Man-Specific, GalNAc/T/Tn-Specific and Neu5Ac-Specific Seaweed Lectins as Glycan Probes for the SARS-CoV-2 (COVID-19) Coronavirus




Abstract
:1. Introduction
2. Seaweed Lectins of Various Carbohydrate-Binding Specificities
2.1. Man-Specific Seaweed Lectins
- -
- The GNA (Galanthus nivalis agglutinin)-related family of lectins consists of protomers organized in a β-prism II or β-trefoil. The red alga Grateloupia chiangii lectin (GCL) and the green alga Boodlea coacta lectin (BCA) present this type of structural organization.
- -
- The jacalin-related family of lectins contains protomers organized in a β-prism I or β-barrel. Most of the mannose-specific lectins from red algae belong to this group of lectins, e.g., griffithsin and lectins from the genera Gracilaria, Eucheuma, Grateloupia, Kappaphycus, and Solieria.
- -
- The legume lectin-related family is made of protomers organized in a β-sandwich or jelly roll fold (two β-sheets). This structural scaffold occurs in a few lectins from the genera Hydropuntia (red algae), Nannochloropsis (yellow-green algae), Ostreococcus (green algae), and Porphyra (red algae).
2.1.1. Man-Specific Seaweed Lectins With a β-Prism II Structure
2.1.2. Man-Specific Seaweed Lectins with a β-Sandwich Structure
2.1.3. Man-Specific Seaweed Lectins with a β-Prism I Structure
2.2. Seaweed Lectins with GalNAc/T/Tn- and Neu5Ac-Specificity
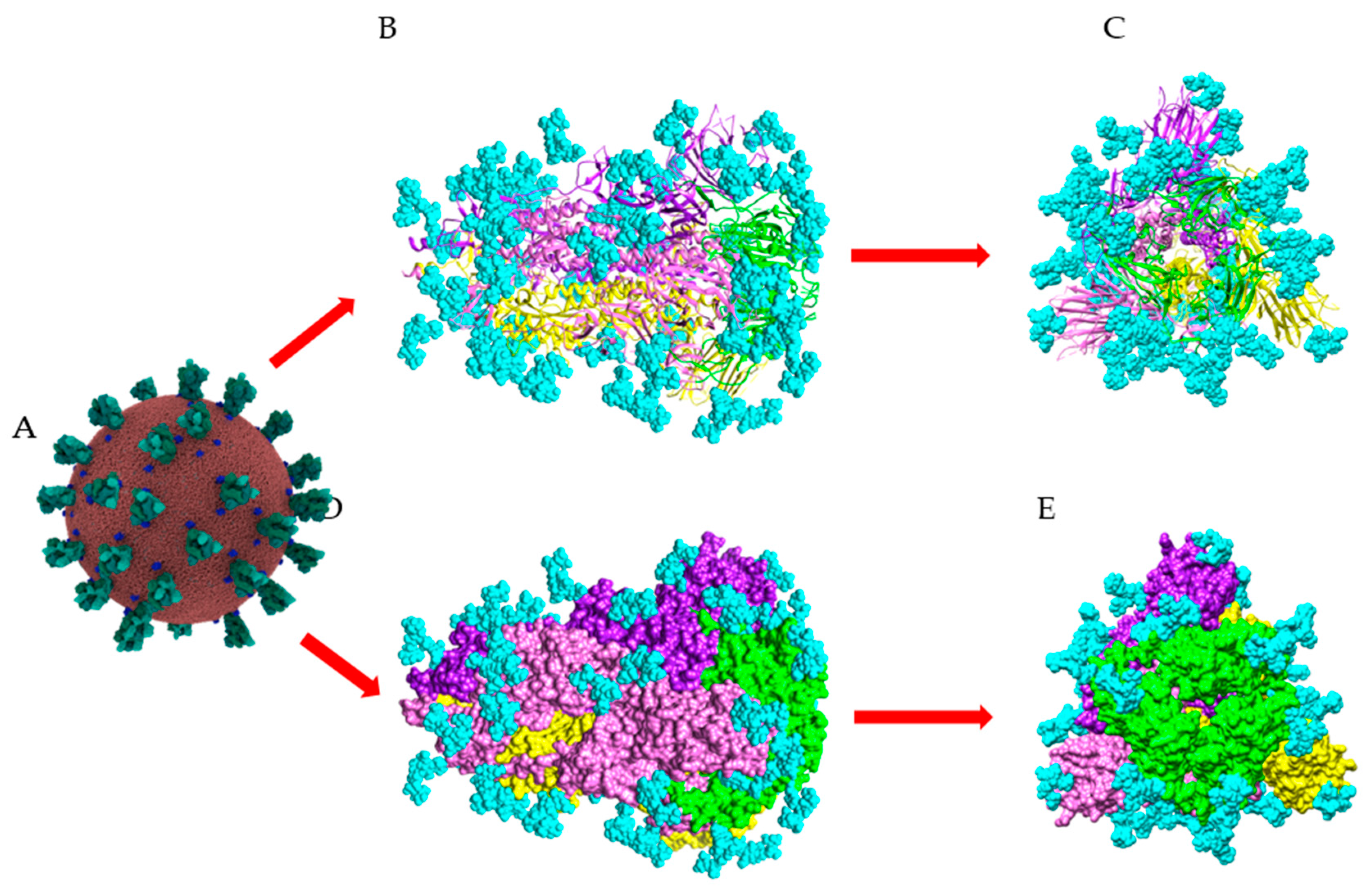
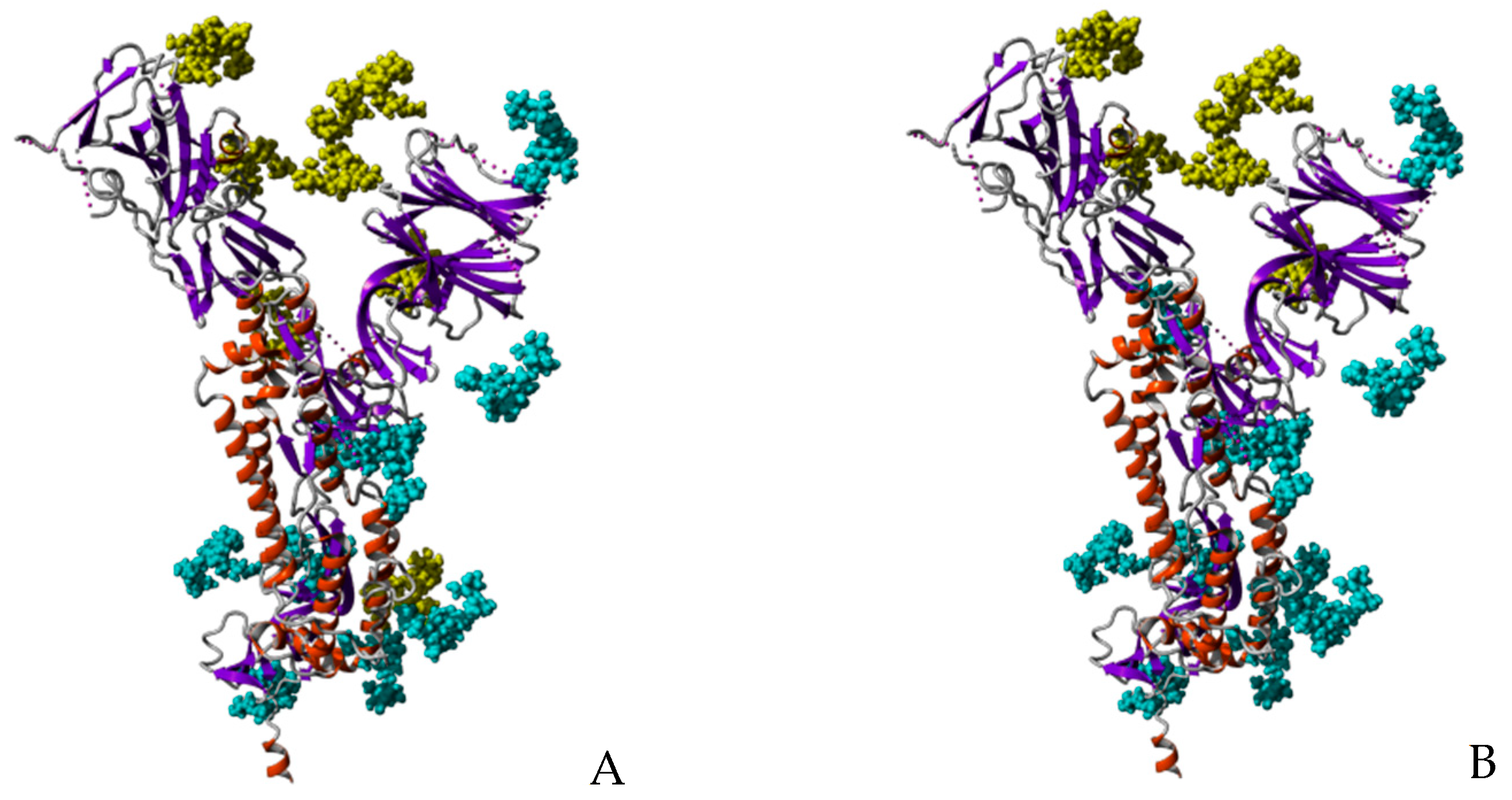
3. Structural Organization and Glycosylation of S Glycoproteins Forming the Spikes of SARS-CoV-2 Virus
- -
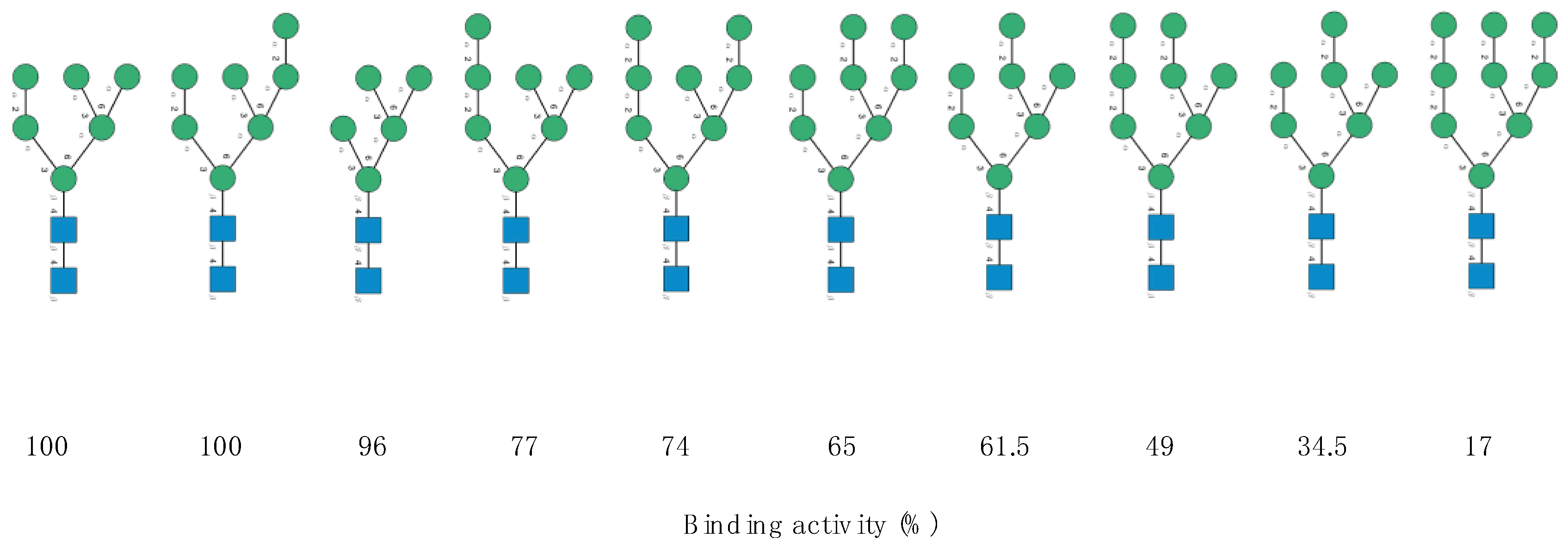
- N-glycosylation sites 149NKS, 165NCT, 282NGT, 657NNS, 709NNS, 801NFT, 1074NGT, 1098NGT and 1194NES (often not glycosylated), are almost exclusively occupied by often sialylated, bi-, tri- and tetra-antennary N-glycans of the complex type (Figure 10)
- -
- N-glycosylation sites 61NVT, 331NIT, 343NAT and 616NCT, are almost exclusively occupied by N-glycans of the high-mannose type (Figure 10)
- -
- The remaining N-glycosylation 74NGT, 122NAT, 234NIT and 717NFT, contain a mix of N-glycans of both types, complex N-glycans and high-mannose N-glycans (Figure 10)
- -
- Both O-glycosylated sites T323, S325, harbor core-1 mucin type O-glycans: GalNAc, T-antigen GalNAcGal, sialylated T-antigen GalNAcGalNeuAc2, and core-2 sialylated O-glycans GalNAcGalNeuAc(GlcNAcGal), and GalNAcGalNeuAc(GlcNAcGalNeuAc) (Figure 10)
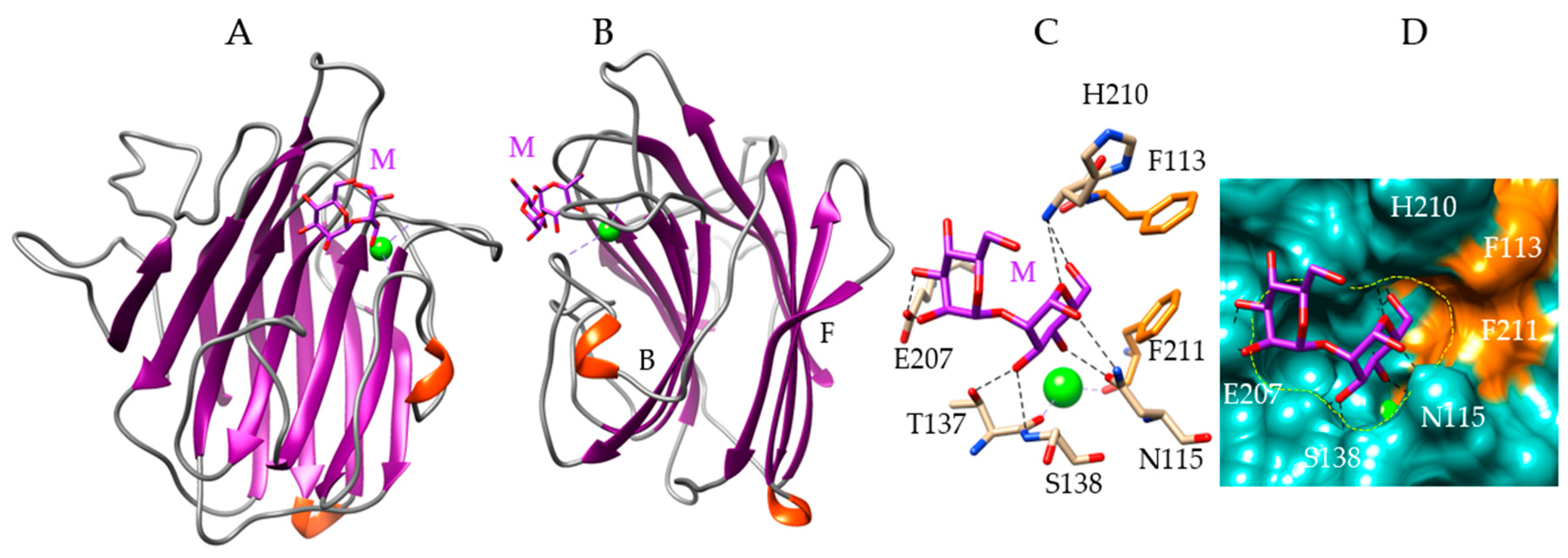
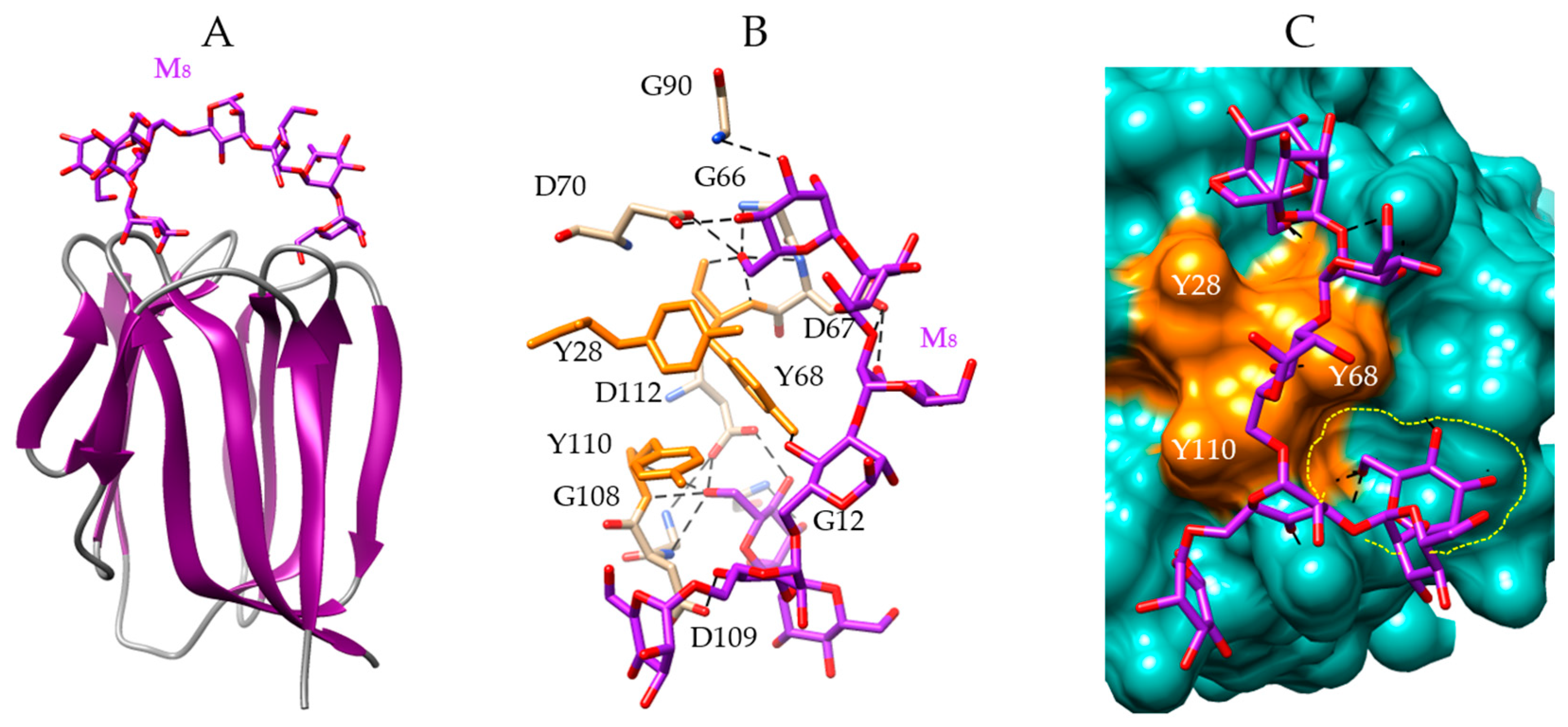
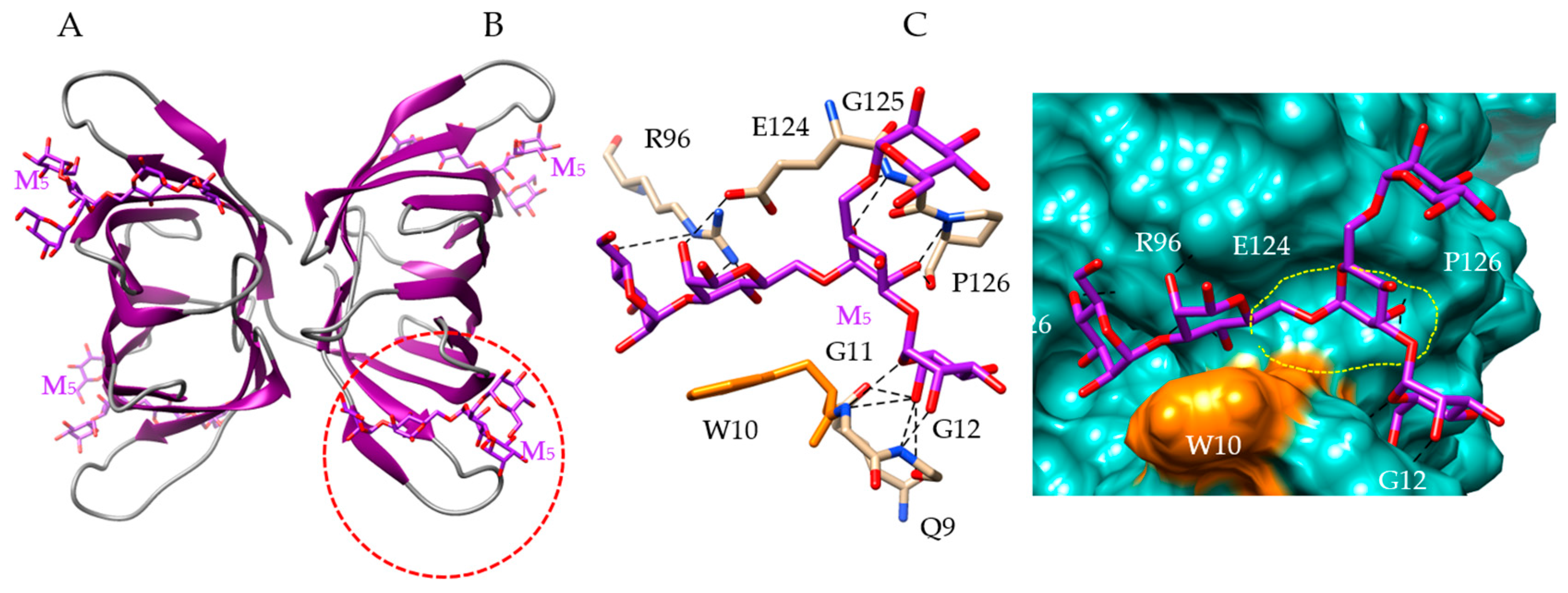
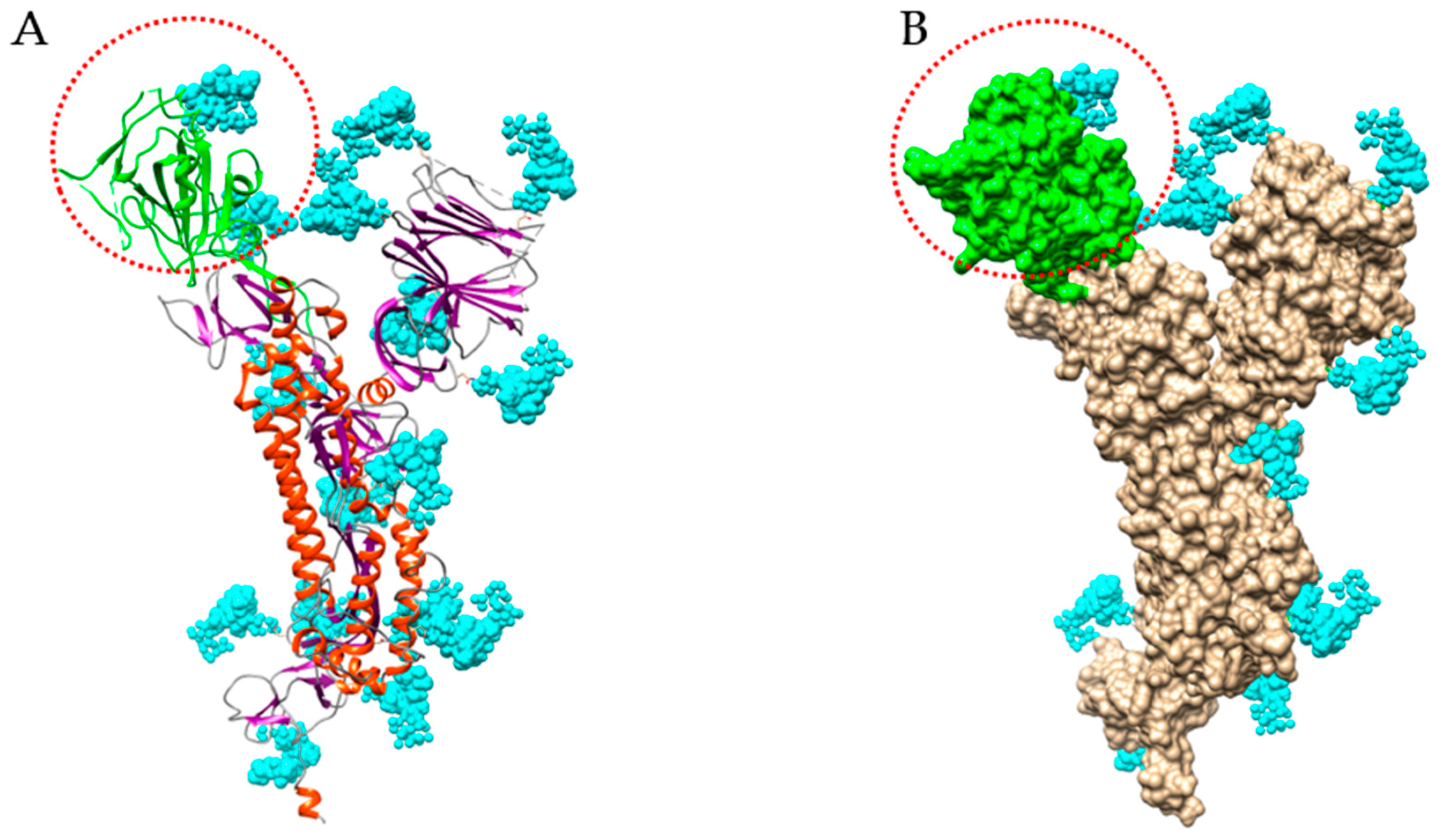
4. Man-Specific Seaweed Lectins Interact with N-glycans Decorating the S-Glycoprotein from SARS-CoV-2
5. Interaction of Other Seaweed Lectins with Different Specificities with the S-Glycoprotein from SARS-CoV-2
6. Bioinformatics
7. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AOL1 | Aglaothamnion oosumiense lectin |
| APA | Allium porrum agglutinin |
| ASL | Agardhiella subulata lectin |
| BCA | Boodlea coacta lectin |
| BOA | Burkholderia oklahomensis agglutinin |
| BPL2 | Lectin 2 of Bryopsis plumosa |
| BU14 | Nannochloropsis gaditana lectin |
| CBS | Carbohydrate-binding site |
| CFA | Carpopeltis flabellata agglutinin |
| CFL | Codium fragile lectin |
| CV-N | Cyanovirin-N (Nostoc ellipsosporum) |
| EEA | Eucheuma amakusanensis lectin |
| ECA | Eucheuma cottonii lectin |
| EDA | Eucheuma denticulatum lectin |
| EPL | Enteromorpha prolifera lectin |
| ESA | Eucheuma serra lectin |
| FRIL | Flt3 receptor interacting lectin from hyacinth bean (Lablab purpureus) |
| Fuc | Fucose |
| Gal | Galactose |
| GalNAc | N-acetylgalactosamine |
| GBPL | Gracilaria bursa-pastoris lectin |
| GCL | Grateloupia chiangii lectin |
| GlcNAc | N-acetylglucosamine |
| GNA | Galanthus nivalis (snowdrop) agglutinin |
| GPT | Glycoprotein trimer |
| GRFT | Griffithsin |
| GTL | Gracilaria tikvahiae lectin |
| HFA | Hydropuntia (Gracilaria) fisheri agglutinin |
| HIV-1 | Human Immunodeficiency Virus |
| HRL40 | Halimeda renschii lectin |
| KAA-2 | Lectin 2 of Kappaphycus alvarezii |
| KSA-2 | Lectin 2 of Kappaphycus striatum |
| Man | Mannose |
| MEL | Meristiella echinocarpa lectin |
| MPL | Meristotheca papulosa agglutinin |
| MVL | Microcystis viridis lectin |
| MVN | Microvirin (Microcystis aeruginisa) |
