Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Results
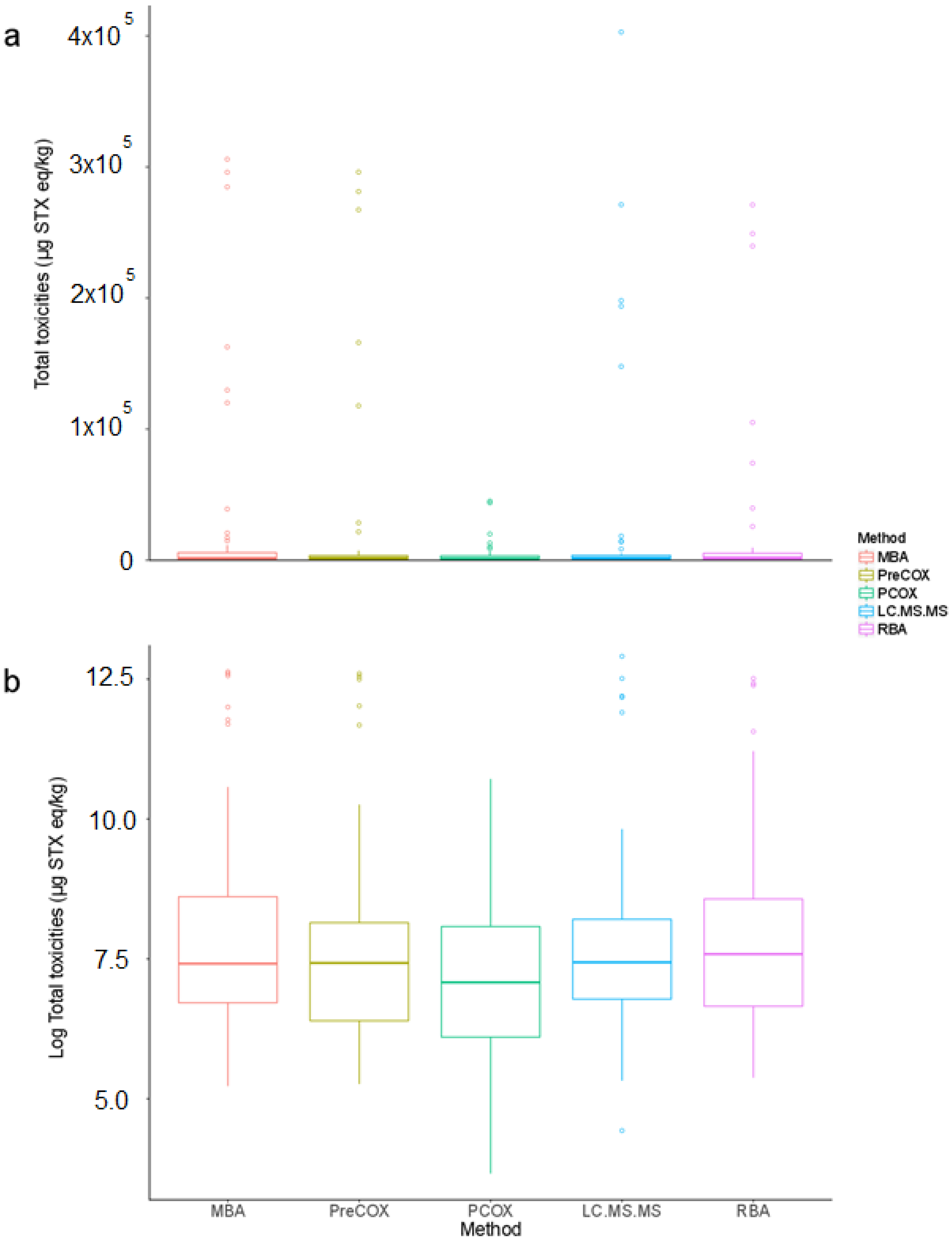
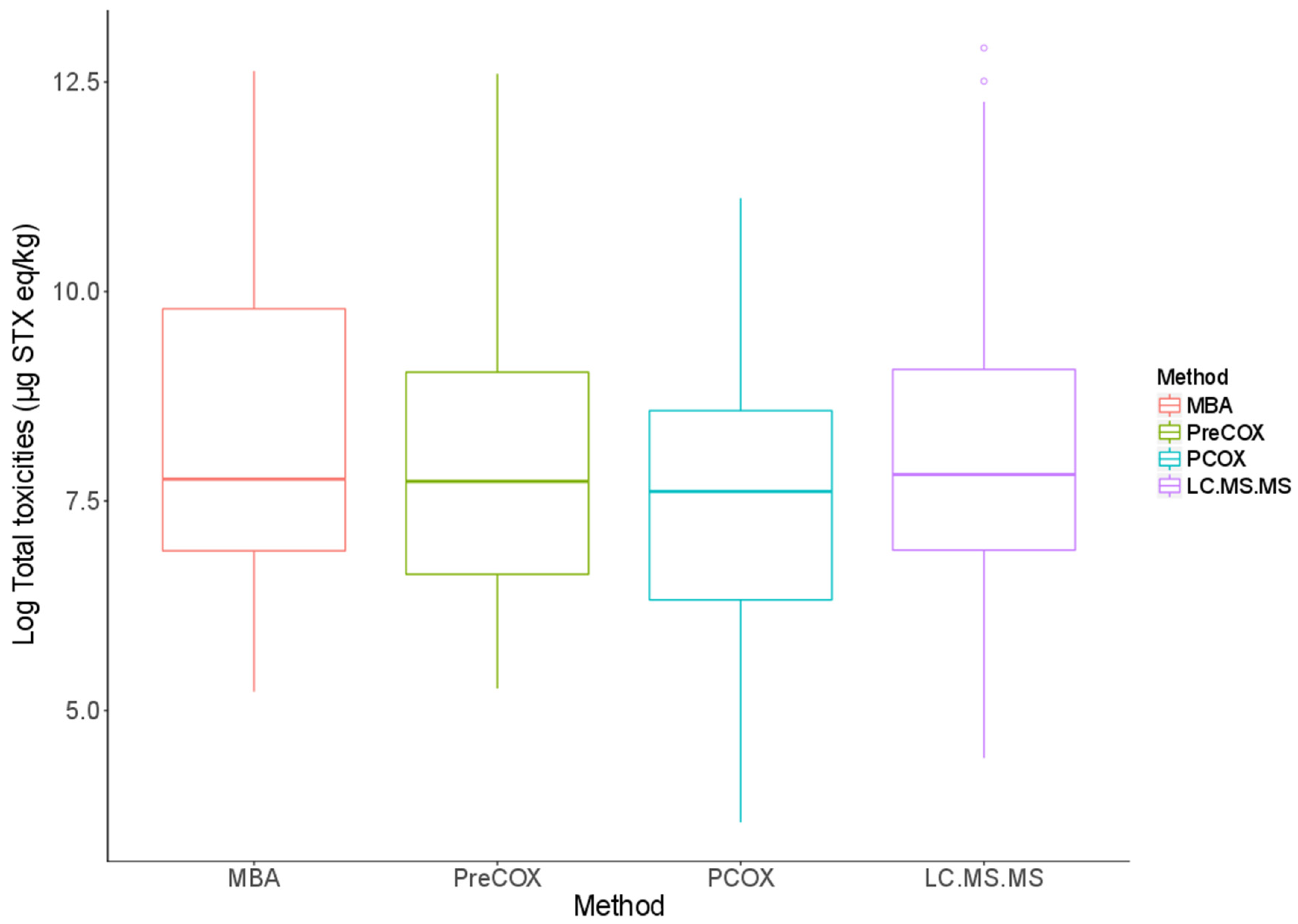
2.1. Toxicities
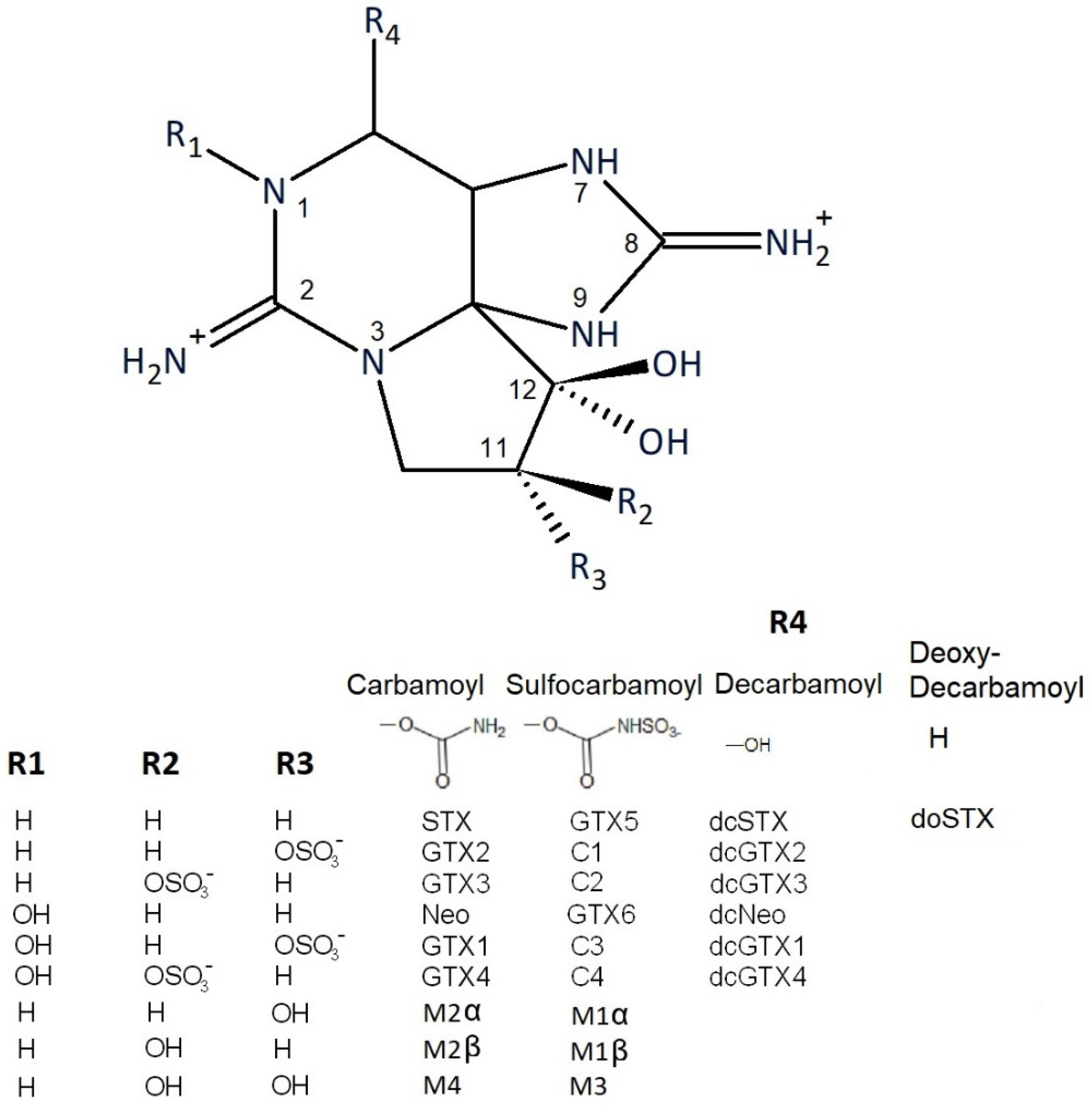
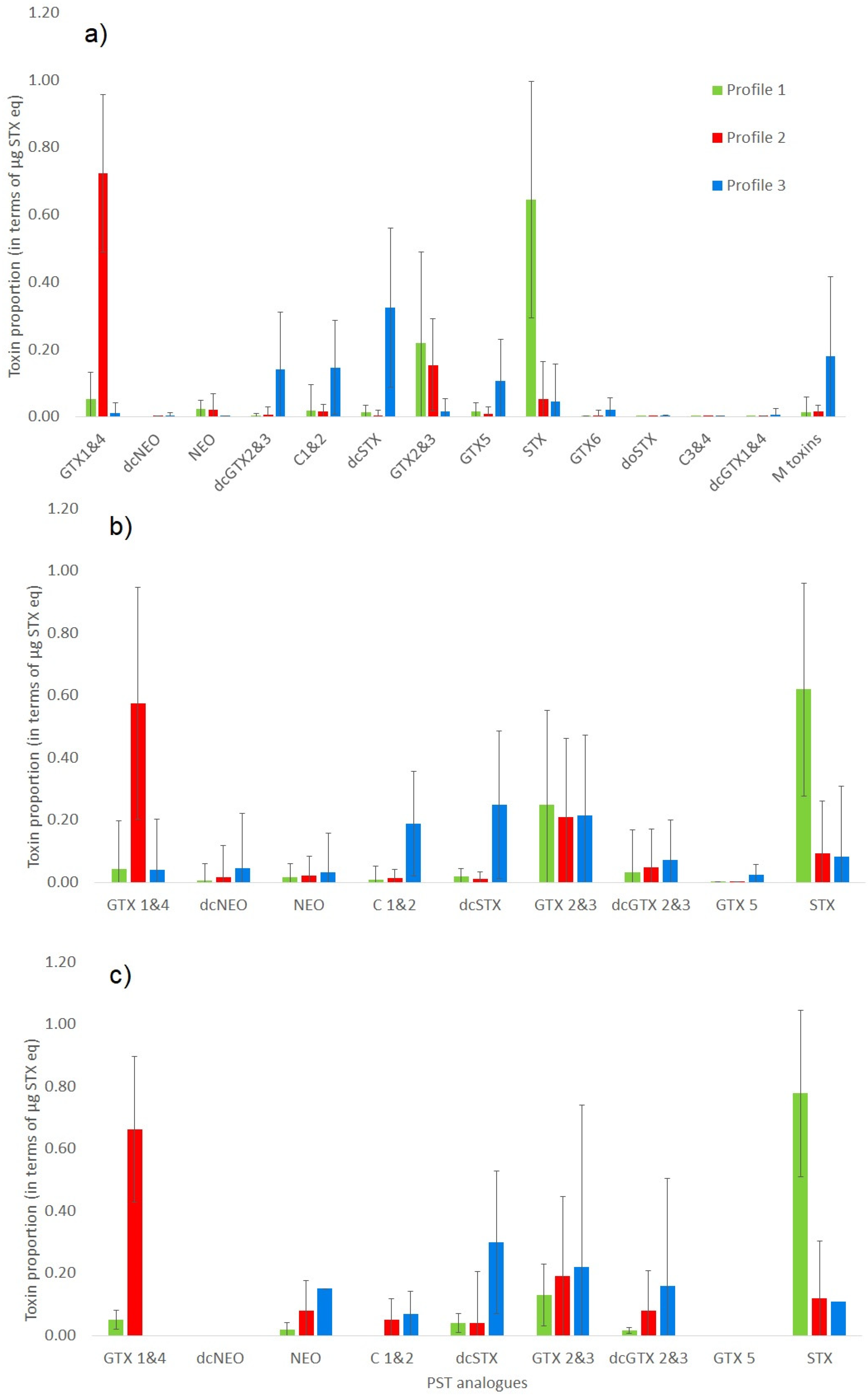
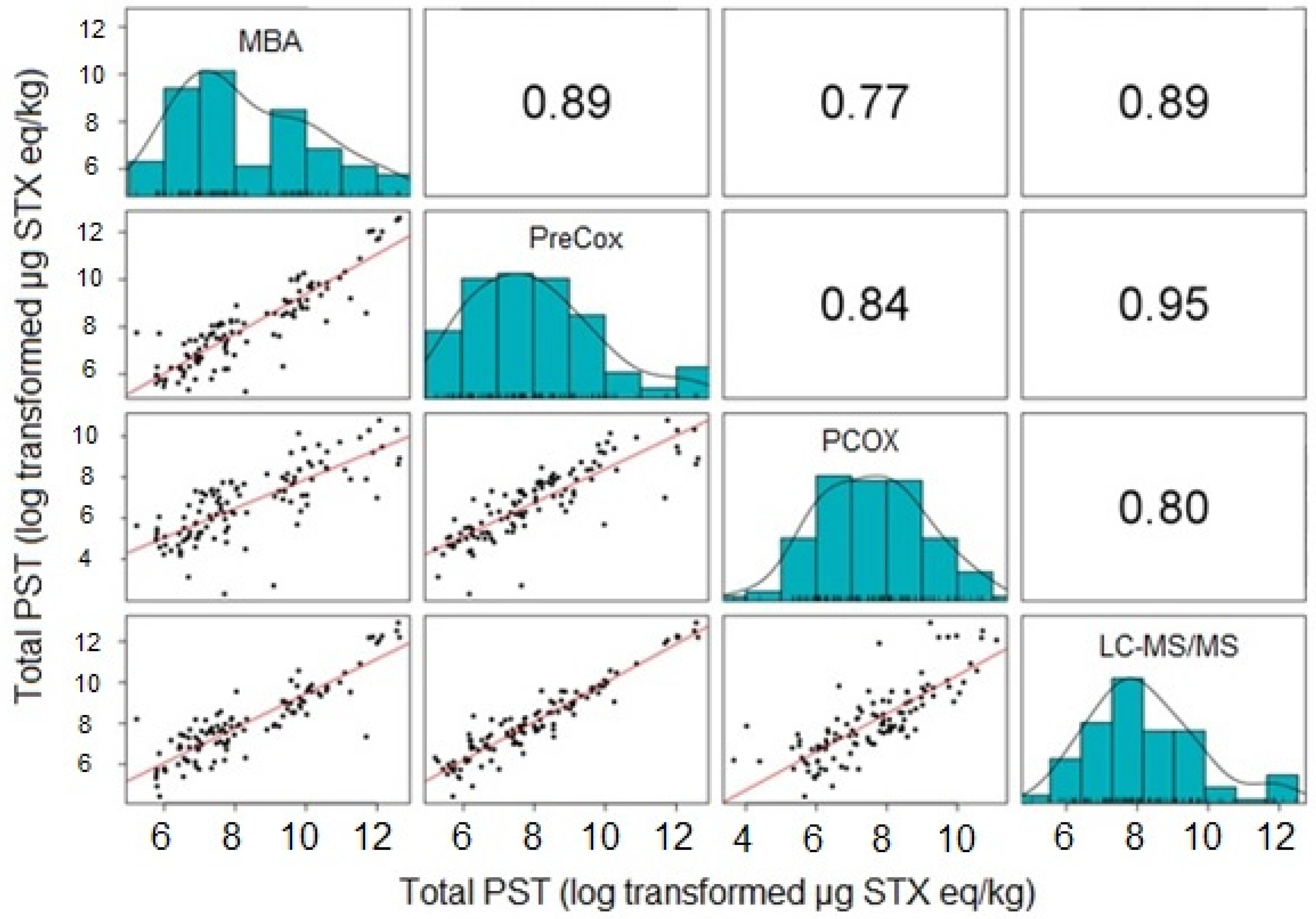
2.2. Quantitative Comparison and Toxin Profiles
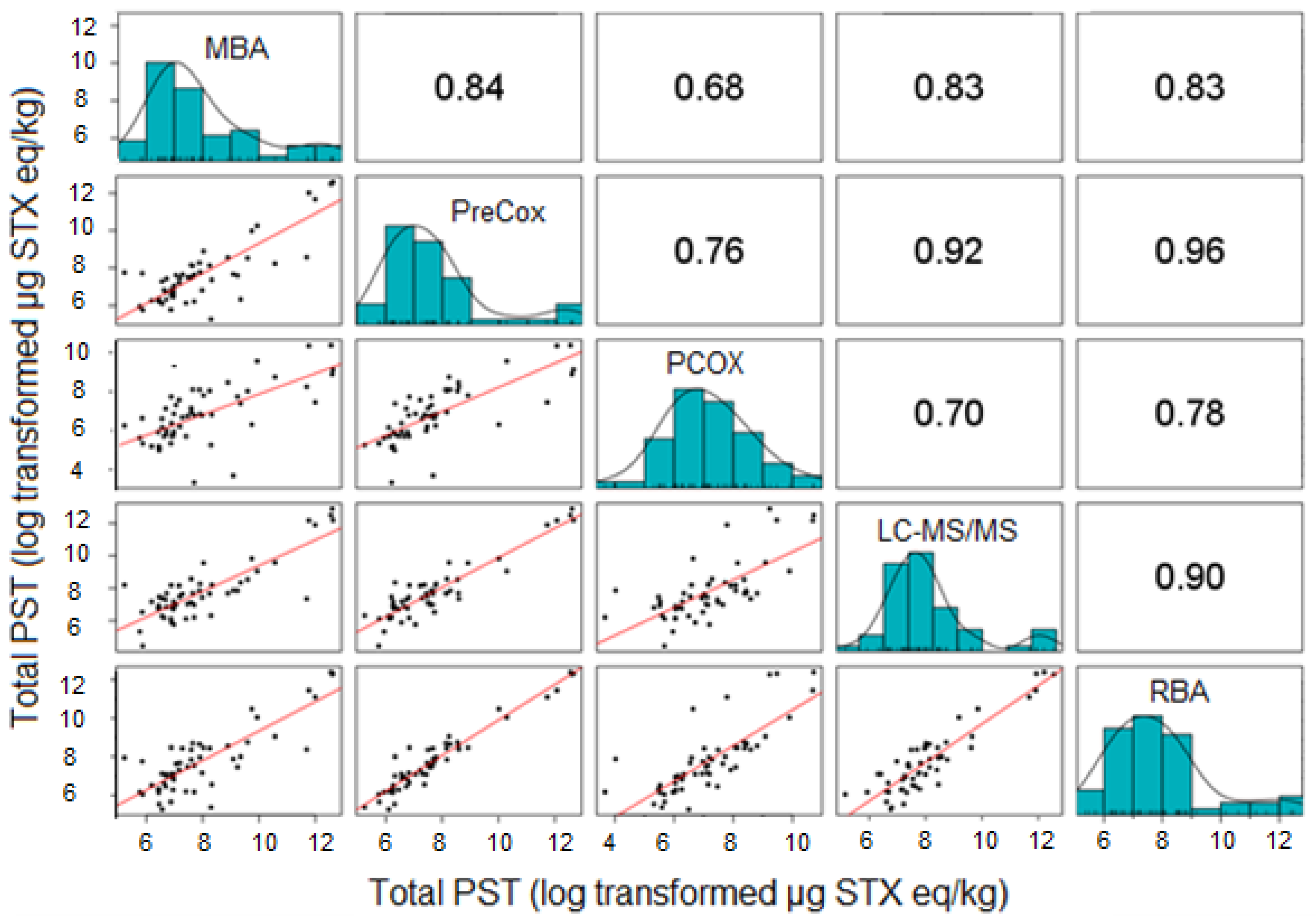
2.2.1. Comparison of Five Quantitative Methods
2.2.2. Comparison of Four Quantitative Methods
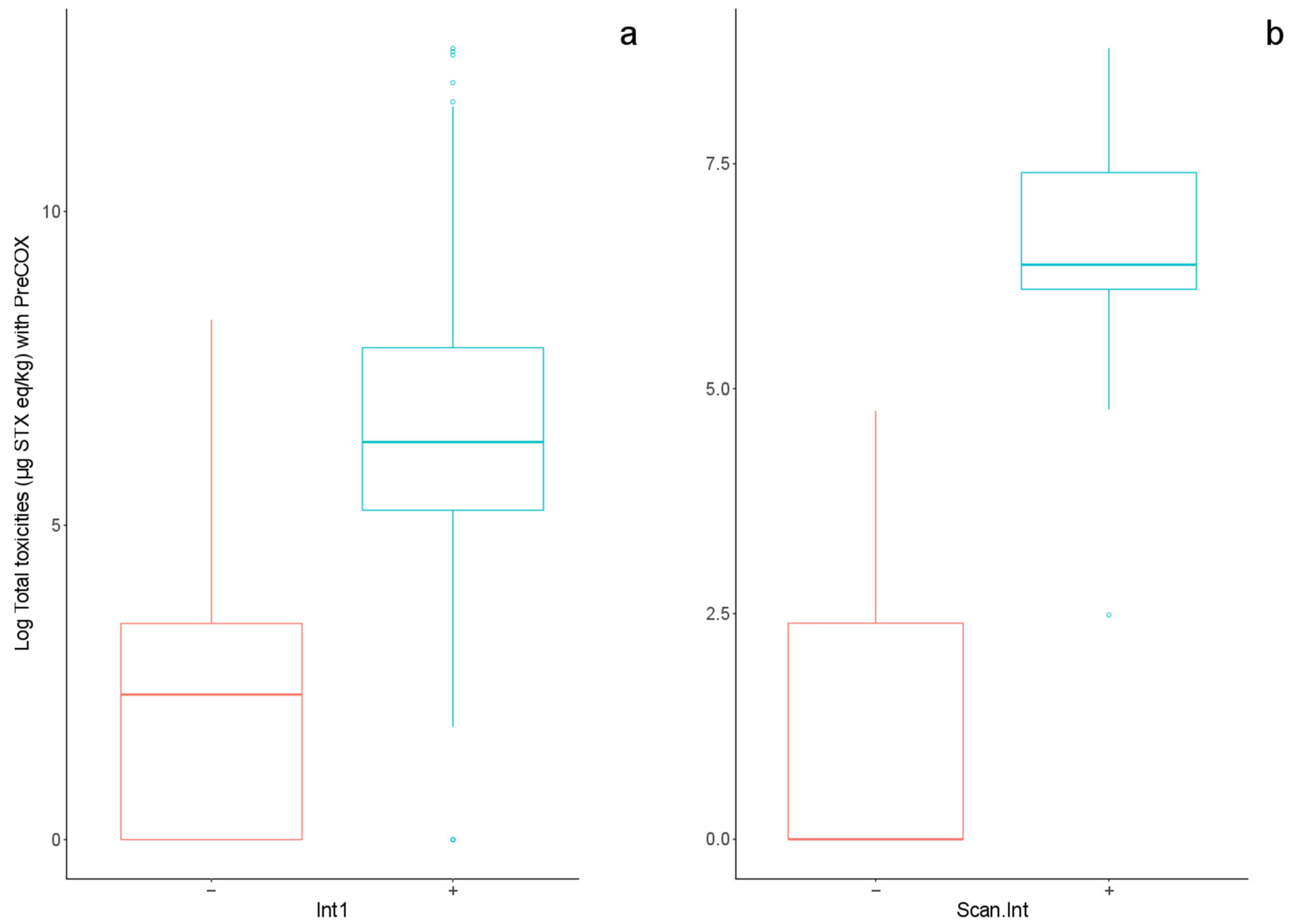
2.2.3. Comparison of Toxin Profiles between Methods
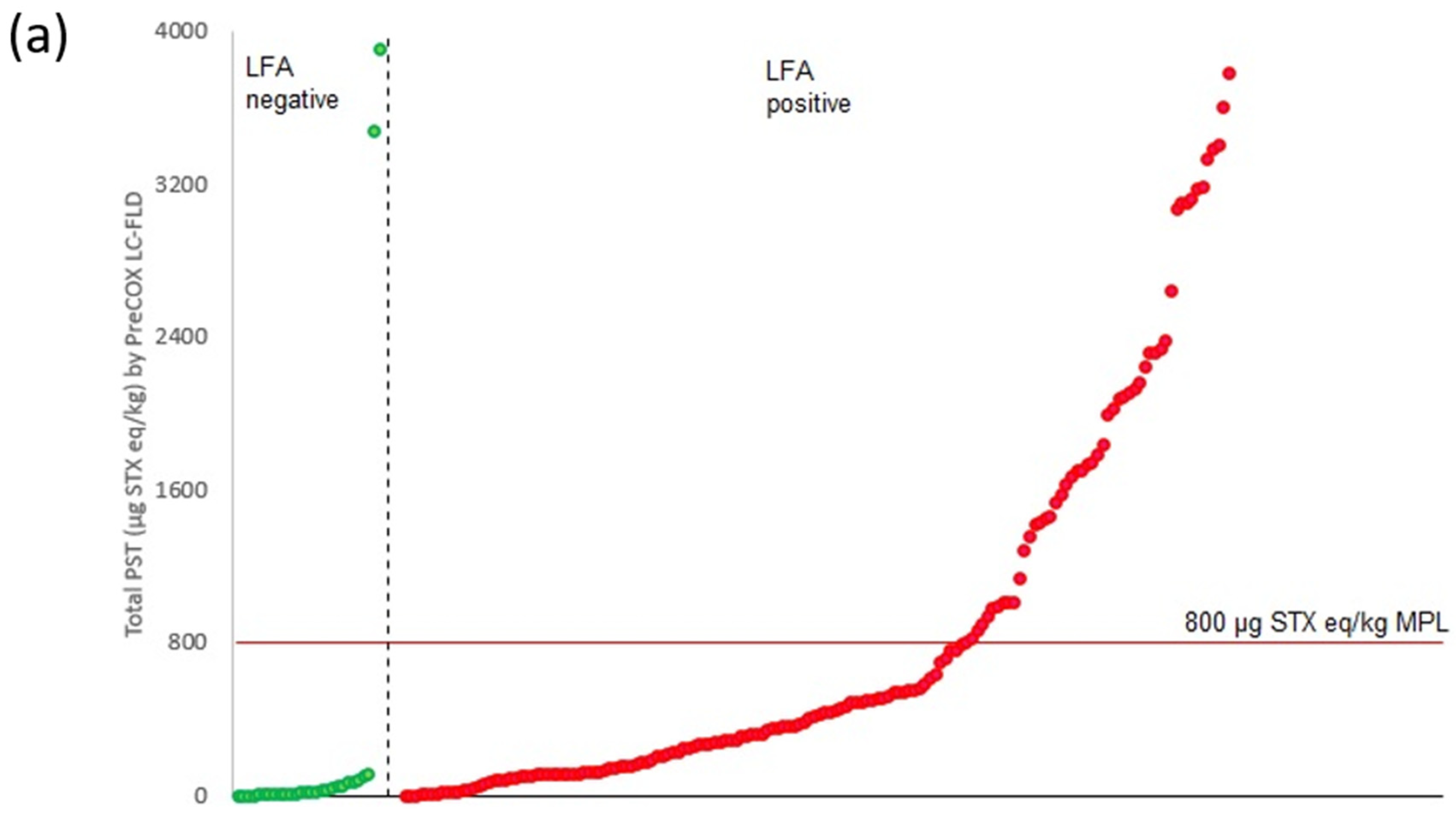
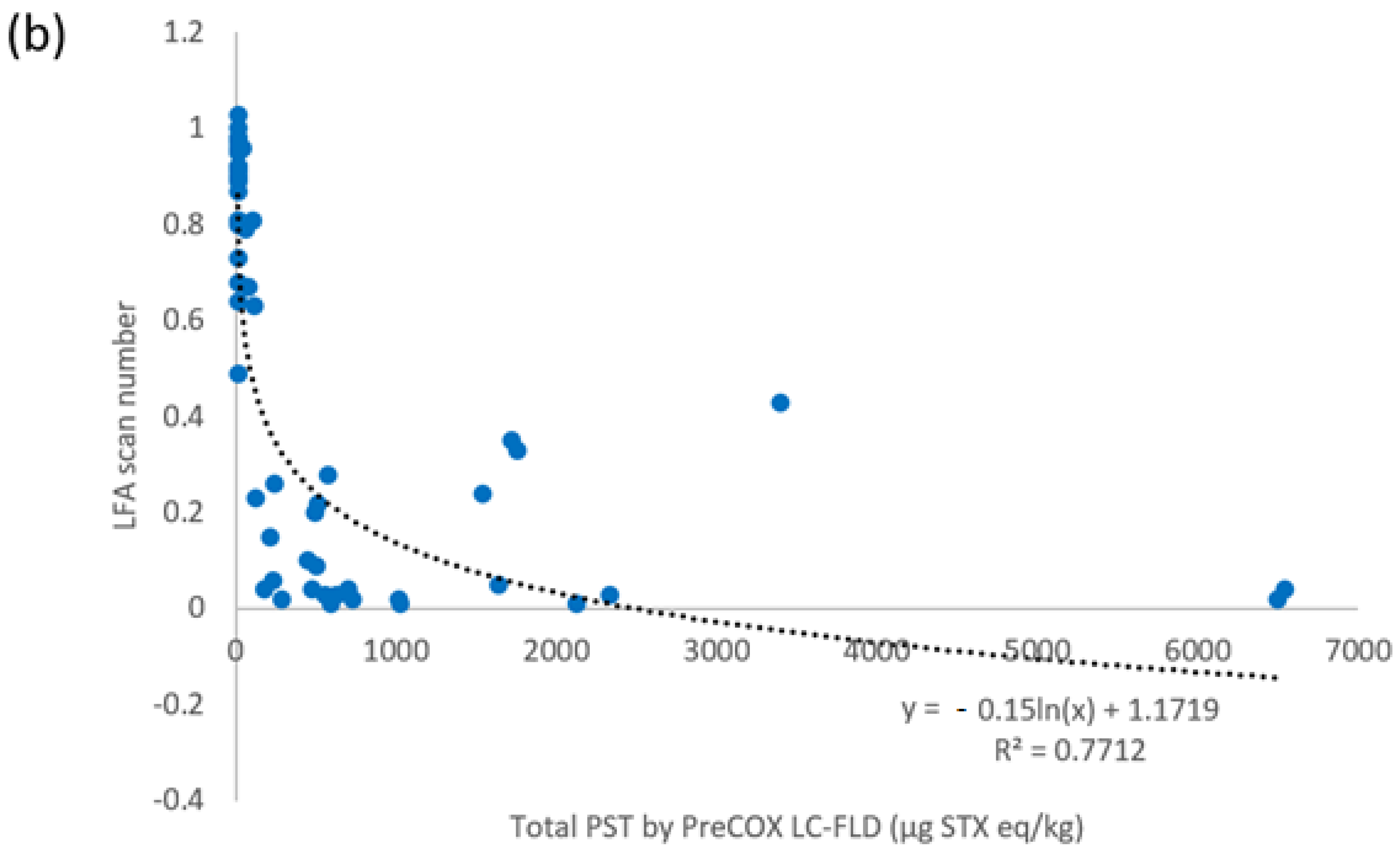
2.3. LFA vs. PreCOX LC-FLD
3. Discussion
3.1. PST Outbreaks in Latin America and Social Impacts
3.2. Method Comparison
3.2.1. MBA and RBA
3.2.2. Instrumental Methods
3.2.3. LFA
3.3. Method Implementation
4. Materials and Methods
4.1. Samples
4.2. Reagents and Chemicals
4.3. Methods of Analysis
4.3.1. Shellfish Extraction and MBA
4.3.2. PreCOX LC-FLD
4.3.3. PCOX LC-FLD
4.3.4. UHPLC-HILIC-MS/MS
4.3.5. Lateral Flow Assay (LFA)
4.3.6. Receptor Binding Assay (RBA)
4.4. Data Assessment
4.4.1. Toxin Results
4.4.2. Statistical Assessment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hallegraef, G.M. Harmful algal blooms: A global overview. In Manual on Harmful Marine Microalgae; Hallegraef, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 2003; pp. 25–49. [Google Scholar]
- Llewellyn, L.; Negri, A.; Robertson, A. Paralytic shellfish toxins in Tropical Oceans. Toxin Rev. 2006, 25, 159–196. [Google Scholar] [CrossRef]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO (Food and Agriculture Organisation of the United Nations). Marine Biotoxins, Paper 80; FAO Food and Nutrition: Rome, Italy, 2004; pp. 5–52. [Google Scholar]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, Y. Post-column derivatisation liquid chromatography method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar] [CrossRef]
- Vale, P. Metabolites of saxitoxin analogues in bivalves contaminated by Gymnodinium catenatum. Toxicon 2010, 55, 162–165. [Google Scholar] [CrossRef]
- Negri, A.P.; Bolch, C.J.S.; Geier, S.; Green, D.H.; Park, T.-G.; Blackburn, S.I. Widespread presence of hydrophobic paralytic shellfish toxins in Gymnodinium catenatum. Harmful Algae 2007, 6, 774–780. [Google Scholar] [CrossRef]
- Lagos, N. Paralytic shellfish poisoning phycotoxins: Occurrence in South America. Comments Toxicol. 2003, 9, 175–193. [Google Scholar] [CrossRef]
- Montoya, N.G.; Akselman, R.; Carignan, M.O.; Carreto, J.I. Pigment profile and toxin composition during a red tide of Gymnodinium catenatum Graham and Myrionecta rubra (Lohman) Jankowski in coastal waters off Mar del Plata, Argentina. Afr. J. Mar. Sci. 2006, 28, 199–202. [Google Scholar] [CrossRef]
- Akselman, R.; Reguera, B.; Lion, M. HAB-MAPS of toxic marine microalgae in coastal and shelf waters of South America. In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; International Society for the study of Harmful Algae: Copenhagen, Denmark, 2006; Volume 12, pp. 243–245. [Google Scholar]
- Lewitus, A.; Horner, R.; Caron, D.; Garcia-Mendoza, E.; Hickey, B.; Hunter, M.; Huppert, D.; Kudela, R.; Langlois, G.; Largier, J.; et al. Harmful algal blooms along the North American west coast region: History, trends, causes and impacts. Harmful Algae 2012, 19, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Fabro, E.; Krock, B.; Torres, A.I.; Paparazzo, F.E.; Schloss, I.R.; Ferreyra, G.A.; Almandoz, G.O. Toxigenic dinoflagellates and associated toxins in San Jorge Gulf, Argentina. Oceanography 2018, 31, 145–153. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Duran-Riveroll, L.M.; Bustillos-Guzmán, J.J.; Leyva-Valencia, I.; Lopez-Cortes, D.J.; López-Cortés, D.J.; Nuñez-Vázquez, E.J.; Hernández-Sandoval, F.E.; Ramirez-Rodriguez, D.V. Paralytic toxin producing dinoflagellates in Latin America: Ecology and physiology. Front. Mar. Sci. 2019, 6, 42. [Google Scholar] [CrossRef]
- Balech, E. El plancton de Mar del Plata durante el periodo 1961–1962. Bol. Inst. Biol. Mar. 1964, 4, 49. [Google Scholar]
