Anti-Influenza A Virus Activity of Rhamnan Sulfate from Green Algae Monostroma nitidum in Mice with Normal and Compromised Immunity

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of RS on In Vitro Replication of Enveloped and Non-Enveloped Viruses
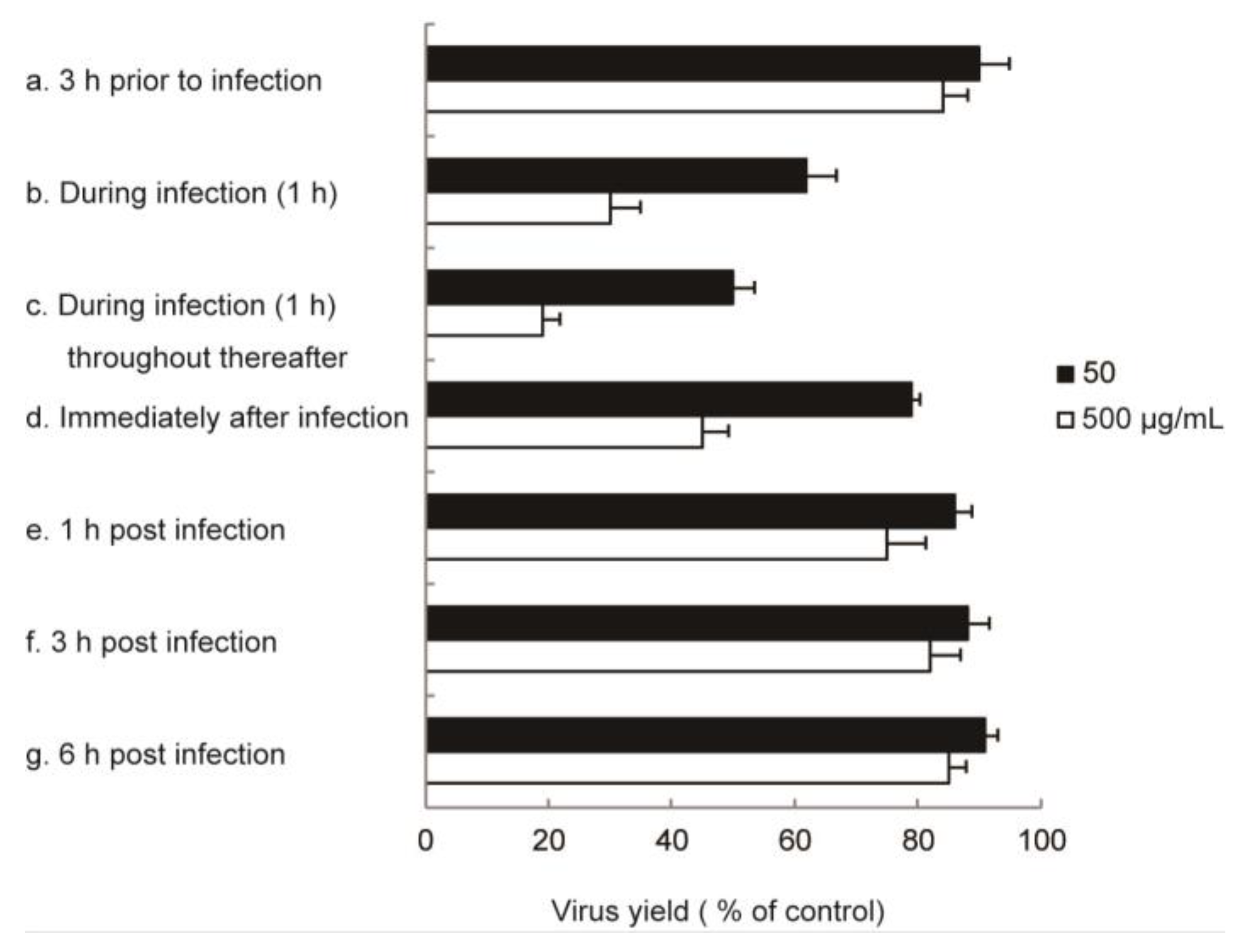
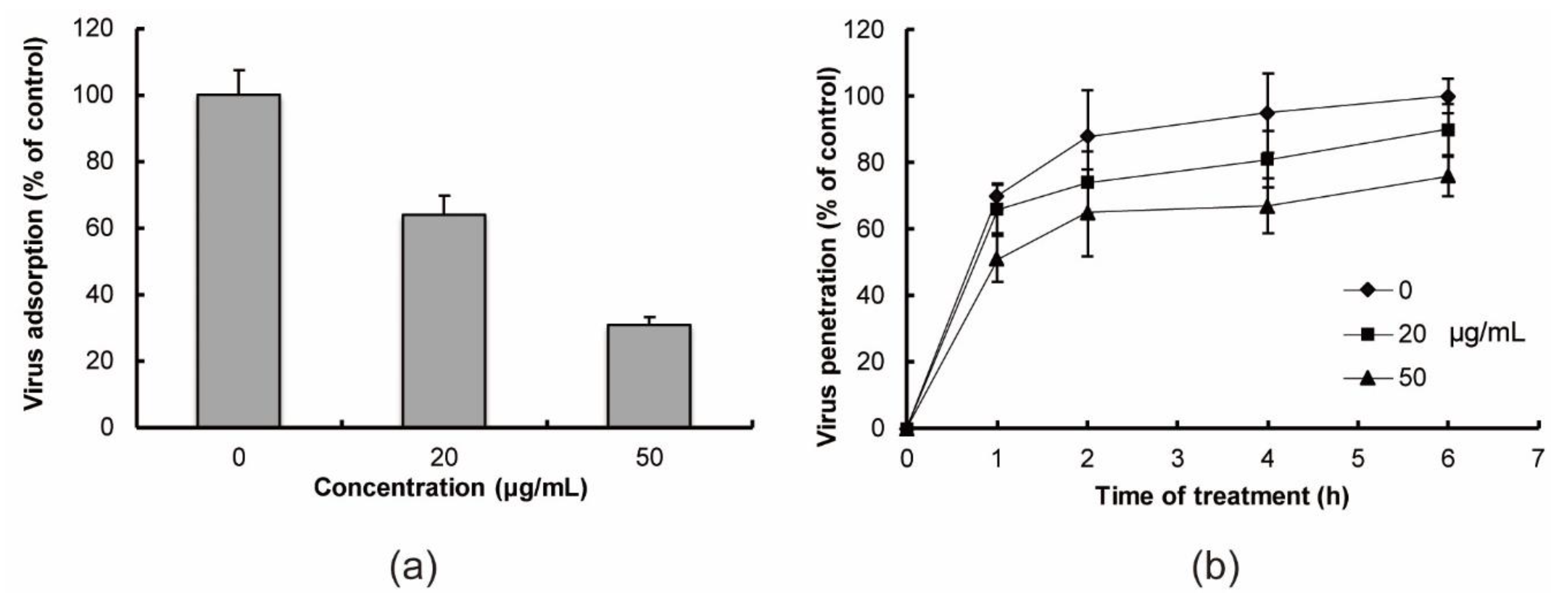
2.2. In Vitro Anti-Influenza Virus Targets
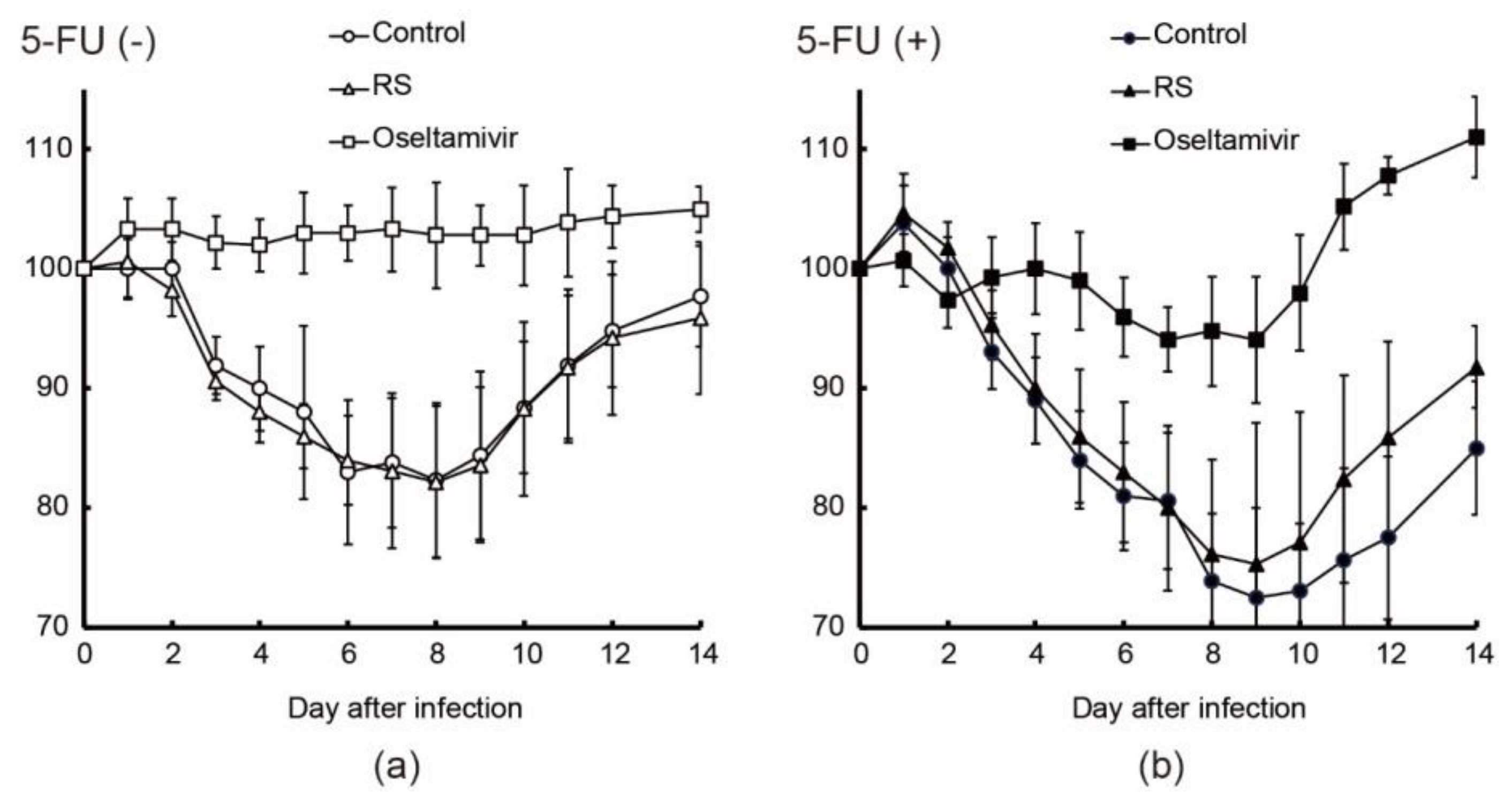
2.3. Effects of RS on Body Weight and Mortality in Immunocompetent and Immunocompromised Mice
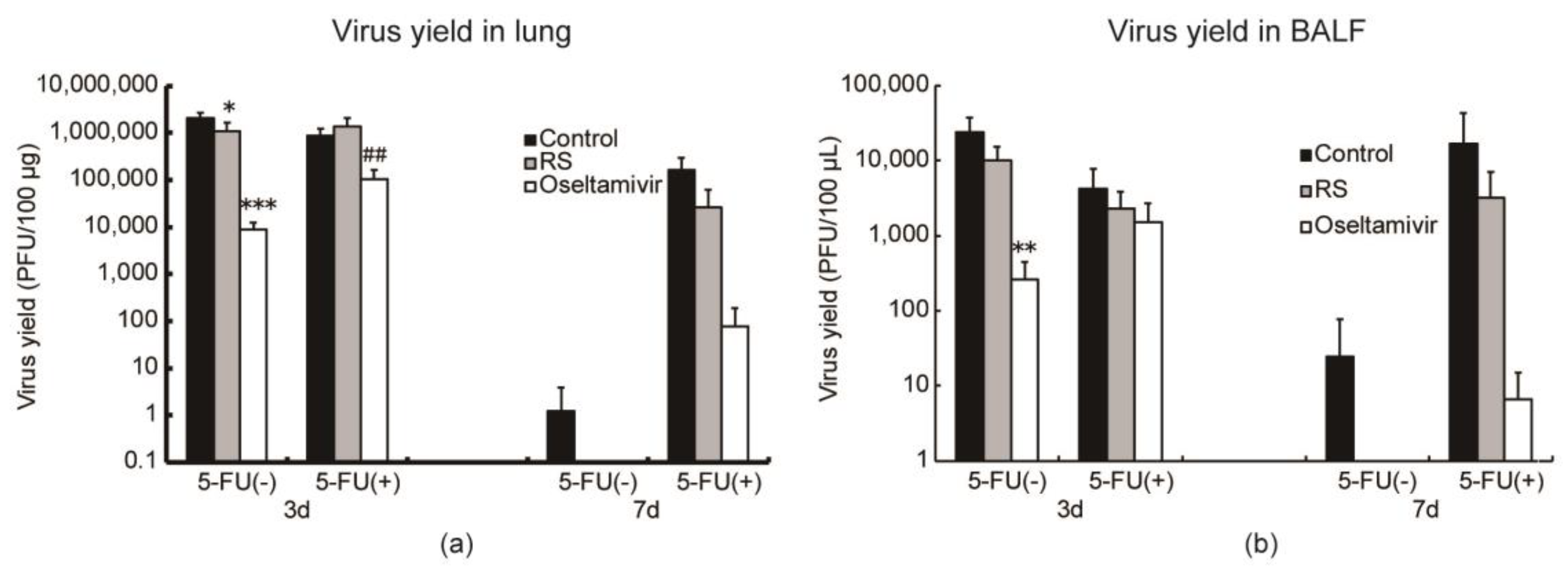
2.4. Effects of RS on Virus Replication in Mice
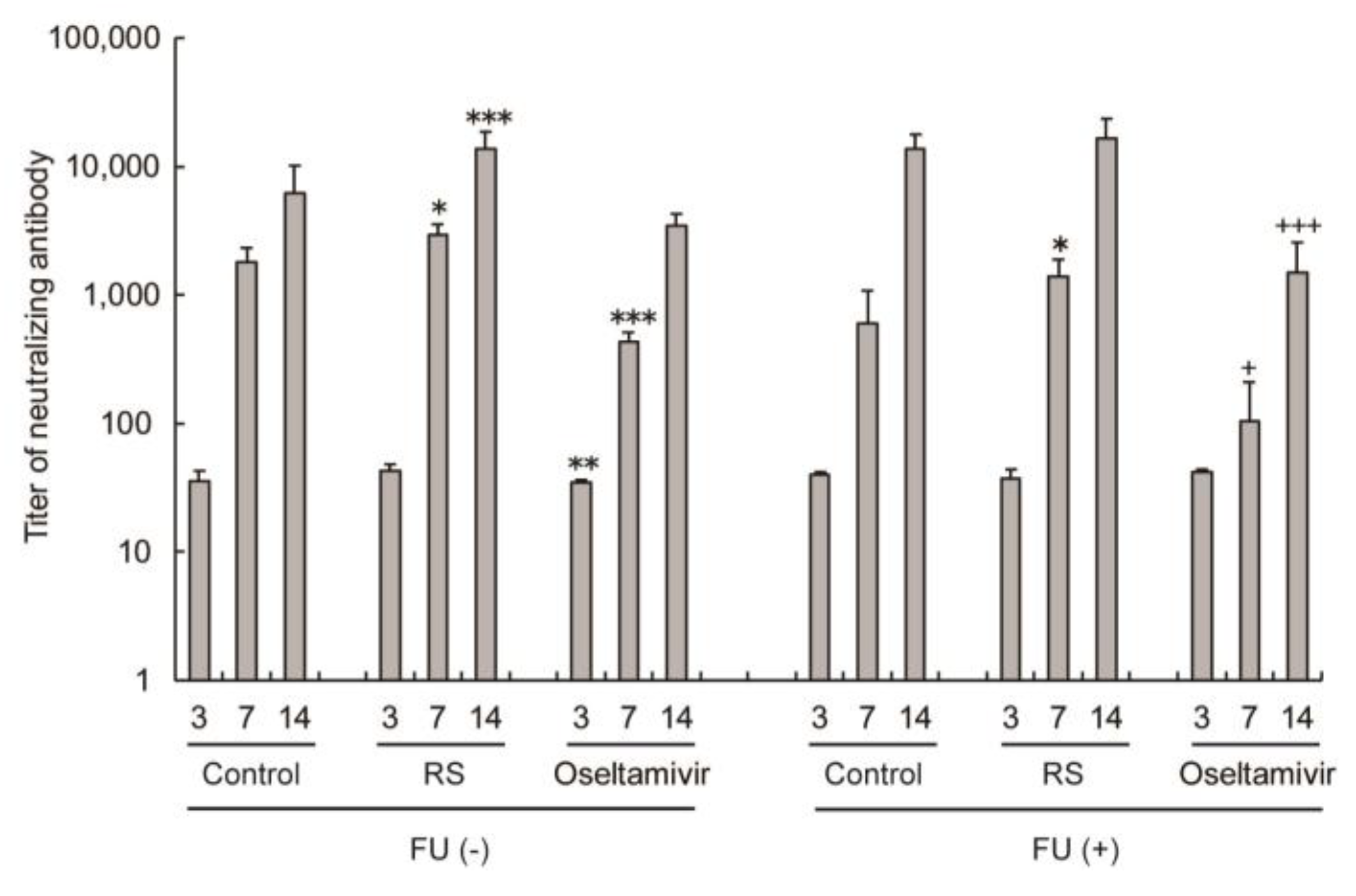
2.5. Effects of RS on Antibody Responses in Sera
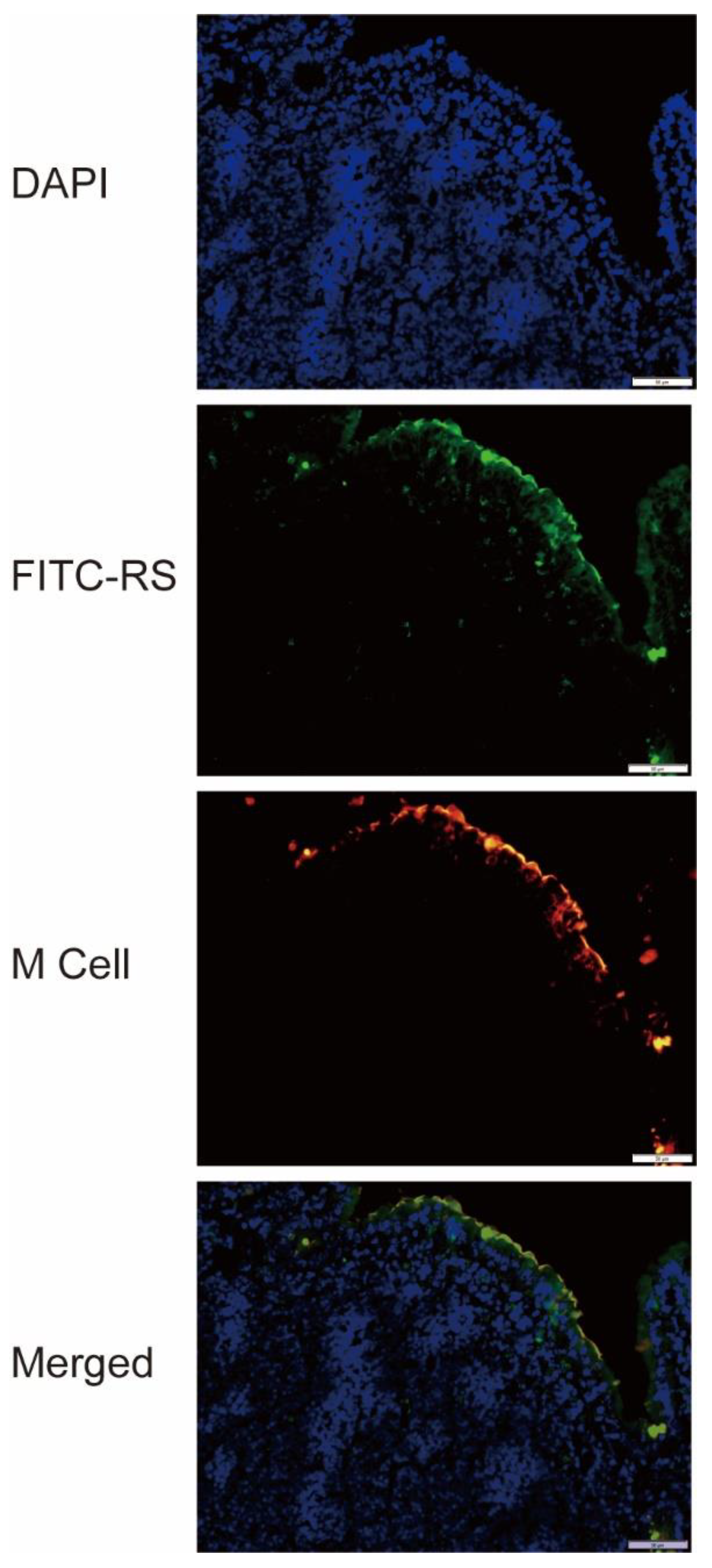
2.6. Detection of RS Uptake by M Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of RS
4.2. Cells and Viruses
4.3. In Vitro Antiviral Assays
4.4. Time-of-Addition Experiments
4.5. Virus Adsorption Assay
4.6. Virus Penetration Assay
4.7. Animals
4.8. Animal Experiments
4.9. Assay for Neutralizing Antibody
4.10. Preparation of FITC-Labeled RS
4.11. Preparation of Sections from Peyer’s Patches
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, C.; Bu, Z.; Chen, H. Avian influenza vaccines against H5N1 ‘bird flu’. Trends Biotechnol. 2014, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Chotpitayasunondh, T.; Cox, N.J.; Daniels, R.; Fry, A.M.; Gubareva, L.V.; Hayden, F.G.; Hui, D.S.; Hungnes, O.; Lackenby, A.; et al. Antiviral resistance during the 2009 influenza A H1N1 pandemic: Public health, laboratory, and clinical perspectives. Lancet Infect. Dis. 2012, 12, 240–248. [Google Scholar] [CrossRef]