| Neu5Ac | N-acetylneuraminic acid (sialic acid) |
| OAA | Oscillatoria agardhii lectin |
| OtL | Ostreococcus tauri lectin |
| PDB | Protein Data Bank |
| PPL | Palmaria palmata lectin |
| PUL | Porphyra umbilicalis lectin |
| RBD | Receptor binding domain |
| SARS-CoV-2 | Severe acute respiratory syndrome-coronavirus |
| Sia | Sialic acid |
| ScL | Solieria chordalis lectin |
| SfL | Solieria filiformis lectin |
| SrL | Solieria robusta lectin |
| SVN | Scytovirin (Scytonema varium) |
| T/Tn | T and Tn antigens (Thomsen Friedenreich antigens) |
References
- Boyd, W.C.; Almodóvar, L.R.; Boyd, L.G. Agglutinins in Marine Algae for Human Erythrocytes. Transfusion 1966, 6, 82–83. [Google Scholar] [CrossRef]
- Blunden, G.; Rogers, D.J.; Farnham, W.F. Survey of British Seaweeds for Hæmagglutinins. Lloydia 1975, 36, 162–168. [Google Scholar]
- Hori, K.; Ikegami, S.; Miyazawa, K.; Ito, K. Mitogenic and Antineoplastic Isoagglutinins from the Red Alga Solieria robusta. Phytochemistry 1988, 27, 2063–2067. [Google Scholar] [CrossRef]
- Sugahara, T.; Ohama, Y.; Fukuda, A.; Hayashi, M.; Kawakubo, A.; Kato, K. The Cytotoxic Effect of Eucheuma serrata Agglutinin (ESA) on Cancer Cells and Its Application to Molecular Probe for Drug Delivery System Using Lipid Vesicles. Cytotechnology 2001, 36, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Sugahara, T.; Ueno, M.; Fukuta, Y.; Ochi, Y.; Akiyama, K.; Miyazaki, T.; Masuda, S.; Kawakubo, A.; Kato, K. The Anti-Tumor Effect of Euchema serra Agglutinin on Colon Cancer Cells In Vitro and In Vivo. Anticancer Drugs 2006, 17, 943–947. [Google Scholar] [CrossRef]
- Hayashi, K.; Walde, P.; Miyazaki, T.; Sakayama, K.; Nakamura, A.; Kameda, K.; Masuda, S.; Umakoshi, H.; Kato, K. Active Targeting to Osteosarcoma Cells and Apoptotic Cell Death Induction by the Novel Lectin Eucheuma serra Agglutinin Isolated from a Marine Red Alga. J. Drug Deliv. 2012, 2012, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chaves, R.P.; Da Silva, S.R.; Neto, L.G.N.; Carneiro, R.F.; Da Silva, A.L.C.; Sampaio, A.H.; De Sousa, B.L.; Cabral, M.G.; Videira, P.A.; Teixeira, E.H.; et al. Structural Characterization of Two Isolectins From the Marine Red Alga Solieria iliformis (Kützing) P.W. Gabrielson and Their Anticancer Effect on MCF-7 Breast Cancer Cells. Int. J. Biol. Macromol. 2018, 107, 1320–1329. [Google Scholar] [CrossRef]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C.; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.; McMahon, J.B.; et al. Isolation and Characterization of Griffithsin, a Novel HIV-Inactivating Protein, from the Red Alga Griffithsia sp. J. Biol. Chem. 2004, 280, 9345–9353. [Google Scholar] [CrossRef] [Green Version]
- Ziółkowska, N.E.; Wlodawer, A. Structural Studies of Algal Lectins with Anti-HIV Activity. Acta Biochim. Pol. 2006, 53, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High Mannose-Specific Lectin (KAA-2) From the Red Alga Kappaphycus alvarezii Potently Inhibits Influenza Virus Infection in a Strain-Independent Manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High Mannose-binding Lectin with Preference for the Cluster of α1–2-Mannose from the Green Alga Boodlea coacta Is a Potent Entry Inhibitor of HIV-1 and Influenza Viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Morimoto, K.; Kubo, T.; Sakaguchi, T.; Nishizono, A.; Hirayama, M.; Hori, K. Entry Inhibition of Influenza Viruses with High Mannose Binding Lectin ESA-2 from the Red Alga Eucheuma serra through the Recognition of Viral Hemagglutinin. Mar. Drugs 2015, 13, 3454–3465. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-Mannose Specific Lectin and Its Recombinants from Carrageenophyte Kappaphycus Alvarezii Represent a Potent Anti-HIV Activity Through High-Affinity Binding to the Viral Envelope Glycoprotein gp120. Mar. Biotechnol. 2016, 18, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.-J.; Han, J.-W.; Jeon, H.; Cho, K.; Kim, J.-H.; Lee, D.-S.; Han, J.W. Characterization of a Novel Mannose-Binding Lectin with Antiviral Activities from Red Alga, Grateloupia chiangii. Biomolecules 2020, 10, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Okuyama, S.; Hori, K. Primary Structure and Carbohydrate Binding Specificity of a Potent Anti-HIV Lectin Isolated from the Filamentous Cyanobacterium, Oscillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of Cyanovirin-N, a Novel Human Immunodeficiency Virus-Inactivating Protein That Binds Viral Surface Envelope Glycoprotein gp120: Potential Applications to Microbicide Development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef] [Green Version]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple Antiviral Activities of Cynovirin-N: Blocking of Human Immunodeficiency Virus Type 1 gp120 Interaction with CD4 and Coreceptor and Inhibition of Diverse Enveloped Viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef]