- Akselman, R.; Carreto, J.I.; Montoya, N.G. Gymnodinium catenatum and autumn toxicity in northern shelf waters of Argentina. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; UNESCO, Xunta de Galicia: Paris, France, 1998; pp. 122–123. [Google Scholar]
- Andrinolo, D.; Santinelli, N.; Otano, S.; Sastre, V.; Lagos, N. Paralytic shellfish toxins in mussel and Alexandrium tamarense at Valdes Peninsula, Chubut, Patagonia Argentina: Kinetics of a natural depuration. J. Shellfish Res. 1999, 18, 203–209. [Google Scholar]
- Santinelli, N.; Gaille, G.; Lettieri, A.; Intergovernmental Oceanographic Commission (IOC) of UNESCO. Harmful algae and PSP toxicity along north Patagonia coast. Harmful Algae News 1994, 9, 6. [Google Scholar]
- Carreto, J.I.; Montoya, N.; Colleoni, A.D.C.; Akselman, R. Alexandrium tamarense blooms and shellfish toxicity in the Argentine Sea: A retrospective view. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia y IOC—UNESCO: Paris, France, 1998; pp. 131–134. [Google Scholar]
- Gayoso, A.M. Observations on Alexandrium tamarense (Lebour) Balech and other dinoflagellate populations in Golfo Nuevo, Patagonia (Argentina). J. Plankton Res. 2001, 23, 463–468. [Google Scholar] [CrossRef]
- Benavides, H.; Prado, L.; Diaz, S.; Carreto, J.I. An exceptional bloom of Alexandrium catenella in the Beagle Channel, Argentina. In Harmful Marine Algal Blooms, Proceedings of the Sixth International Conference on Toxic Marine Phytoplankton, Nantes, France, 18–22 October 1993; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier: Paris, France, 1995. [Google Scholar]
- Almandoz, G.O.; Harnando, M.P.; Ferreyra, G.A.; Schloss, I.R.; Ferrario, M.E. Seaonsal phytoplankton dynamics in extreme southern South America (Beagle Channel, Argentina). J. Sea Res. 2011, 66, 47–57. [Google Scholar] [CrossRef]
- Almandoz, G.O.; Cefarelli, A.O.; Diodato, S.; Montoya, N.G.; Benavides, H.R.; Carigan, M.; Hernando, M.; Fabro, E.; Metfies, K.; Lundholm, N.; et al. Harmful phytoplankton in the Beagle Channel (South America) as a potential threat to aquaculture activities. Mar. Pollut. Bull. 2019, 145, 105–117. [Google Scholar] [CrossRef]
- Brazeiro, A.; Mendez, S.M.; Ferrari, G. First toxic bloom of Alexandrium tamarense in Uruguay: Associated environmental factors. Rev. Atl. 1997, 19, 19–29. [Google Scholar]
- Mendez, S.; Ferrari, G. Floraciones toxicas de Gymnodinium catenatum en aguas uruguayas. Publ. Com. Tec. Mix. Frente Marit. 2003, 19, 97–102. [Google Scholar]
- Mendez, S.M.; Medina, D. Twenty-three years of red tide monitoring at fixed stations along the coast of Uruguay. In Harmful Algae 2002; Steidinger, K.A., Landsberg, J.H., Tomas, C.R., Vargo, G.A., Eds.; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography and Intergovernmental Oceanographic Commission of UNESCO: Tallahassee, FL, USA, 2004; pp. 341–343. [Google Scholar]
- Garate-Lizarraga, I.; Bustillos-Guzman, J.J.; Morquecho, L.; Band-Schmidt, C.J.; Alonso-Rodriguez, R.; Erler, K.; Luckas, B.; Reyes-Salinas, A.; Gongora-Gonzalez, D.T. Comparative paralytic shellfish toxin profiles in the strains of Gymnodinium catenatum Graham from the Gulf of California, Mexico. Mar. Poll. Bull. 2005, 50, 208–236. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Bustillos-Guzmán, J.J.; López-Cortés, D.J.; Gárate-Lizárraga, L.; Nuñez-Vázquez, E.J.; Hernández-Sandoval, F.E. Ecological and physiological studies of Gymnodinium catenatum in the Mexican Pacific: A review. Mar. Drugs 2010, 8, 1935–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa, J.L.; Sánchez-Paz, A.; Cruz-Villacorta, A.; Nuñez-Vázquez, E.; Sierra Beltrán, A. Toxic events in the northwest Pacific coastline of Mexico during 1992–1995: Origin and impacts. Hydrobiologica 1997, 352, 195–200. [Google Scholar] [CrossRef]
- Bustillos-Guzmán, J.J.; Leyva-Valencia, I.; Hernández-Sandoval, F.E.; Band-Schmidt, C.J.; López-Cortés, D.J.; Núñez-Vázquez, E.J. Ficotoxinas en aguas del Golfo de California: Una revision. In Florecimientos Algales Nocivos en México; García-Mendoza, E., Quijano-Scheggia, S.I., Olivos-Ortíz, A., Núñez-Vazquez, E.J., Eds.; CICESE: Ensenada, Mexico, 2016; pp. 162–179. [Google Scholar]