- Samson, M.; Pizzorno, A.; Abed, Y.; Boivin, G. Influenza virus resistance to neuraminidase inhibitors. Antivir. Res. 2013, 98, 174–185. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Negishi, H.; Mori, M.; Mori, H.; Yamori, Y. Supplementation of elderly Japanese men and women with fucoidan from seaweed increases immune responses to seasonal influenza vaccination. J. Nutr. 2013, 143, 1794–1798. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Lee, J.B.; Nakano, T.; Hayashi, T. Anti-influenza A virus characteristics of a fucoidan from sporophyll of Undaria pinnatifida in mice with normal and compromised immunity. Microbes Infect. 2013, 15, 302–309. [Google Scholar] [CrossRef]
- Maeda, M.; Uehara, T.; Harada, N.; Sekiguchi, M.; Hiraoka, A. Heparinoid-active sulphated polysaccharides from Monostroma nitidum and their distribution in the chlorophyta. Phytochemistry 1991, 30, 3611–3614. [Google Scholar] [CrossRef]
- Zang, L.; Shimada, Y.; Tanaka, T.; Nishimura, N. Rhamnan sulphate from Monostroma nitidum attenuates hepatic steatosis by suppressing lipogenesis in a diet-induced obesity zebrafish model. J. Funct. Foods 2015, 17, 364–370. [Google Scholar] [CrossRef]
- Lee, J.B.; Koizumi, S.; Hayashi, K.; Hayashi, T. Structure of rhamnan sulfate from the green alga Monostroma nitidum and its anti-herpetic effect. Carbohydr. Polym. 2010, 81, 572–577. [Google Scholar] [CrossRef]