- Barrientos, L.G.; O’Keefe, B.R.; Bray, M.; Sanchez, A.; Gronenborn, A.M.; Boyd, M.R. Cyanovirin-N Binds to the Viral Surface Glycoprotein, GP1,2 and Inhibits Infectivity of Ebola Virus. Antivir. Res. 2003, 58, 47–56. [Google Scholar] [CrossRef]
- Helle, F.; Wychowski, C.; Vu-Dac, N.; Gustafson, K.R.; Voisset, C.; Dubuisson, J. Cyanovirin-N Inhibits Hepatitis C Virus Entry by Binding to Envelope Protein Glycans. J. Biol. Chem. 2006, 281, 25177–25183. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Smee, D.F.; Turpin, J.A.; Saucedo, C.J.; Gustafson, K.R.; Mori, T.; Blakeslee, D.; Buckheit, R.; Boyd, M.R. Potent Anti-Influenza Activity of Cyanovirin-N and Interactions with Viral Hemagglutinin. Antimicrob. Agents Chemother. 2003, 47, 2518–2525. [Google Scholar] [CrossRef] [Green Version]
- Huskens, D.; Férir, G.; Vermeire, K.; Kehr, J.-C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a Novel α(1,2)-Mannose-specific Lectin Isolated from Microcystis aeruginosa, Has Anti-HIV-1 Activity Comparable with That of Cyanovirin-N but a Much Higher Safety Profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bewley, C.A.; Cai, M.; Ray, S.; Ghirlando, R.; Yamaguchi, M.; Muramoto, K. New Carbohydrate Specificity and HIV-1 Fusion Blocking Activity of the Cyanobacterial Protein MVL: NMR, ITC and Sedimentation Equilibrium Studies. J. Mol. Biol. 2004, 339, 901–914. [Google Scholar] [CrossRef]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.L.; Gardella, R.S.; Sowder, R.C.; Turpin, J.; Watson, K.; Buckheit, R.W.; et al. A Potent Novel Anti-HIV Protein from the Cultured Cyanobacteriumb Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef]
- Shajahan, A.; Supekar, N.T.; Gleinich, A.S.; Azadi, P. Deducing the N- and O- Glycosylation Profile of the Spike Protein of Novel Coronavirus SARS-CoV-2. Glycobiology 2020, cwaa042. [Google Scholar] [CrossRef]
- Sanda, M.; Morrison, L.; Goldman, R. N and O Glycosylation of the SARS-CoV-2 Spike Protein. bioRxiv 2020. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Herrera, N.G.; Morano, N.C.; Celikgil, A.; Georgiev, G.I.; Malonis, R.J.; Lee, J.H.; Tong, K.; Vergnolle, O.; Massimi, A.B.; Yen, L.Y.; et al. Characterization of the SARS-CoV-2 S Protein: Biophysical, Biochemical, Structural, and Antigenic Analysis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Xiong, X.; The CITIID-NIHR COVID-19 BioResource Collaboration; Qu, K.; Ciazynska, K.A.; Hosmillo, M.; Carter, A.P.; Ebrahimi, S.; Ke, Z.; Scheres, S.H.W.; Bergamaschi, L.; et al. A Thermostable, Closed SARS-CoV-2 Spike Protein Trimer. Nat. Struct. Mol. Biol. 2020, 27, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Tsybovsky, Y.; Olia, A.S.; Gorman, J.; Rapp, M.A.; Cerutti, G.; Katsamba, P.S.; Nazzari, A.; Schon, A.; Wang, P.D.; et al. A pH-Dependent Switch Mediates Conformational Masking of SARS-CoV-2 Spike. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e9. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- Li, M.-Y.; Li, L.; Zhang, Y.; Wang, X.-S. Expression of the SARS-CoV-2 Cell Receptor Gene ACE2 in a Wide Variety of Human Tissues. Infect. Dis. Poverty 2020, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Maurya, V.K.; Prasad, A.K.; Bhatt, M.L.B.; Saxena, S.K. Structural, Glycosylation and Antigenic Variation Between 2019 Novel Coronavirus (2019-nCov) and SARS Coronavirus (SARS-CoV). VirusDisease 2020, 31, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Hori, K.; Matsuda, H.; Miyazawa, K.; Ito, K. A Mitogenic Agglutinin from the Red Alga Carpopeltis flabellata. Phytochemistry 1987, 26, 1335–1338. [Google Scholar] [CrossRef]
- Kawakubo, A.; Makino, H.; Ohnishi, J.-I.; Hirohara, H.; Hori, K. Occurrence of Highly Yielded Lectins Homologous Within the Genus Eucheuma. J. Apppl. Phycol. 1999, 11, 149–156. [Google Scholar] [CrossRef]