- Santiago-Morales, S.I. Florecimientos algales nocivos en la costa de Oaxaca. In Florecimientos Algales Nocivos en México; García-Mendoza, E., Quijano-Scheggia, S.I., Olivos-Ortíz, A., Núñez-Vazquez, E.J., Eds.; CICESE: Ensenada, Mexico, 2016; pp. 242–255. [Google Scholar]
- Morquecho, L. Pyrodinium bahamense one of the most significant harmful dinoflagellate in Mexico. Front. Mar. Sci. 2019, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Becerrill, D.U.; Lau, W.L.S.; Hil, K.S.; Leaw, C.P.; Varona-Cordero, F.; Lim, P.T. Abundance and distribution of the potentially toxic thecate dinoflagellate Alexandrium tamiyavanichii (Dinophyceae) in the central Mexican Pacific, using the quantitative PCR method. Front. Mar. Sci. 2018, 5, 366. [Google Scholar] [CrossRef]
- Altamirano, R.C.; Sierra-Beltrán, A.P. Biotoxins from freshwater and marine harmful algal blooms occurring in Mexico. Toxin Rev. 2008, 27, 27–77. [Google Scholar] [CrossRef]
- Okolodkov, Y.B.; Garate-Lizárraga, I. An annotated checklist of dinoflagellates (Dinophyceae) from the Mexican Pacific. Acta Bot. Mex. 2006, 74, 1–154. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Becerril, D.U.; Alonso-Rodiguez, R.; Alvarez-Góngora, C.; Barón-Campis, S.A.; Ceballos-Corona, G.; Herrera-Silveira, J.; Meave del Castillo, M.E.; Juárez-Ruiz, N.; Merino-Virgilio, F.; Morales-Blake, A.; et al. Toxic and harmful marine phytoplankton and microalgae (HABs) in Mexican Coasts. J. Environ. Sci. Health Part A 2007, 42, 1349–1363. [Google Scholar] [CrossRef]
- Guzman, L.; Vivanco, X.; Vidal, G.; Pizarro, G.; Hernández, C.; Tocornal, M.A. Spatial and temporal variability of Alexandrium catenella and PSP in southern Chile (438–558S) (May 2006–July 2010). In Proceedings of the 14th International Conference on Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Pagou, K.A., Hallegraeff, G.M., Eds.; International Society for the Study of Harmful Algae, Intergovernmental Oceanographic Commission of UNESCO: Copenhagen, Denmark, 2013; pp. 69–71. [Google Scholar]
- Hernández, C.; Díaz, P.A.; Molinet, C.; Seguel, M. Exceptional climate anomalies and northwards expansion of Paralytic Shellfish Poisoning outbreaks in Southern Chile. Harmful Algae News 2016, 54, 1–2. [Google Scholar]
- Tufts, N.R. Molluscan transvectors of paralytic shellfish poisoning. In Toxic Dinoflagellate Blooms; Taylor, D.L., Seliger, H.H., Eds.; Elsevier: New York, NY, USA, 1979; pp. 403–408. [Google Scholar]
- Shumway, S.E. Phycotoxin-related shellfish poisoning: Bivalve molluscs are not the only vectors. Rev. Fish. Sci. 1995, 3, 1–31. [Google Scholar] [CrossRef]
- Lasta, M.; Ciocco, N.; Bremec, C.; Roux, A. Moluscos bivalvos y gasterópodos. In El Mar Argentino y sus Recursos Pesqueros; Instituto Nacional de Investigación y Desarrollo Pesquero: Mar del Plata, Argentina, 1998; Volume 2, pp. 115–142. [Google Scholar]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef]
- Turner, A.D.; Tarnovius, S.; Goya, A.B. Paralytic shellfish toxins in the marine gastropod Zidona Dufresnei and Adelomelon Beckii from Argentina: Toxicity and toxin profiles. J. Shellfish Res. 2014, 33, 519–530. [Google Scholar] [CrossRef]
- Goya, A.B.; Tarnovius, S.; Hatfield, R.G.; Coates, L.; Lewis, A.M.; Turner, A.D. Paralytic shellfish poisoning toxicity and associated profiles in bivalve mollusc shellfish from Argentina. Harmful Algae 2020, 99, 101910. [Google Scholar] [CrossRef] [PubMed]
- Elbusto, C.A.; Ballabene, A.C.; Campero, C.M.; Ramírez, E.E.; Villanueva, C.R. Toxina paralizante de los moluscos del Mar Argentino. Acta Bioquímica Latinoam. 1981, XV, 447–456. [Google Scholar]
- Carreto, J.I.; Lasta, M.I.; Negri, R.M.; Benavides, H.R. Los fenómenos de marea y toxicidad de moluscos bivalvos en el Mar Argentino. Contr. Inst. Nac. Investig. Des. Pesq. 1981, 399, 93. [Google Scholar]
- Carreto, J.I.; Elbusto, C.; Sancho, H.; Carignan, M.; Yasumoto, T.; Oshima, Y. Comparative studies on Paralytic Shellfish Toxin profiles of marine snails, mussels and an Alexandrium tamarense isolate from the Mar Del Plata coast (Argentina). Rev. Investig. Des. Pesq. 1996, 10, 101–107. [Google Scholar]