- Tako, M.; Yamashiro, Y.; Teruya, T.; Uechi, S. Structure-function relationship of rhamnan sulfate isolated from commercially cultured edible green seaweed, Monostroma nitidum. Am. J. Appl. Chem. 2017, 5, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Harada, N.; Maeda, M. Chemical structure of antithrombin-active rhamnan sulfate from Monostrom nitidum. Biosci. Biotechnol. Biochem. 1998, 62, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Du, P.; Liu, X.; Cao, S.; Qin, L.; He, M.; He, X.; Mao, W. Anticoagulant properties of a green algal rhamnan-type sulfated polysaccharide and its low-molecular-weight fragments prepared by mild acid degradation. Mar. Drugs 2018, 16, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, S.; Cao, S.; He, X.; Qin, L.; He, M.; Yang, Y.; Hao, J.; Mao, W. Structural characteristics and anticoagulant property in vitro and in vivo of a seaweed sulfated rhamnan. Mar. Drugs 2018, 16, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashiro, Y.; Nakamura, M.; Yogi, T.; Teruya, T.; Konishi, T.; Uechi, S.; Tako, M. Anticoagulant activity of rhamnan sulfate isolated from commercially cultured Monostroma nitidum. Int. J. Biomed. Mater. Res. 2017, 5, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mao, W.; Hou, Y.; Gao, Y.; Qi, X.; Zhao, C.; Chen, Y.; Chen, Y.; Li, N.; Wang, C. Preparation, structure and anticoagulant activity of a low molecular weight fraction produced by mild acid hydrolysis of sulfated rhamnan from Monostroma latissimum. Bioresour. Technol. 2012, 114, 414–418. [Google Scholar] [CrossRef]