- Hung, L.D.; Sato, Y.; Hori, K. High- Mannose N-Glycan-Specific Lectin from the Red Alga Kappaphycus striatum (Carrageenophyte). Phytochemistry 2011, 72, 855–861. [Google Scholar] [CrossRef]
- Kawakubo, A.; Makino, H.; Ohnishi, J.-I.; Hirohara, H.; Hori, K. The Marine Red Alga Eucheuma serra J. Agardh, a High Yielding Source of Two Isolectins. J. Apppl. Phycol. 1997, 9, 331–338. [Google Scholar]
- Hori, K.; Sato, Y.; Ito, K.; Fujiwara, Y.; Iwamoto, Y.; Makino, H.; Kawakubo, A. Strict Specificity for High-Mannose Type N-Glycans and Primary Structure of a Red Alga Eucheuma serra Lectin. Glycobiology 2007, 17, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, R.K.; Hori, K.; Miyazawa, K.; Ito, K. Isolation and Characterisation of a New Haemagglutinin From the Red Alga Gracilaria bursa-pastoris. Experientia 1990, 46, 975–977. [Google Scholar] [CrossRef] [PubMed]
- Chaves, R.P.; Da Silva, S.R.; Da Silva, J.P.F.A.; Carneiro, R.F.; De Sousa, B.L.; Abreu, J.O.; De Carvalho, F.C.T.; Rocha, C.R.C.; Farias, W.R.L.; De Sousa, O.V.; et al. Meristiella echinocarpa Lectin (MEL): A New Member of the OAAH-Lectin Family. J. Appl. Phycol. 2018, 30, 2629–2638. [Google Scholar] [CrossRef]
- Suttisrisung, S.; Senapin, S.; Withyachumnarnkul, B.; Wongprasert, K. Identification and Characterization of a Novel Legume-Like Lectin cDNA Sequence from the Red Marine Algae Gracilaria fisheri. J. Biosci. 2011, 36, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Van Holle, S.; Damme, E.J. Messages from the Past: New Insights in Plant Lectin Evolution. Front. Plant Sci. 2019, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Carpinelli, E.C.; Telatin, A.; Vitulo, N.; Forcato, C.; D’Angelo, M.; Schiavon, R.; Vezzi, A.; Giacometti, G.M.; Morosinotto, T.; Valle, G. Chromosome Scale Genome Assembly and Transcriptome Profiling of Nannochloropsis gaditana in Nitrogen Depletion. Mol. Plant 2014, 7, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Hori, K.; Miyazawa, K.; Ito, K. Isolation and Characterization of Glycoconjugate-Specific Isoagglutinins from a Marine Green Alga Boodlea coacta Murray et De Toni. Bot. Mar. 1986, 29, 323–328. [Google Scholar] [CrossRef]
- Hwang, H.-J.; Han, J.-W.; Jeon, H.; Han, J.W. Induction of Recombinant Lectin Expression by an Artificially Constructed Tandem Repeat Structure: A Case Study Using Bryopsis plumosa Mannose-Binding Lectin. Biomolecules 2018, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, A.L.; Sanz, L.; Sánchez, E.I.; Wolfenstein-Todel, C.; Calvete, J.J. Isolation of Two Novel Mannan-and L-Fucose-Binding Lectins from the Green Alga Enteromorpha prolifera: Biochemical Characterization of EPL-2. Arch. Biochem. Biophys. 2003, 415, 245–250. [Google Scholar] [CrossRef]
- Derelle, E.; Ferraz, C.; Rombauts, S.; Rouzé, P.; Worden, A.Z.; Robbens, S.; Partensky, F.; Degroeve, S.; Echeynié, S.; Cooke, R.; et al. Genome Analysis of the Smallest Free-Living Eukaryote Ostreococcus tauri Unveils Many Unique Features. Proc. Natl. Acad. Sci. USA 2006, 103, 11647–11652. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, E.J.M.; Rougé, P.; Peumans, W. Plant Lectins. In Protein Interactions: Plant Lectins; Kamerling, J.P., Boons, G.J., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.I., Eds.; Elsevier: New York, NY, USA, 2007; pp. 564–599. [Google Scholar]
- Hardman, K.D.; Ainsworth, C.F. Structure of Concanavalin A at 2.4-Ang Resolution. Biochemistry 1972, 11, 4910–4919. [Google Scholar] [CrossRef]
- Einspahr, H.; Parks, E.H.; Suguna, K.; Subramanian, E.; Suddath, F.L. The Crystal Structure of Pea Lectin at 3.0-A Resolution. J. Biol. Chem. 1986, 261, 16518–16527. [Google Scholar] [PubMed]
- Loris, R.; Van Overberge, D.; Dao-Thi, M.-H.; Poortmans, F.; Maene, N.; Wyns, L. Structural Analysis of Two Crystal Forms of Lentil Lectin at 1.8 A Resolution. Proteins 1994, 20, 330–346. [Google Scholar] [CrossRef] [PubMed]
- Velloso, L.M.; Svensson, K.; Schneider, G.; Pettersson, R.F.; Lindqvist, Y. Crystal Structure of the Carbohydrate Recognition Domain of p58/Ergic-53, a Protein Involved in Glycoprotein Export from the Endoplasmic Reticulum. J. Biol. Chem. 2002, 277, 15979–15984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, T.; Cowieson, N.P.; Hakamata, W.; Ideo, H.; Fukushima, K.; Kurihara, M.; Kato, R.; Yamashita, K.; Wakatsuki, S. Structural Basis for Recognition of High Mannose Type Glycoproteins by Mammalian Transport Lectin VIP36. J. Biol. Chem. 2007, 282, 28246–28255. [Google Scholar] [CrossRef] [Green Version]