- Carreto, J.I.; Montoya, N.G.; Akselman, R.; Negri, R.M.; Carignan, M.O.; Cucchi Colleoni, D.A. Differences in the PSP toxin profiles of Mytilus Edulis during spring and autumn blooms of Alexandrium tamarense off Mar Del Plata coast, Argentina. In Harmful Algae 2002; Steidinger, K.A., Landsberg, J.H., Tomas, C.R., Vargo, G.A., Eds.; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography and Intergovernmental Oceanographic Commission of UNESCO: Tallahassee, FL, USA, 2002; pp. 100–102. [Google Scholar]
- Goya, A.B.; Maldonado, S. Evolution of PSP toxicity in shellfish from the Beagle Channel (Tierra del Fuego, Argentina): An overview. In Molluscan Shellfish Safety; Sauve, G., Ed.; Springer: Dordrecht, The Netherlands, 2014; Volume XXVI, pp. 15–23. [Google Scholar]
- Guzmán, L.; Campodónico, I.; Antunovic, M. Estudios sobre un florecimiento tóxico causado por Gonyaulax catenella en Magallanes. Distribución y niveles de toxicidad de veneno paralítico de los mariscos (Noviembre de 1972–Noviembre de 1973). Ans. Inst. Pat. Punta Arenas 1975, 6, 209–223. [Google Scholar]
- Carreto, J.I.; Benavides, H.R.; Negri, R.H.; Glorioso, P.D. Toxic red tide in the Argentine Sea. Phytoplankton distribution and survival of the toxic dinoflagellate Gonyaulax excavate in frontal area. J. Plankton Res. 1986, 8, 15–28. [Google Scholar] [CrossRef]
- Esteves, J.L.; Santinelli, N.; Sastre, V.; Diaz, R.; Rivas, O. A toxic dinoflagellate bloom and PSP production associated with upwelling in Golfo Nuevo, Patagonia Argentina. Hydrobiologica 1992, 242, 115–122. [Google Scholar] [CrossRef]
- Ciocco, N. La marisquería mediante buceo en el Golfo San José (Chubut, Argentina). Informe Técnico del Área de Pesca del Plan de Manejo Integrado de la Zona Costera Patagónica (GEF-PNUD, Patagonia, Argentina); 1995; pp. 1–39. ISSN 0328-462X. [Google Scholar]
- Carreto, J.I.; Akselman, R.; Benavides, H.; Montoya, N.G.; Negri, R. El proyecto “Marea Roja” del Instituto Nacional de Investigación y Desarrollo Pesquero. In Floraciones Algales Nocivas en el Cono Sur Americano; Sar, E.A., Ferrario, M.E., Reguera, B., Eds.; Instiuto Español de Oceanografía: Madrid, Spain, 2002; pp. 209–215. [Google Scholar]
- Guzmán, L.; Pacheco, H.; Pizarro, G.; Alarcón, C. Alexandrium catenella y veneno paralizante de los mariscos en Chile. In Floraciones Algales Nocivas en el Cono Sur Americano; Sar, E.A., Ferrario, M.E., Reguera, B., Eds.; Instituto Español de Oceanografía: Madrid, Spain, 2002; pp. 235–256. [Google Scholar]
- Persich, G.R.; Kulisb, D.M.; Lilly, E.L.; Anderson, D.M.; Garcia, V.M.T. Probably origin and toxin profile of Alexandrium tamarense (Lebour) Balech from southern Brazil. Harmful Algae 2006, 5, 36–44. [Google Scholar] [CrossRef]
- Montoya, N.G.; Carignan, M.O.; Carreto, J.I. Alexandrium tamarense/catenella blooms in the Southwestern Atlantic: Paralytic shellfish toxin production and its trophic transference. In Plankton Ecology of the Southwestern Atlantic; Hoffmeyer, M.S., Sabatini, M., Brandini, F.P., Calliari, D.L., Santinelli, N.H., Eds.; Springer: Cham, Switzerland, 2018; pp. 453–476. [Google Scholar]
- Villalobos, L.G.; Santinelli, N.H.; Sastre, A.V.; Marino, G.; Almandoz, G.O. Spatiotemporal distribution of paralytic shellfish poisoning (PSP) toxins in shellfish from Argentine Patagonian coast. Heliyon 2019, 5, e01979. [Google Scholar] [CrossRef] [Green Version]
- Medina, D.; Inocente, G.; López, C. PSP in bivalve molluscs along the Uruguayan coast. In Toxic Phytoplankton Blooms in the Sea, Proceedings of the Fifth International Conference on Toxic Marine Phytoplankton, Newport, RI, USA, 28 October–1 November 1991; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 425–428. [Google Scholar]
- Medina, D.; Goya, A.B.; Rozas, C. Molluscan shellfish safety in South America. In Molluscan Shellfish Safety; Sauvé, G., Ed.; Published Spring Science and Business Media: Dordrecht, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Turner, A.D.; Tarnovius, S.; Medina, D.; Salhi, M. Use of a liquid chromatographic method for assessment of paralytic shellfish poisoning toxin profiles in mussels and clams from Uruguay. J. Shellfish Res. 2015, 34, 1129–1140. [Google Scholar] [CrossRef]