- Li, N.; Liu, X.; He, X.; Wang, S.; Cao, S.; Xia, Z.; Xian, H.; Qin, L.; Mao, W. Structure and anticoagulant property of a sulfated polysaccharide isolated from the green seaweed Monostroma angicava. Carbohydr. Polym. 2017, 159, 195–206. [Google Scholar] [CrossRef]
- Wang, S.; Wang, W.; Hao, C.; Yunjia, Y.; Qin, L.; He, M.; Mao, W. Antiviral activity against enterovirus 71 of sulfated rhamnan isolated from the green alga Monostroma latissimum. Carbohydr. Polym. 2018, 200, 43–53. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hayashi, T.; Sankawa, U.; Maeda, M. Antiviral activities against HSV-1, HCMV, and HIV-1 of rhamnan sulfate from Monostroma latissimum. Planta Med. 1999, 65, 439–441. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Maeda, M.; Hayashi, T. Antiherpetic activities of sulfated polysaccharides from green algae. Planta Med. 2004, 70, 813–817. [Google Scholar] [CrossRef]
- Okamoto, T.; Akita, N.; Terasawa, M.; Hayashi, T.; Suzuki, K. Rhamnan sulfate extracted from Monostroma nitidum attenuates blood coagulation and inflammation of vascular endothelial cells. J. Nat. Med. 2019, 73, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Hugot, J.P.; Barreau, F. Peyer’s Patches: The immune sensors of the intestine. Int. J. Inflamm. 2010, 2010, 823710. [Google Scholar] [CrossRef] [Green Version]
- Lelouard, H.; Fallet, M.; de Bovis, B.; Meresse, S.; Gorvel, J.P. Peyer’s patch dendritic cells sample antigens by extending dendrites through M cell-specific transcellular pores. Gastroenterology 2012, 142, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Harrop, H.A.; Rider, C.C. Heparin