- Han, J.W.; Klochkova, T.A.; Shim, J.B.; Yoon, K.; Kim, G.H. Isolation and Characterization of a Sex-Specific Lectin in a Marine Red Alga, Aglaothamnion oosumiense Itono. Appl. Environ. Microbiol. 2012, 78, 7283–7289. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.M.; Song, S.-C.; Hwang, P.-Y.; Wu, J.H.; Chang, K.S.S. Binding Studies on the Combining Site of a GalNAcalpha1-Specific Lectin with Thomsen-Friedenreich Activity Prepared from Green Marine Algae Codium fragile Subspecies Tomentosoides. Eur. J. Biochem. 1995, 233, 145–151. [Google Scholar]
- Wu, A.M.; Song, S.-C.; Chang, S.-C.; Kabat, E. Further Characterization of the Binding Properties of a GalNAc Specific Lectin from Codium fragile Subspecies tomentosoides. Glycobiology 1997, 7, 1061–1066. [Google Scholar] [CrossRef] [Green Version]
- Chiles, T.C.; Bird, K.T. Gracilaria Tikvahiae Agglutinin. Partial Purification and Preliminary Characterization of Its Carbohydrate Specificity. Carbohydr. Res. 1990, 207, 319–326. [Google Scholar] [CrossRef]
- Kamiya, H.; Ogata, K.; Hori, K.; Hori, K.O.K. Isolation and Characterization of a New Agglutinin in the Red Alga Palmaria palmata (L.) O. Kuntze. Bot. Mar. 1982, 25, 537–540. [Google Scholar] [CrossRef]
- Rogers, D.J.; Topliss, J.A. Purification and Characterization of an Anti-Sialic Acid Agglutinin from the Red Alga Solieria chordalis (C. Ag.). Bot. Mar. 1983, 26, 301–306. [Google Scholar] [CrossRef]
- Criado, M.; Ferreirós, C. Selective Interaction of a Fucus vesiculosus Lectin-Like Mucopolysaccharide with Several Candida Species. Ann. Inst. Pasteur Microbiol. 1983, 134, 149–154. [Google Scholar] [CrossRef]
- Ritchie, G.; Harvey, D.J.; Feldmann, F.; Stroeher, U.; Feldmann, H.; Royle, L.; Dwek, R.A.; Rudd, P.M. Identification of N- Linked Carbohydrates from Severe Acute Respiratory Syndrome (SARS) Spike Glycoprotein. Virology 2010, 399, 257–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A Highly Conserved Cryptic Epitope in the Receptor Binding Domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Liu, H.; Wu, N.C.; Lee, C.-C.D.; Zhu, X.; Zhao, F.; Huang, D.; Yu, W.; Hua, Y.; Tien, H.; et al. Structural Basis of a Public Antibody Response to SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in Humanized Mice and Convalescent Humans Yield a SARS-CoV-2 Antibody Cocktail. Science 2020, eabd0827. [Google Scholar] [CrossRef]
- Hurlburt, N.K.; Wan, Y.-H.; Stuart, A.B.; Feng, J.; McGuire, A.T.; Stamatatos, L.; Pancera, M. Structural Basis for Potent Neutralization of SARS-Cov-2 and Role of Antibody Affinity Maturation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhou, D.; Duyvesteyn, H.M.E.; Chen, C.-P.; Huang, C.-G.; Chen, T.-H.; Shih, S.-R.; Lin, Y.-C.; Cheng, C.-Y.; Cheng, S.-H.; Huang, Y.-C.; et al. Structural Basis for the Neutralization of SARS-Cov-2 by an Antibody from a Convalescent Patient. Nat. Struct. Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human Neutralizing Antibodies Elicited by SARS-CoV-2 Infection. Nat. Cell Biol. 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding to Its Receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Wu, L.; Gao, G.; et al. A Human Neutralizing Antibody Targets the Receptor-Binding Site of SARS-CoV-2. Nat. Cell Biol. 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Wu, N.C.; Yuan, M.; Liu, H.; Lee, C.C.D.; Zhu, X.; Bangaru, S.; Torres, J.L.; Caniels, T.G.; Brouwer, P.J.M.; van Gils, M.J.; et al. An alternative binding mode of IGHV3-53 antibodies to the SARS-CoV-2 receptor binding domain. Cell Rep. 2020, 33, 108274. [Google Scholar] [CrossRef]
- Pinto, D.; Park, Y.-J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Structural and Functional Analysis of a Potent Sarbecovirus Neutralizing Antibody. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Henderson, R.; Edwards, R.J.; Mansouri, K.; Janowska, K.; Stalls, V.; Gobeil, S.; Kopp, M.; Li, D.; Parks, R.; Hsu, A.L.; et al. Controlling the SARS-CoV-2 Spike Glycoprotein Conformation. Nat. Struct. Mol. Biol. 2020, 1–9. [Google Scholar] [CrossRef]
