7.89 b | 62.18 ± 4.16 b | 39.76 ± 9.75 a |
| BT474 | 93.63 | 100 ± 0.47 e | 92.98 ± 0.63 d | 91.94 ± 1.51 d | 77.65 ± 0.62 c | 23.95 ± 0.48 b | 1.51 ± 0.33 a |
| BT483 | 103.7 | 100 ± 4.78 c | 92.85 ± 1.58 c | 92.02 ± 0.94 c | 80.73 ± 3.44 b | 74.37 ± 4.19 b | 63.75 ± 7.59 a |
| BT549 | 71.08 | 100 ± 4.22 d | 73.59 ± 6.23 c | 61.09 ± 2.07 b | 59.93 ± 3.01 b | 53.59 ± 6.37 b | 21.90 ± 2.98 a |
| SK-BR-3 | 91.16 | 100 ± 3.75 d | 84.69 ± 3.09 d | 79.04 ± 3.27 c | 72.08 ± 10.11 c | 42.47 ± 3.36 b | 12.37 ± 0.22 a |
| ZR-75-30 | 88.55 | 100 ± 7.48 e | 62.04 ± 1.25 d | 54.80 ± 1.54 c | 53.02 ± 1.95 c | 28.72 ± 4.29 b | 8.66 ± 2.59 a |
| Hs578T | 94.04 | 100 ± 4.15 e | 73.88 ± 5.52 d | 70.56 ± 4.27 d | 63.17 ± 5.46 c | 40.01 ± 1.21 b | 2.11 ± 0.80 a |
| MCF-7 | 105.6 | 100 ± 1.04 d | 93.65 ± 1.04 c | 91.78 ± 0.43 c | 89.33 ± 1.25 c | 70.25 ± 5.69 b | 42.30 ± 0.38 a |
| Au565 | 109.2 | 100 ± 2.95 d | 72.38 ± 6.32 c | 67.98 ± 3.35 c | 57.67 ± 2.14 b | 51.28 ± 8.26 b | 2.04 ± 0.72 a |
| Bcap-37 | 78.92 | 100± 2.22 f | 89.54 ± 3.70 e | 80.12 ± 4.72 d | 64.51 ± 3.71 c | 47.27 ± 3.03 b | 1.42 ± 0.13 a |
| MCF-12A | ~246 | 100 ± 5.67 e | 90 ± 4.72 d | 79.49 ± 4.46 c | 76.34 ± 0.71bc | 68.71 ± 3.53 b | 53.88 ± 6.01 a |
| MCF-10A | ~280 | 100 ± 3.21 d | 89.68 ± 3.75 c | 88.82 ± 2.65 c | 86.07 ± 2.70 c | 72.92 ± 4.60 b | 57.90 ± 5.09 a |
| HS578BST | ~285 | 100 ± 3.42 c | 99.23 ± 0.75 c | 98.26 ± 1.81 c | 97.22 ± 0.18 c | 92.15 ± 3.87 b | 72.30 ± 3.27 a |
| Cell Lines | Molecular Classification | Years | Source and Disease | |||
|---|---|---|---|---|---|---|
| Name | Subtypes | ER | PR | HER2 | ||
| MDA-MB-453 | HER2 | + | 1976 | Female, 48 years, Caucasian, pericardial effusion, metastatic carcinoma | ||
| BT-549 | Basal-like | − | − | − | 1978 | Female, 72 years, Caucasian, breast, ductal carcinoma [59] |
| MDA-MB-361 | Luminal B | + | − | + | 1974 | Female, 40 years, Caucasian, brain, adenocarcinoma [60,61] |
| HS578T | Basal-like | − | − | − | 1977 | Female, 74 years, Caucasian, breast, carcinoma [62] |
| Bcap-37 | Luminal A | + | + | − | 1981 | Female, 48 years, Chinese, breast, medullary carcinoma [63] |
| HCC1395 | Basal-like | − | − | − | 1998 | Female, 43 years, Caucasian, breast/duct, ductal carcinoma [34,64] |
| BT-483 | Basal-like | − | − | − | 1978 | Female, 23 years, Caucasian, breast, ductal carcinoma [65,66] |
| SK-BR-3 | HER2 | − | − | + | 1970 | Female, 43 years, Caucasian, pleural effusion, adenocarcinoma [67] |
| BT-20 | Basal-like | − | − | − | 1958 | Female, 74 years, Caucasian, breast, carcinoma [68] |
| ZR-75-30 | Luminal A | + | + | − | 1978 | Female, 47 years, Black, ascites, ductal carcinoma [69] |
| HCC1806 | Basal-like | − | − | − | 1998 | Female, 60 years, Black, breast, squamous carcinoma [34,64] |
| MCF-7 | Luminal A | + | + | − | 1973 | Female, 69 years, Caucasian, pleural effusion, adenocarcinoma [59,60,70] |
| AU565 | HER2 | − | − | + | 1970 | Female, 43 years, Caucasian, malignant pleural effusion, malignant pleural effusion [67] |