and its derivatives bind to HIV-1 recombinant envelope glycoproteins, rather than to recombinant HIV-1 receptor, CD4. Glycobiology 1998, 8, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulard, M.; Lortat-Jacob, H.; Mondor, I.; Roca, G.; Wyatt, R.; Sodroski, J.; Zhao, L.; Olson, W.; Kwong, P.D.; Sattentau, Q.J. Selective interactions of polyanions with basic surfaces on human immunodeficiency virus type 1 gp120. J. Virol. 2000, 74, 1948–1960. [Google Scholar] [CrossRef] [Green Version]
- Vives, R.R.; Imberty, A.; Sattentau, Q.J.; Lortat-Jacob, H. Heparan sulfate targets the HIV-1 envelope glycoprotein gp120 coreceptor binding site. J. Biol. Chem. 2005, 280, 21353–21357. [Google Scholar] [CrossRef] [Green Version]
- Prokofjeva, M.M.; Imbs, T.I.; Shevchenko, N.M.; Spirin, P.V.; Horn, S.; Fehse, B.; Zvyagintseva, T.N.; Prassolov, V.S. Fucoidans as potential inhibitors of HIV-1. Mar. Drugs 2013, 11, 3000–3014. [Google Scholar] [CrossRef]
- Ison, M.G.; Gnann, J.W., Jr.; Nagy-Agren, S.; Treannor, J.; Paya, C.; Steigbigel, R.; Elliott, M.; Weiss, H.L.; Hayden, F.G. Safety and efficacy of nebulized zanamivir in hospitalized patients with serious influenza. Antivir. Ther. 2003, 8, 183–190. [Google Scholar]
- Leekha, S.; Zitterkopf, N.L.; Espy, M.J.; Smith, T.F.; Thompson, R.L.; Sampathkumar, P. Duration of influenza A virus shedding in hospitalized patients and implications for infection control. Infect. Control Hosp. Epidemiol. 2007, 28, 1071–1076. [Google Scholar] [CrossRef]