- McCallum, M.; Walls, A.C.; Bowen, J.E.; Corti, D.; Veesler, D. Structure-Guided Covalent Stabilization of Coronavirus Spike Glycoprotein Trimers in the Closed Conformation. Nat. Struct. Mol. Biol. 2020, 27, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.O.; West, A.P.; Huey-Tubman, K.E.; Hoffmann, M.A.; Sharaf, N.G.; Hoffman, P.R.; Koranda, N.; Gristick, H.B.; Gaebler, C.; Muecksch, F.; et al. Structures of Human Antibodies Bound to SARS-CoV-2 Spike Reveal Common Epitopes and Recurrent Features of Antibodies. Cell 2020, 182, 828–842.e16. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.-W.; Sahi, V.; Figueroa, A.; et al. Potent Neutralizing Antibodies Against Multiple Epitopes on SARS-CoV-2 Spike. Nat. Cell Biol. 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-L.; Goldsmith, J.A.; Schaub, J.M.; DiVenere, A.M.; Kuo, H.-C.; Javanmardi, K.; Le, K.C.; Wrapp, D.; Lee, A.G.; Liu, Y.; et al. Structure-Based Design of Prefusion-Stabilized SARS-CoV-2 Spikes. Science 2020, 369, 1501–1505. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A Neutralizing Human Antibody Binds to the N-Terminal Domain of the Spike Protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Pascal, K.E.; Tompkins-Tinch, C.; Nyalile, T.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C.; Veinotte, K.; et al. SARS-CoV-2 Spike Protein Variant D614G Increases Infectivity and Retains Sensitivity to Antibodies That Target the Receptor Binding Domain. bioRxiv 2020. [Google Scholar] [CrossRef]
- Huo, J.; Zhao, Y.; Ren, J.; Zhou, D.; Duyvesteyn, H.M.E.; Ginn, H.M.; Carrique, L.; Malinauskas, T.; Ruza, R.R.; Shah, P.N.; et al. Neutralization of SARS-CoV-2 by Destruction of the Prefusion Spike. Cell Host Microbe 2020, 30351–30356. [Google Scholar] [CrossRef]
- Melero, R.; Sorzano, C.O.S.; Foster, B.; Vilas, J.-L.; Martínez, M.; Marabini, R.; Ramírez-Aportela, E.; Sanchez-Garcia, R.; Herreros, D.; Del Caño, L.; et al. Continuous Flexibility Analysis of SARS-Cov-2 Spike Prefusion Structures. IUCrJ 2020, 7. [Google Scholar] [CrossRef]
- Ke, Z.; Oton, J.; Qu, K.; Cortese, M.; Zila, V.; McKeane, L.; Nakane, T.; Zivanov, J.; Neufeldt, C.J.; Cerikan, B.; et al. Structures and Distributions of SARS-Cov-2 Spike Proteins on Intact Virions. Nat. Cell Biol. 2020, 1–7. [Google Scholar] [CrossRef]
- Cao, Y.; Su, B.; Guo, X.; Sun, W.; Deng, Y.; Bao, L.; Zhu, Q.; Zhang, X.; Zheng, Y.; Geng, C.; et al. Potent Neutralizing Antibodies Against Sars-Cov-2 Identified by High-Throughput Single-Cell Sequencing of Covalescent Patients’ B Cells. Cell 2020, 182, 73–84. [Google Scholar] [PubMed]
- Liu, Y.-M.; Mahmud, S.-A.; Chen, X.; Chen, T.-H.; Liao, K.-S.; Lo, J.M.; Wu, Y.-M.; Ho, M.-C.; Wu, C.-Y.; Wong, C.-H.; et al. A Carbohydrate-Binding Protein from the Edible Lablab Beans Effectively Blocks the Infections of Influenza Viruses and SARS-CoV-2. Cell Rep. 2020, 32, 108016. [Google Scholar] [CrossRef] [PubMed]
- Bangaru, S.; Ozorowski, G.; Turner, H.L.; Antanasijevic, A.; Huang, D.; Wang, X.; Torres, J.L.; Diedrich, J.K.; Tian, J.-H.; Portnoff, A.D.; et al. Structural Analysis of Full-Length SARS-Cov-2 Spike Protein from an Advanced Vaccine Candidate. Science 2020, eabe1502. [Google Scholar] [CrossRef]
- Ziółkowska, N.E.; O’Keefe, B.R.; Mori, T.; Zhu, C.; Giomarelli, B.; Vojdani, F.; Palmer, K.E.; McMahon, J.B.; Wlodawer, A. Domain-Swapped Structure of the Potent Antiviral Protein Griffithsin and Its Mode of Carbohydrate Binding. Structure 2006, 14, 1127–1135. [Google Scholar] [CrossRef] [Green Version]
- Ziółkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; McMahon, J.B.; Palmer, K.E.; Dwek, R.A.; Wormald, M.R.; Wlodawer, A. Crystallographic, Thermodynamic, and Molecular Modeling Studies of the Mode of Binding of Oligosaccharides to the Potent Antiviral Protein Griffithsin. Proteins 2007, 67, 661–670. [Google Scholar] [CrossRef]
- Moulaei, T.; Shenoy, S.R.; Giomarelli, B.; Thomas, C.; McMahon, J.B.; Dauter, Z.; O’Keefe, B.R.; Wlodawer, A. Monomerization of Viral Entry Inhibitor Griffithsin Elucidates the Relationship between Multivalent Binding to Carbohydrates and Anti-HIV Activity. Structure 2010, 18, 1104–1115. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Whitley, M.J.; Furey, W.; Kollipara, S.; Gronenborn, A.M. Burkholderia Oklahomensisagglutinin Is a Canonical Two-Domain OAA-Family Lectin: Structures, Carbohydrate Binding and Anti-HIV Activity. FEBS J. 2013, 280, 2056–2067. [Google Scholar] [CrossRef] [Green Version]