- Saldate-Castañeda, O.; Vázquez-Castellanos, J.L.; Galván, J.; Sánchez-Anguiano, A.; Nazar, A. Intoxicaciones por toxina paralizante de molusco en Oaxaca. Salud Publica Mex. 1991, 33, 240–247. [Google Scholar] [PubMed]
- Gárate-Lizárraga, I.; Bustillos-Guzmán, J.J.; López-Cortex, D.J.; Hernández-Sandoval, F.; Erler, K.; Luckas, B. Paralytic shellfish toxin profiles in net phytoplankton samples from Bahía Concepción, Gulf of California, Mexico. Mar. Poll. Bull. 2006, 52, 800–815. [Google Scholar] [CrossRef] [PubMed]
- Medina-Elizalde, J.; Garcia-Mendoza, E.; Turner, A.D.; Sanchez-Bravo, Y.A.; Murillo-Martinez, R. Transformation and depuration of paralytic shellfish toxins in the geoduck clam Panopea globosa from the Northern Gulf of California. Front. Mar. Sci. 2018, 5, 335. [Google Scholar] [CrossRef]
- García, C.; Bravo, C.; Lagos, M.; Lagos, N. Paralytic shellfish poisoning: Post-mortem analysis of tissue and body fluid samples from human victims in the Patagonia fjords. Toxicon 2004, 43, 149–158. [Google Scholar] [CrossRef]
- Díaz, P.A.; Álvarez, G.; Varela, D.; Pérez-Santos, I.; Díaz, M.; Molinet, C.; Seguel, M.; Aguilera-Belmonte, A.; Guzmán, L.; Uribe, E.; et al. Impacts of harmful algal blooms on the aquaculture industry: Chile as a case study. Perspect. Phycol. 2019, 6, 39–50. [Google Scholar] [CrossRef]
- Molinet, C.; Niklitschek, E.; Seguel, M.; Diaz, P. Trends of natural accumulation and detoxification of paralytic shellfish poison in two bivalves from the northwest Patagonian inland sea. Rev. Biol. Mar. Oceanogr. 2010, 45, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, G.; Diaz, P.A.; Godoy, M.; Araya, M.; Ganuza, I.; Pino, R.; Alvarez, F.; Rengel, J.; Hernandez, C.; Uribe, E.; et al. Paralytic shellfish toxins in surf clams Mesodesma donacium during a large bloom of Alexandrium catenella dinoflagellates associated to an intense shellfish mass mortality. Toxins 2019, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Terrazas, J.O.; Contreras, H.R.; Garcia, C. Prevalence, variability and bioconcentration of saxitoxin-group in different marine species present in the food chain. Toxins 2017, 9, 190. [Google Scholar] [CrossRef]
- Strub, P.T.; James, C.; Montecino, V.; Rutlant, J.A.; Blanco, J.L. Ocean circulation along the southern Chile transition region (38°–46°S): Mean, seasonal and interannual variability, with a focus on 2014–2016. Prog. Oceanogr. 2019, 172, 159–198. [Google Scholar] [CrossRef]
- Trainer, V.L.; Moore, S.K.; Hallegraef, G.; Kudela, R.M.; Clement, A.; Mardones, J.I.; Cochlan, W.P. Pelagic harmful algal blooms and climate change: Lessons from nature’s experiments with extremes. Harmful Algae 2020, 91, 101591. [Google Scholar] [CrossRef] [PubMed]
- Anon. Regulation (EC) No 854/2004 of the European Parliament and of the Council of 29th April 2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption. Off. J. Eur. Union 2004, L226, 83–127. [Google Scholar]
- Anon. AOAC Official method 2005.06 Quantitative Determination of Paralytic Shellfish Poisoning Toxins in Shellfish Using Pre-Chromatographic Oxidation and Liquid Chromatography with Fluorescence Detection; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Anon. AOAC Official method 959.08. Paralytic shellfish poison. Biological method. Final action. In AOAC Official Methods for Analysis, 18th ed.; Natural toxins (Truckses, M.W., chapter ed.); AOAC International: Gaithersburg, MD, USA, 2005; Chapter 49; pp. 79–80. [Google Scholar]
- Anon; Committee on Toxicity of Chemicals in Food, Consumer Products and the Environment. Statement on Risk Assessment and Monitoring of Paralytic Shellfish Poisoning (PSP) Toxins in Support of Human Health. COT Statement 2006/08. July 2006. Available online: http://www.food.gov.uk/science/ouradvisors/toxicity/statements/cotstatements2006/cotstatementpsp200608 (accessed on 20 November 2020).