- Treanor, J.J.; Hayden, F.G.; Vrooman, P.S.; Barbarash, R.; Bettis, R.; Riff, D.; Singh, S.; Kinnersley, N.; Ward, P.; Mills, R.G. Efficacy and safety of the oral neuraminidase inhibitor oseltamivir in treating acute influenza: A randomized controlled trial. US oral neuraminidase study group. JAMA 2000, 283, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Memoli, M.J.; Athota, R.; Reed, S.; Czajkowski, L.; Bristol, T.; Proudfoot, K.; Hagey, R.; Voell, J.; Fiorentino, C.; Ademposi, A.; et al. The natural history of influenza infection in the severely immunocompromised vs nonimmunocompromised hosts. Clin. Infect. Dis. 2014, 58, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, K.; Shimura, T.; Suzuki, H.; Nomoto, K.; Asao, T.; Kuwano, H. Antitumor effect of a splenic injection of 5-fluorouracil on metastatic liver cancer in mice. J. Pharmacol. Exp. Ther. 2004, 308, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Ohno, H. Intestinal M cells. J. Biochem. 2016, 159, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, F.; Yan, J.; Baran, J.T.; Allendorf, D.J.; Hansen, R.D.; Ostroff, G.R.; Xing, P.X.; Cheung, N.K.; Ross, G.D. Mechanism by which orally administered beta-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models. J. Immunol. 2004, 173, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Tanioka, A.; Tanabe, K.; Hosono, A.; Kawakami, H.; Kaminogawa, S.; Tsubaki, K.; Hachimura, S. Enhancement of intestinal immune function in mice by beta-D-glucan from aureobasidium pullulans ADK-34. Scand. J. Immunol. 2013, 78, 61–68. [Google Scholar] [CrossRef]