- Koharudin, L.M.; Gronenborn, A.M. Structural Basis of the Anti-HIV Activity of the Cyanobacterial Oscillatoria agardhii Agglutinin. Structure 2011, 19, 1170–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the Precision of Comparative Models with Yasara Nova—A Self-Parametrizing Force Field. Proteins 2002, 47, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemistry of Protein Structures. J. Appl. Crystallogr. 1993, 126, 283–291. [Google Scholar] [CrossRef]
- Melo, F.; Feytmans, E. Assessing Protein Structures with a Non-Local Atomic Interaction Energy. J. Mol. Biol. 1998, 277, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the Estimation of the Absolute Quality of Individual Protein Structure Models. Bioinformatics 2010, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL Workspace: A Web-Based Environment for Protein Structure Homology Modelling. Bioinformatics 2005, 22, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, A Protein-Small Molecule Docking Web Service Based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.S.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-Spectrum In Vitro Activity and In Vivo Efficacy of the Antiviral Protein Griffithsin against Emerging Viruses of the Family Coronaviridae. J. Virol. 2010, 84, 5456. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Séron, K.; Labitt, R.N.; Danneels, A.; Palmer, K.E.; Whittaker, G.R.; Dubuisson, J.; Belouzard, S. Middle East Respiratory Syndrome Coronavirus Infection Is Inhibited by Griffithsin. Antivir. Res. 2016, 133, 1–8. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; Van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; Van Ranst, M. Plant Lectins Are Potent Inhibitors of Coronaviruses by Interfering with Two Targets in the Viral Replication Cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Mani, J.S.; Johnson, J.B.; Steel, J.C.; Broszczak, D.A.; Neilsen, P.M.; Walsh, K.B.; Naiker, M. Natural Product-Derived Phytochemicals as Potential Agents Against Coronaviruses: A Review. Virus Res. 2020, 284, 197989. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The Potential of Algal Biotechnology to Produce Antiviral Compounds and Biopharmaceuticals. Molecules 2020, 25, 4049. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; Suhail, M.; Kamal, M.A.; Ahmad, F.; Azhar, E.I. Emergence of Human Pathogenic Coronaviruses: Lectins as Antivirals for SARS-CoV-2. Curr. Pharm. Des. 2020, 26, 1–10. [Google Scholar] [CrossRef]
- Gupta, R.K.; Apte, G.R.; Lokhande, K.B.; Mishra, S.; Pal, J.K. Carbohydrate-Binding Agents: Potential of Repurposing for COVID-19 Therapy. Curr. Protein Pept. Sci. 2020, 21, 1–13. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seaweed Family | Seaweed Species | Lectin | Structural Scaffold | Ref. |
|---|---|---|---|---|
| Red algae | Agardhiella subulata | ASL-1, | β-barrel | [7] |
| ASL-2 | β-barrel | |||
| Carpopeltis flabellata | CFA | unknown | [36] | |
| Eucheuma amakusaensis | EAA-1 | β-barrel | [37] | |
| EAA-2 | β-barrel | |||
| EAA-3 | β-barrel | |||
| Eucheuma cottonii | ECA-1 | β-barrel | [37] | |
| ECA-2 | β-barrel | |||
| Eucheuma denticulatum | EDA-1 | β-barrel | [38] | |
| EDA-2 | β-barrel | |||
| Eucheuma serra | ESA-1 | β-barrel | [39,40] | |
| ESA-2 | β-barrel | |||
| Gracilaria bursa-pastoris | GBPL | unknown | [41] | |
| Grateloupia chiangii | GCL | β-prism II | [15] | |
| Griffthsia sp. | griffithsin | β-barrel | [8] | |
| Kappaphycus alvarezii | KAA-2 | β-barrel | [10] | |
| Kappaphycus striatum | KSA-2 | β-barrel | [38] | |
| Meristiella echinocarpa | MEL | β-barrel | [42] | |
| Meristotheca papulosa | MPA-1 | β-barrel | [42] | |
| MPA-2 | β-barrel | |||
| Solieria filiformis | SfL-1 | β-barrel | [12] | |
| SfL-2 | β-barrel | |||
| Solieria robusta | SrL | β-barrel | [3] | |
| Hydropuntia (Gracilaria) fisheri | HFA | β-sandwich | [43] | |
| Porphyra umbilicalis | NgL | β-sandwich | [44] | |
| Yellow-green algae | Nannochloropsis gaditana | BU14 | β-sandwich | [45] |
| Green algae | Boodlea coacta | BCA | β-prism II | [11,46] |
| Bryopsis plumosa | BPL-2 | MFP2-like scaffold | [47] | |
| Enteromorpha prolifera | EPL-1/2 | unknown | [48] | |
| Halimeda renschii | HRL40-1/2 | unknown | [14] | |
| Ostreococcus tauri | OtL | β-sandwich | [49] |
| Seaweed Family | Seaweed Species | Lectin | Structural Scaffold | Ref. |