- Turner, A.D.; Dhanji-Rapkova, M.; Algoet, M.; Suarez-Isla, B.A.; Cordova, M.; Caceres, C.; van de Riet, J.; Murphy, C.J.; Case, M.; Lees, D.N. Investigations into matrix components affecting the performance of the official bioassay reference method for quantitation of paralytic shellfish poisoning toxins in oysters. Toxicon 2012, 59, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.F.; Me’nard, C.; Cleroux, C. Evaluation of prechromatographic oxidation for liquid chromatographic determination of paralytic shellfish poisons in shellfish. J. AOAC Int. 1995, 78, 514–520. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Interlaboratory study. J. AOAC Int. 2004, 87, 83–100. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar] [CrossRef] [Green Version]
- Anon. Commission Regulation (EC) No 1664/2006 of 6th Nov. 2006 amending Regulation (EC) No 2074/2005 as regards implementing measures for certain products of animal origin intended for human consumption and repealing certain implementing measures. Off. J. Eur. Union 2006, L320, 13–45. [Google Scholar]
- Anon. Commission regulation (EU) No 2017/1980 of 31 October 2017 ammending Annex III to Regulation (EV) No 2074/2005 as regards paralytic shellfish poison (PSP) detection method. Off. J. Eur. Union 2017, L285, 8–9. [Google Scholar]
- Turner, A.D.; Hatfield, R.G.; Maskrey, B.H.; Algoet, M.; Lawrence, J.F. Evaluation of the new European Union reference method for paralytic shellfish toxins in shellfish: A review of twelve years regulatory monitoring using pre-column oxidation LC-FLD. Trends Anal. Chem. 2019, 113, 124–139. [Google Scholar] [CrossRef]
- Van de Riet, J.M.; Gibbs, R.S.; Chou, F.W.; Muggah, P.M.; Rourke, W.A.; Burns, G.; Thomas, K.; Quilliam, M.A. Liquid chromatographic post-column oxidation method for analysis of paralytic shellfish toxins in mussels, clams, scallops, and oysters: Single-laboratory validation. J. AOAC Int. 2009, 92, 1690–1704. [Google Scholar]
- Van de Riet, J.M.; Gibbs, R.S.; Muggah, P.M.; Rourke, W.A.; MacNeil, J.D.; Quilliam, M.A. Liquid chromatographic post-column oxidation (PCOX) method for the determination of paralytic shellfish toxins in mussels, clams, oysters and scallops: Collaborative study. J. AOAC Int. 2011, 94, 1154–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anon. AOAC official method 2011.02 determination of paralytic shellfish poisoning toxins in mussels, clams, oysters and scallops. In Post-Column Oxidation Method (PCOX). First Action 2011; AOAC International: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Turner, A.D.; Norton, D.M.; Hatfield, R.G.; Morris, S.; Reese, A.R.; Algoet, M.; Lees, D. N Single laboratory validation of the AOAC HPLC method (2005.06) for mussels: Refinement and extension of the method to additional toxins. J. AOAC Int. 2009, 92, 190–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Norton, D.M.; Hatfield, R.G.; Rapkova-Dhanji, M.; Algoet, M.; Lees, D.N. Single laboratory validation of a refined AOAC LC method for oysters, cockles and clams in UK shellfish. J. AOAC Int. 2010, 93, 1482–1493. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Hatfield, R.G.; Rapkova, M.; Higman, W.; Algoet, M.; Suarez-Isla, B.A.; Cordova, M.; Caceres, C.; van de Riet, J.; Gibbs, R.; et al. Comparison of AOAC 2005.06 LC official method with other methodologies for the quantitation of paralytic shellfish poisoning toxins in UK shellfish species. Anal. Bioanal. Chem. 2011, 399, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Harwood, D.T.; Selwood, A.I.; van Ginkel, R.; Waugh, C.; McNabb, P.S.; Munday, R.; Hay, B.; Thomas, K.; Quilliam, M.A.; Malhi, N.; et al. Paralytic shellfish toxins, including deoxydecarbamoyl-STX, in wild-caught Tasmanian abalone (Halitotis rubra). Toxicon 2014, 90, 213–225. [Google Scholar] [CrossRef]
- NSSP. National Shellfish Sanitation Program (NSSP) Guide for the Control of Molluscan Shellfish. 2013 Revision. 2013. Available online: http://www.fda.gov/downloads/Food/GuidanceRegulation/FederalStateFoodPrograms/UCM415522.pdf (accessed on 17 December 2014).