- Kawashima, T.; Murakami, K.; Nishimura, I.; Nakano, T.; Obata, A. A sulfated polysaccharide, fucoidan, enhances the immunomodulatory effects of lactic acid bacteria. Int. J. Mol. Med. 2012, 29, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed, Cladosiphon okamuranus. Mar. Drugs 2014, 13, 48–64. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Host cell | Envelope | Cytotoxicity (CC50, μg/mL) | Antiviral activity (EC50, μg/mL) | Selectivity index (SI) (CC50/EC50) | ||
|---|---|---|---|---|---|---|---|
| A a | B b | A | B | ||||
| HSV-1 | Vero | + | 16,000 ± 640 | 6.5 ± 0.49 | 31 ± 2.8 | 2500 ± 93 | 520 ± 28 |
| HSV-2 | Vero | + | 16,000 ± 640 | 0.93 ± 0.18 | 4.7 ± 0.35 | 17,000 ± 2600 | 3400 ± 120 |
| HCMV | MRC-5 | + | 12,000 ± 920 | 1.1 ± 0.18 | 67 ± 9.9 | 11,000 ± 930 | 180 ± 13 |
| Measles virus | Vero | + | 16,000 ± 6 40 | 8.3 ± 0.57 | 4300 ± 450 | 1900 ± 210 | 3.7 ± 0.29 |
| Mumps virus | Vero | + | 16,000 ± 640 | 1.5 ± 0.19 | 51 ± 6.4 | 11,000 ± 910 | 310 ± 26 |
| IFV | MDCK | + | 17,000 ± 2000 | 41 ± 6.4 | 310 ± 42 | 410 ± 16 | 55 ± 1.1 |
| HIV | HeLa | + | 9900 ± 780 | 1.2 ± 0.15 | 1.2 ± 0.11 | 8300 ± 370 | 8300 ± 190 |
| Human coronavirus | MRC-5 | + | 12,000 ± 1900 | 0.77 ± 0.17 | 0.99 ± 0.13 | 16,000 ± 990 | 12,000 ± 350 |
| Adenovirus | HeLa | - | 4300 ± 220 | 480 ± 42 | >1000 | 9.0 ± 0.37 | <4 |
| Poliovirus | Vero | - | 16,000 ± 640 | >5000 | >5000 | <3 | <3 |
| Coxsackie virus | Vero | - | 16,000 ± 640 | 2600 ± 170 | >5000 | 6.2 ± 0.14 | <3 |
| Rhinovirus | HeLa | - | 13,000 ± 640 | 530 ± 29 | >1000 | 25 ± 0.14 | <13 |