|---|---|---|---|---|
| Rhodophyceae | Aglaothamnion oosumiense | AOL1 | unknown | [56] |
| Chlorophyceae | Codium fragile | CFL | unknown | [57,58] |
| Seaweed Family | Seaweed Species | Lectin | Structural Scaffold | Ref. |
|---|---|---|---|---|
| Rhodophyceae | Gracilaria tikvahiae | GTL | unknown | [59] |
| Palmaria palmata | PPL | unknown | [60] | |
| Solieria chordalis | ScL | β-barrel | [61] | |
| Phaeophyceae | Fucus vesiculosus | ? * | unknown | [62] |
| RBD/S-GPT | PDB Code | RX/Cryo-Em | Resolution (Å) | Ref. |
|---|---|---|---|---|
| RBD | 6W41 | RX | 3.084 Å | [64] |
| RBD | 6XC2, 6XC3, 6XC4, 6XC7 | RX | 2.3 Å–3.11 Å | [65] |
| RBD | 6XDG | Cryo-Em | 3.9 Å | [66] |
| RBD | 6XE1 | RX | 2.75 Å | [67] |
| RBD | 6YLA, 6YM0, 6YOM | RX, Cryo-Em | 2.42 Å–4.36 Å | Tbp |
| RBD | 6YZ7, 6Z2M, 6ZH9 | RX | 2.71 Å–3.31 Å | Tbp |
| RBD | 6ZCZ, 6ZER, 6ZFO | RX, Cryo-Em | 2.65 Å–4.4 Å | [68] |
| RBD | 7BWJ | RX | 2.85 Å | [69] |
| RBD | 7BZ5 | RX | 1.84 Å | [70] |
| RBD | 7C01 | RX | 2.88 Å | [28] |
| RBD | 7C8V | RX | 2.15 Å | Tbp |
| RBD | 7JMP | RX | 1.712 Å | [71] |
| S-GPT | 6VYB | Cryo-Em | 3.2 Å | [72] |
| S-GPT | 6WPT | Cryo-Em | 3.1 Å–3.7 Å | [73] |
| S-GPT | 6X2A | Cryo-Em | 2.9 Å–3.6 Å | [74] |
| S-GPT | 6X6P | Cryo-Em | 3.22 Å | [29] |
| S-GPT | 6X79 | Cryo-Em | 2.9 Å | [75] |
| S-GPT | 6XCN | Cryo-Em | 3.42 Å–3.66 Å | [76] |
| S-GPT | 6XEY | Cryo-Em | 3.27 Å | [77] |
| S-GPT | 6XF5,6XF6 | Cryo-Em | 3.45 Å–4.0 Å | Tbp |
| S-GPT | 6XKL | Cryo-Em | 3.21 Å | [78] |
| S-GPT | 6XLU,6XM0,6XM3,6XM4,6XM5 | Cryo-Em | 2.4 Å–3.1 Å | [31] |
| S-GPT | 6XR8 | Cryo-Em | 2.9 Å | [79] |
| S-GPT | 6XS6 | Cryo-Em | 3.7 Å | [80] |
| S-GPT | 6Z43 | Cryo-Em | 3.3 Å | Tbp |
| S-GPT | 6Z97 | Cryo-Em | 3.4 Å | [81] |
| S-GPT | 6ZDH | Cryo-Em | 3.7 Å | [68] |
| S-GPT | 6ZGE,6ZGH,6ZGG,6ZGI,6ZHD | Cryo-Em | 2.6 Å–6.8 Å | Tbp |
| S-GPT | 6ZOX,6ZOY,6ZOZ,6ZP0,6ZP1,6ZP2 | Cryo-Em | 3.0 Å–3.5 Å | [29] |
| S-GPT | 6ZOW,6ZP5, 6ZP7 | Cryo-Em | 3.0 Å–3.3 Å | [82] |
| S-GPT | 6ZWV | Cryo-Em | 3.5 Å | [83] |
| S-GPT | 7BYR | Cryo-Em | 3.84 Å | [84] |
| S-GPT | 7CN9 | Cryo-Em | 4.7 Å | [85] |
| S-GPT | 7JJI | Cryo-Em | 3.6 Å | [86] |
| Seaweed Family | Lectin | Virus | Glycan Recognized | Ref. |
|---|---|---|---|---|
| Rhodophyceae | ESA-2 (Eucheuma serra) | Influenza | High-mannose | [12] |
| GCL (Grateloupia chiangii) | Influenza | High-mannose | [15] | |
| Herpes | ||||
| Griffithsin (Griffithsia sp.) | HIV-1 | High-mannose | [8] | |
| Hepatitis C | ||||
| SARS-CoV | [9] | |||
| HRL-40 (Halimeda renschii)) | Influenza | High-mannose | [14] | |
| KAA-2 (Kappaphycus alvarezii) | Influenza | High-mannose | [10] | |
| HIV-1 | [13] | |||
| Chlorophyceae | BCA (Boodlea coacta) | Influenza | High-mannose | [11] |
| HIV-1 | ||||
| * Cyanobacteria | MVN (Microcystis aeruginosa) | HIV-1 | High-mannose | [22] |
| MVL (Microcystis viridis) | HIV-1 | High-mannose | [23] | |
| CV-N (Nostoc ellipsosprum) | HIV-1 | High-mannose | [17,18] | |
| Herpes | [18] | |||
| Ebola | [19] | |||
| Hepatitis C | [20] | |||
| Influenza | [21] | |||
| OAA (Oscillatoria agardhii) | HIV-1 | High-mannose | [16] | |
| SVN (Scytonema varium) | HIV-1 | High-mannose | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barre, A.; Damme, E.J.M.V.; Simplicien, M.; Benoist, H.; Rougé, P. Man-Specific, GalNAc/T/Tn-Specific and Neu5Ac-Specific Seaweed Lectins as Glycan Probes for the SARS-CoV-2 (COVID-19) Coronavirus. Mar. Drugs 2020, 18, 543. https://doi.org/10.3390/md18110543
Barre A, Damme EJMV, Simplicien M, Benoist H, Rougé P. Man-Specific, GalNAc/T/Tn-Specific and Neu5Ac-Specific Seaweed Lectins as Glycan Probes for the SARS-CoV-2 (COVID-19) Coronavirus. Marine Drugs. 2020; 18(11):543. https://doi.org/10.3390/md18110543
Chicago/Turabian StyleBarre, Annick, Els J.M. Van Damme, Mathias Simplicien, Hervé Benoist, and Pierre Rougé. 2020. "Man-Specific, GalNAc/T/Tn-Specific and Neu5Ac-Specific Seaweed Lectins as Glycan Probes for the SARS-CoV-2 (COVID-19) Coronavirus" Marine Drugs 18, no. 11: 543. https://doi.org/10.3390/md18110543
APA StyleBarre, A., Damme, E. J. M. V., Simplicien, M., Benoist, H., & Rougé, P. (2020). Man-Specific, GalNAc/T/Tn-Specific and Neu5Ac-Specific Seaweed Lectins as Glycan Probes for the SARS-CoV-2 (COVID-19) Coronavirus. Marine Drugs, 18(11), 543. https://doi.org/10.3390/md18110543