- Van Dolah, F.M.; Keighfield, T.A.; Doucette, G.J.; Bean, L.; Niedzwiadek, B.; Rawn, D.F.K. Single laboratory validation of the microplate receptor binding assay for paralytic shellfish toxins in shellfish. J. AOAC Int. 2009, 92, 1705–1713. [Google Scholar]
- Van Dolah, F.M.; Fire, S.E.; Keighfield, T.A.; Mikulski, C.M.; Doucette, G.J. Determination of paralytic shellfish toxins in shellfish by receptor binding assay: Collaborative study. J. AOAC Int. 2012, 95, 795–812. [Google Scholar] [CrossRef]
- Anon. AOAC Official Method 2011.27. Paralytic Shellfish Toxins (PSTs) in Shellfish, Receptor Binding Assay; AOAC International: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single laboratory validation of a multi-toxin UPLC-HILIC-MS/MS method for quantitation of paralytic shellfish toxins in bivalve shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Fong, S.Y.T.; Hungerford, J.; McNabb, P.S.; Boundy, M.J.; Harwood, D.T. Ultrahigh-performance hydrophilic interaction liquid chromatography with tandem mass spectrometry method for the determination of paralytic shellfish toxins and tetrodotoxin in mussels, oysters, clams, cockles and scallops: Collaborative study. J. AOAC Int. 2020, 103, 1–35. [Google Scholar] [CrossRef]
- Laycock, M.V.; Donovan, M.A.; Easy, D.J. Sensitivity of lateral flow tests to mixtures of saxitoxins and applications to shellfish and phytoplankton monitoring. Toxicon 2010, 55, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Tarnovius, S.; Johnson, S.; Higman, W.A.; Algoet, M. Testing and application of a refined rapid detection method for paralytic shellfish poisoning toxins in UK shellfish. Toxicon 2015, 100, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Baugh, K.A.; Wright, B.; RaLonde, R.; Nance, S.L.; Tatarenkova, N.; Etheridge, S.M.; Lefebvre, K.A. Comparative determination of paralytic shellfish toxins (PSTs) using five different toxin detection methods in shellfish species collected in the Aleutian Islands, Alaska. Toxicon 2009, 54, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R.; Magalhaes, V.F.; Froscio, S.M. Comparison of analytical tools and biological assays for detection of paralytic shellfish poisoning toxins. Anal. Bioanal. Chem. 2010, 397, 1655–1671. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; Alfonso, A.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Comparative analysis of pre- and post-column oxidation methods for detection of paralytic shellfish toxins. Toxicon 2010, 56, 448–457. [Google Scholar] [CrossRef] [PubMed]
- DeGrasse, S.L.; Van de Riet, J.; Hatfield, R.; Turner, A.D. Pre- versus post-column oxidation liquid chromatography fluorescence detection of paralytic shellfish toxins. Toxicon 2011, 57, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gigirey, B.; Rodriguez-Velasco, M.L.; Otero, A.; Vieites, J.M.; Cabado, A.G. A comparative study for PSP toxins quantification by using MBA and HPLC official methods in shellfish. Toxicon 2012, 60, 864–873. [Google Scholar] [CrossRef]
- Turner, A.D.; Lewis, A.M.; Rourke, W.A.; Higman, W.A. Interlaboratory comparison of two AOAC liquid chromatographic fluorescence detection methods for paralytic shellfish toxin analysis through characterization of an oyster reference material. J. AOAC Int. 2014, 97, 380–390. [Google Scholar] [CrossRef]
- Turner, A.D.; Broadwater, M.; Van Dolah, F. Use of the receptor binding assay for the determination of paralytic shellfish poisoning toxins in bivalve molluscs from Great Britain and the assessment of method performance in oysters. Toxicon 2018, 148, 155–164. [Google Scholar] [CrossRef]
- Urbina, M.A.; Luna-Jorquera, G.; Thiel, M.; Acuna-Ruz, T.; Amenabar, M.A.; Andrade, C.; Ahrendt, C.; Castillo, C.; Chevallier, A.; Cornejo-D’Ottone, M.; et al. A country’s response to tackling plastic pollution in aquatic ecosystems: The Chilean way. Aquat. Conserv. 2020. [Google Scholar] [CrossRef]
- Aune, T.; Ramstad, H.; Heinenreich, B.; Landsverk, T.; Waaler, T.; Eggas, E.; Julshamn, K. Zinc accumulation in oysters giving mouse deaths in paralytic shellfish poisoning bioassay. J. Shellfish Res. 1998, 17, 1243–1246. [Google Scholar]
- Vale, P.; de Sampayo, M.A. Determination of paralytic shellfish toxins in Portuguese shellfish by automated pre-column oxidation. Toxicon 2001, 39, 561–571. [Google Scholar] [CrossRef]
- Velez, P.P.; Sierralta, J.; Alcayaga, C.; Fonseca, M.; Loyola, H.; Johns, D.C.; Tomaselli, G.F.; Marban, E.; Suarez-Isla, B.A. A functional assay for paralytic shellfish toxins that uses recombinant sodium channels. Toxicon 2001, 39, 929–935. [Google Scholar] [CrossRef]
- Ruberu, S.R.; Langlois, G.W.; Masuda, M.; Kittredge, C.; Kasum Perera, S.; Kudela, R.M. Receptor binding assay for the detection of paralytic shellfish poisoning toxins: Comparison to the mouse bioassay and applicability under regulatory use. Food Addit. Contam. Part A 2017, 35, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Fast, M.D.; Cembella, A.D.; Ross, N.W. In vitro transformation of paralytic shellfish toxins in the clams Mya arenaria and Protothaca staminea. Harmful Algae 2006, 5, 79–90. [Google Scholar] [CrossRef]
- Artigas, M.L.; Vale, P.J.V.; Gomes, S.S.; Bothelo, M.J.; Rodrigues, S.M.; Amorim, A. Profiles of paralytic shellfish poisoning toxins in shellfish from Portugal explained by carbamoylase activity. J. Chromatogr. A 2007, 1160, 99–105. [Google Scholar] [CrossRef]
- Turner, A.D.; Lewis, A.M.; O’Neil, A.; Hatfield, R.G. Transformation of paralytic shellfish poisoning toxins in UK surf clams (Spisula solida) for targeted production of reference materials. Toxicon 2013, 65, 41–58. [Google Scholar] [CrossRef]
- Hignutt, E. Suitability of postcolumn oxidation liquid chromatography method AOAC 2011.02 for monitoring paralytic shellfish toxins in Alaskan shellfish—Initial pilot study versus mouse bioassay and in-house validation. J. AOAC Int. 2014, 97, 293–298. [Google Scholar] [CrossRef]
- Rourke, W.A.; Murphy, C.J.; Pitcher, G.; Van de Riet, J.M.; Burns, B.G.; Thomas, K.M.; Quilliam, M.A. Rapid postcolumn methodology for determination of paralytic shellfish toxins in shellfish tissue. J. AOAC Int. 2008, 91, 589–597. [Google Scholar] [CrossRef] [Green Version]
- Franca, S.; Alvito, P.; Sousa, I.; Gago, A.; Rodrıguez-Vasquez, J.A.; Leao, J.M.; Comesana, M.; Thibault, P.; Burdaspal, P.; Bustos, J.; et al. The toxin profile of some PSP toxin producing dinoflagellates occurring in Portuguese coastal waters as determined by alternative analytical methods. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 519–522. [Google Scholar]
- Costa, P.R.; Botelho, M.J.; Lefebvre, K.A. Characterization of paralytic shellfish toxins in seawater and sardines (Sardina pilchardus) during blooms of Gymnodinium catenatum. Hydrobiologia 2010, 655, 89–97. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, P.; Guilherme, S.; Barata, M.; Nicolau, L.; Santos, M.A.; Pacheco, M.; Pousao-Ferreira, P. Biotransformation modulation and genotoxicity in white seabream upon exposure to paralytic shellfish toxins produced by Gymnodinium catenatum. Aquat. Toxicol. 2012, 106–107, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Moita, T.; Rodrigues, S.M. Estimating the contribution of N-sulfocarbamoyl paralytic shellfish toxin analogs GTX6 and C3+4 to the toxicity of mussels (Mytilus galloprovincialis) over a bloom of Gymnodinium catenatum. Harmful Algae 2014, 31, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, M.; Pham, L.; Csuti, D.; Dodd, M.; Inami, G.B.; Brenden, R.A. Paralytic shellfish poisoning surveillance in Californica using the Jellett Rapid PSP test. Harmful Algae 2006, 5, 69–73. [Google Scholar] [CrossRef]
- DeGrasse, S.; Conrad, S.; DiStefano, P.; Vanegas, C.; Wallace, D.; Jensen, P.; Hickey, J.M.; Cenci, F.; Pitt, J.; Deardorff, D.; et al. Onboard screening dockside testing as a new means of managing paralytic shellfish poisoning risks in federally closed waters. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 288–300. [Google Scholar] [CrossRef]
- Wong, C.-K.; Hung, P.; Ng, E.A.L.; Lee, K.L.H.; Wong, G.T.C.; Kam, K.-M. Operational application of a rapid antibody-based detection assay for first line screening of paralytic shellfish toxins in shellfish. Harmful Algae 2010, 9, 636–646. [Google Scholar] [CrossRef]
- Dennison, N.; Anderson, D.B. The 3 “R”S approach to marine biotoxin testing in the UK. AATEX 14, special issue, 757–761. In Proceedings of the 6th World Congress on Alternatives and Animal Use in the Life Sciences, Tokyo, Japan, 21–25 August 2007. [Google Scholar]
- Guy, A.L.; Griffin, G. Adopting alternatives for the regulatory monitoring of shellfish for paralytic shellfish poisoning in Canada: Interface between federal regulators, science and ethics. Regul. Toxicol. Pharmacol. 2009, 54, 256–263. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Punn, R.; Algoet, M.; Turner, A.D. A rapid method for the analysis of paralytic shellfish toxins utilising standards pressure HPLC: Refinement of AOAC 2005.06. J. AOAC Int. 2016, 99, 475–480. [Google Scholar] [CrossRef]
- ISSC. Interstate Shellfish Sanitation Conference: Single Laboratory Validation Protocol for Method Approval (Iii). 2020. Available online: http://www.issc.org/laboratory-method-references (accessed on 6 October 2020).