| Sample | FU-Treat | Survived/Challenged | Mortality | Day of Death |
|---|---|---|---|---|
| Control | - | 11/11 | 0% | |
| RS | - | 11/11 | 0% | |
| Oseltamivir | - | 10/10 | 0% | |
| Control | + | 9/11 | 18% | 9, 10 d |
| RS | + | 10/10 | 0% | |
| Oseltamivir | + | 6/6 | 0% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terasawa, M.; Hayashi, K.; Lee, J.-B.; Nishiura, K.; Matsuda, K.; Hayashi, T.; Kawahara, T. Anti-Influenza A Virus Activity of Rhamnan Sulfate from Green Algae Monostroma nitidum in Mice with Normal and Compromised Immunity. Mar. Drugs 2020, 18, 254. https://doi.org/10.3390/md18050254
Terasawa M, Hayashi K, Lee J-B, Nishiura K, Matsuda K, Hayashi T, Kawahara T. Anti-Influenza A Virus Activity of Rhamnan Sulfate from Green Algae Monostroma nitidum in Mice with Normal and Compromised Immunity. Marine Drugs. 2020; 18(5):254. https://doi.org/10.3390/md18050254
Chicago/Turabian StyleTerasawa, Masahiro, Kyoko Hayashi, Jung-Bum Lee, Kaoru Nishiura, Koichi Matsuda, Toshimitsu Hayashi, and Toshio Kawahara. 2020. "Anti-Influenza A Virus Activity of Rhamnan Sulfate from Green Algae Monostroma nitidum in Mice with Normal and Compromised Immunity" Marine Drugs 18, no. 5: 254. https://doi.org/10.3390/md18050254
APA StyleTerasawa, M., Hayashi, K., Lee, J. -B., Nishiura, K., Matsuda, K., Hayashi, T., & Kawahara, T. (2020). Anti-Influenza A Virus Activity of Rhamnan Sulfate from Green Algae Monostroma nitidum in Mice with Normal and Compromised Immunity. Marine Drugs, 18(5), 254. https://doi.org/10.3390/md18050254