| HCC1937 | Basal-like | − | − | − | 1998 | Female, 23 years, Caucasian, breast/duct, ductal carcinoma [64,71] |
| MDA-MB-231 | Basal-like | − | − | − | 1974 | Female, 51 years, Caucasian, pleural effusion, adenocarcinoma [60,61] |
| MDA-MB-157 | Basal-like | − | − | − | 1972 | Female, 44 years, Black, breast/medulla, medullary carcinoma [72] |
| BT-474 | Luminal B | + | + | + | 1978 | Female, 60 years, Caucasian, breast/duct, ductal carcinoma [59,65,66] |
| MCF-12A | Normal | 1992 | Female, 60 years, Caucasian, breast, fibrocystic, disease [66] | |||
| MCA-10A | Normal | 1984 | Female, 36 years, Caucasian, breast, fibrocystic, disease [37,66] | |||
| HS578BST | Normal | 1977 | Female, 74 years, Caucasian, breast, normal [62] | |||
| Primer Name (nAChR Subunits) | Sequence | Product Size |
|---|---|---|
| α3-F | AACGTGTCTGACCCAGTCATCAT | 139 bp |
| α3-R | AGGGGTTCCATTTCAGCTTGTAG | |
| α4-F | CGGCAGCCGCCGGATGAC | 201 bp |
| α4-R | ATGAAATTCGGCTCCTGGACCTA | |
| α5-F | ACTCCACCGGCAAACTACAA | 267 bp |
| α5-R | CAGGCGCTTGATTACAAATGA | |
| α6-F | GGCCTCTGGACAAGACAA | 414 bp |
| α6-R | AAGATTTTCCTGTGTTCCC | |
| α7-F | CCACCAACATTTGGCTGCAA | 218 bp |
| α7-R | TATGCCTGGAGGCAGGTACT | |
| α9-F | TGGCACGATGCCTATCTCAC | 171 bp |
| α9-R | TGATCAGCCCATCATACCGC | |
| α10-F | TGACCTCTTTGCCAACTACAC | 138 bp |
| α10-R | CACAGATACAGGGTCAGCAC | |
| β2-F | GGCATGTACGAGGTGTCCTT | 152 bp |
| β2-R | ACCAAGTCGATCTCTGTGCG | |
| β3-F | GGTCCGCCCTGTATTACATTC | 146 bp |
| β3-R | AGCGTCTAACTTGTGGTCTG | |
| β4-F | TCACAGCTCATCTCCATCAAGCT | 100 bp |
| β4-R | CCTGTTTCAGCCAGACATTGGT | |
| GAPDH-F | CAGCCTCAAGATCATCAGCA | 103 bp |
| GAPDH-R | TGTGGTCATGAGTCCTTCCA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Zhangsun, M.; Dong, S.; Liu, Y.; Qian, J.; Zhangsun, D.; Luo, S. Differential Expression of Nicotine Acetylcholine Receptors Associates with Human Breast Cancer and Mediates Antitumor Activity of αO-Conotoxin GeXIVA. Mar. Drugs 2020, 18, 61. https://doi.org/10.3390/md18010061
Sun Z, Zhangsun M, Dong S, Liu Y, Qian J, Zhangsun D, Luo S. Differential Expression of Nicotine Acetylcholine Receptors Associates with Human Breast Cancer and Mediates Antitumor Activity of αO-Conotoxin GeXIVA. Marine Drugs. 2020; 18(1):61. https://doi.org/10.3390/md18010061
Chicago/Turabian StyleSun, Zhihua, Manqi Zhangsun, Shuai Dong, Yiqiao Liu, Jiang Qian, Dongting Zhangsun, and Sulan Luo. 2020. "Differential Expression of Nicotine Acetylcholine Receptors Associates with Human Breast Cancer and Mediates Antitumor Activity of αO-Conotoxin GeXIVA" Marine Drugs 18, no. 1: 61. https://doi.org/10.3390/md18010061
APA StyleSun, Z., Zhangsun, M., Dong, S., Liu, Y., Qian, J., Zhangsun, D., & Luo, S. (2020). Differential Expression of Nicotine Acetylcholine Receptors Associates with Human Breast Cancer and Mediates Antitumor Activity of αO-Conotoxin GeXIVA. Marine Drugs, 18(1), 61. https://doi.org/10.3390/md18010061