- EFSA. Marine biotoxins in shellfish—Saxitoxin group. Scientific opinion of the panel on contaminants in the food chain. European Food Safety Authority. EFSA J. 2009, 1019, 1–76. [Google Scholar] [CrossRef]
- Laycock, M.V.; Kralovec, J.; Richards, R.C. Some in vitro chemical interconversions of paralytic shellfish poisoning (PSP) toxins useful in the preparation of analytical standards. J. Mar. Biotechnol. 1995, 3, 121–125. [Google Scholar]
- Turner, A.D.; Stubbs, B.; Coates, L.; Dhanji-Rapkova, M.; Hatfield, R.G.; Lewis, A.M.; Rowland-Pilgrim, S.; O’Neil, A.; Stubbs, P.; Ross, S.; et al. Variability of paralytic shellfish toxin occurrence and profiles in bivalve molluscs from Great Britain from official control monitoring as determined by pre-column oxidation liquid chromatography and implications for applying immunochemical tests. Harmful Algae 2014, 31, 87–99. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Shellfish | n | MBA | PreCOX | PCOX | LC-MS/MS | RBA |
|---|---|---|---|---|---|---|---|
| Argentina | Clams | 9 | 1020 | 1174 | 959 | 2001 | 1544 |
| Mussels | 11 | 107,912 | 103,352 | 10,899 | 111,426 | 86,292 | |
| Scallops | 4 | 10,427 | 1480 | 3277 | 5064 | 4147 | |
| Snails | 11 | 5622 | 6775 | 4377 | 3719 | 9283 | |
| Chile | Clams | 3 | 40,717 | 2018 | 1933 | 1675 | 1829 |
| Mussels | 8 | 3902 | 1756 | 1039 | 1547 | 2332 | |
| Oysters | 1 | 1070 | 870 | 523 | 859 | 1016 | |
| Scallops | 2 | 339 | 349 | 339 | 145 | 501 | |
| Mexico | Geoduck (w) | 1 | 4120 | 1582 | 1277 | 3680 | 792 |
| Uruguay | Clams | 2 | 2315 | 356 | 259 | 785 | 380 |
| Mussels | 5 | 5536 | 2395 | 1904 | 4743 | 2641 |
| MBA | PreCOX | PCOX | LCMSMS | |
|---|---|---|---|---|
| PreCOX | 0.225 | |||
| PCOX | 0.00035 | 0.006 | ||
| LCMSMS | 0.766 | 0.884 | 0.004 | |
| RBA | 0.884 | 0.019 | 0.00003 | 0.884 |
| Region | Shellfish | n | MBA | PreCOX | PCOX | LC-MS/MS |
|---|---|---|---|---|---|---|
| Argentina | Clams | 10 | 1020 | 1174 | 959 | 2001 |
| Mussels | 34 | 62,878 | 53,633 | 11,240 | 59,801 | |
| Scallops | 9 | 13,508 | 4516 | 3912 | 7816 | |
| Snails | 23 | 5993 | 5591 | 4506 | 3586 | |
| Chile | Clams | 5 | 24,692 | 1343 | 1204 | 1271 |
| Mussels | 19 | 4981 | 2275 | 1886 | 2071 | |
| Oysters | 1 | 1070 | 870 | 523 | 859 | |
| Scallops | 3 | 336 | 414 | 439 | 143 | |
| Mexico | Geoduck (w) | 1 | 4120 | 1582 | 1277 | 3680 |
| Uruguay | Clams | 3 | 1810 | 307 | 200 | 679 |
| Mussels | 7 | 6667 | 5663 | 7284 | 9223 |
| MBA | PreCOX | PCOX | LC-MS/MS | |
|---|---|---|---|---|
| PreCOX | 6.50 × 10−5 | |||
| PCOX | 7.20 × 10−10 | 1.20 × 10−5 | ||
| LC-MS/MS | 0.0007 | 0.331 | 1.50 × 10−5 |
| Mussel | Clam | Oyster | Geoduck | Snail | Scallop | Misc | Total | |
|---|---|---|---|---|---|---|---|---|
| Total tests | 84 | 41 | 6 | 73 | 32 | 16 | 2 | 254 |
| Invalid tests | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 2 |
| Disagreements a | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 4 |
| Total Pos | 81 | 24 | 1 | 51 | 27 | 13 | 2 | 199 |
| Total Neg | 2 | 16 | 5 | 22 | 2 | 2 | 0 | 49 |
| False Neg b | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 |
| False Pos c | 17 | 8 | 0 | 46 | 3 | 3 | 0 | 77 |
| Analogue | PreCOX | PCOX | LC-MS/MS |
|---|---|---|---|
| STX | y | y | y |
| GTX2 | y | y | y |
| GTX3 | y | y | y |
| GTX1 | y | y | y |
| GTX4 | y | y | y |
| GTX5 | y | y | y |
| GTX6 | y | - | y |
| NEO | y | y | y |
| C1 | y | y | y |
| C2 | y | y | y |
| C3 | y | y | y |
| C4 | y | y | y |
| dcSTX | y | y | y |
| dcGTX2 | y | y | y |
| dcGTX3 | y | y | y |
| dcGTX1 | - | - | y |
| dcGTX4 | - | - | y |
| dcNEO | y | - | y |
| doSTX | - | - | y |
| M1 | - | - | y |
| M2 | - | - | y |
| M3 | - | - | y |
| M4 | - | - | y |
| Samples | Species | Common Name | Source |
|---|---|---|---|
| Mussels | Mytilus edulis | Blue mussels | Uruguay, Argentina |
| Mytilus chilensis | Chilean blue mussels | Chile | |
| Aulacomya ater | Ribbed mussels | Argentina | |
| Brachidontes rodriguezii | Scorched mussels | Argentina | |
| Oysters | Crassostrea gigas | Pacific oysters | Chile |
| Clams | Gari solida | Gari clams | Chile |
| Venus antiqua | Venus clams | Chile, Argentina | |
| Donax hanleyanus | Wedge clams | Uruguay, Argentina | |
| Mesodesma mactroides | Yellow clams | Argentina | |
| Panopea globosa | Gulf of California geoduck clam | Mexico | |
| Scallops | Argopecten purpuratus | Scallops | Chile |
| Zygochlamys patagonica | Patagonian scallops | Argentina | |
| Aequipecten tehuelchus | Tehuelche scallops | Argentina | |
| Marine snails | Adelomelon beckii | Beck’s volute sea snail | Argentina |
| Zidona dufresnei | Angular volute sea snail | Argentina | |
| Squid | Illex argentinus | Mantle squid | Argentina |
| Miscellaneous | Exact species unknown | Limpets, shrimp heads | Argentina |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turner, A.D.; Tarnovius, S.; Hatfield, R.G.; Teixeira Alves, M.; Broadwater, M.; Van Dolah, F.; Garcia-Mendoza, E.; Medina, D.; Salhi, M.; Goya, A.B.; et al. Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America. Mar. Drugs 2020, 18, 616. https://doi.org/10.3390/md18120616
Turner AD, Tarnovius S, Hatfield RG, Teixeira Alves M, Broadwater M, Van Dolah F, Garcia-Mendoza E, Medina D, Salhi M, Goya AB, et al. Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America. Marine Drugs. 2020; 18(12):616. https://doi.org/10.3390/md18120616
Chicago/Turabian StyleTurner, Andrew D., Sophie Tarnovius, Robert G. Hatfield, Mickael Teixeira Alves, Maggie Broadwater, Frances Van Dolah, Ernesto Garcia-Mendoza, Dinorah Medina, Maria Salhi, Alejandra B. Goya, and et al. 2020. "Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America" Marine Drugs 18, no. 12: 616. https://doi.org/10.3390/md18120616
APA StyleTurner, A. D., Tarnovius, S., Hatfield, R. G., Teixeira Alves, M., Broadwater, M., Van Dolah, F., Garcia-Mendoza, E., Medina, D., Salhi, M., Goya, A. B., Barrera, F., Carrasco, D., Rubilar, I., & Suarez-Isla, B. A. (2020). Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America. Marine Drugs, 18(12), 616. https://doi.org/10.3390/md18120616