
Purification and Identification of Novel Xanthine Oxidase Inhibitory Peptides Derived from Round Scad (Decapterus maruadsi) Protein Hydrolysates

 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. XO Inhibitory Activity of RSHs
2.2. Molecular Weight Distribution of RSHs
2.3. Purification and Identification of XO Inhibitory Peptides
2.4. Fluorescence Quenching Studies of XO by FPSV and FPFP
2.5. Reversibility and the Type of Inhibition
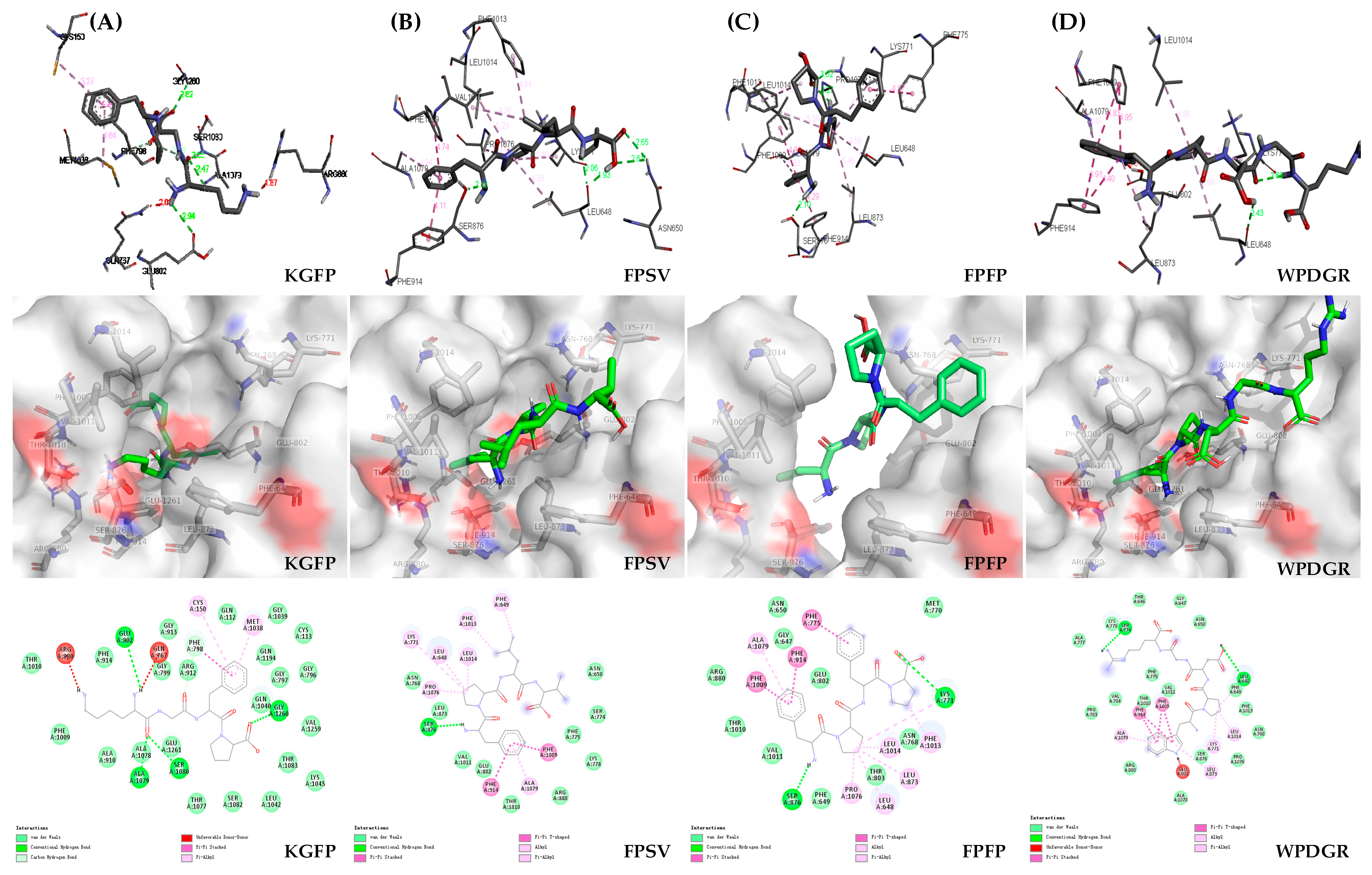
2.6. Molecular Docking and Visual Analysis
3. Materials and Methods
3.1. Materials
3.2. Preparation of Round Scad Hydrolysates (RSHs)
3.3. XO Inhibitory Activity Assay
3.4. Determination of the Degree of Hydrolysis
3.5. Molecular Weight Distribution of RSHs
3.6. Purification of RSHs with Immobilized Metal Affinity Chromatography (IMAC)
3.7. Identification and Synthesis of Peptide
3.8. Fluorescence Spectroscopy Assay
3.9. Determination of Reversibility and the Type of Inhibition
3.10. Molecular Docking and Interaction Analysis
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.; Zhou, H.N.; Zhang, R.R.; Wang, X.K.; He, S.W.; Zhang, J.B.; Guo, X.; Wang, Q.; Sun, J.Y. Anti-hyperuricemic and nephroprotective effect of geniposide in chronic hyperuricemia mice. J. Funct. Foods 2019, 61, 103355. [Google Scholar] [CrossRef]
- Prasad Sah, O.S.; Qing, Y.X. Associations Between Hyperuricemia and Chronic Kidney Disease: A Review. Nephrourol. Mon. 2015, 7, e27233. [Google Scholar] [CrossRef] [Green Version]
- Falasca, G.F. Metabolic diseases: Gout. Clin. Dermatol. 2006, 24, 498–508. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhang, G.W.; Pan, J.H.; Gong, D.M. Novel Insights into the Inhibitory Mechanism of Kaempferol on Xanthine Oxidase. J. Agric. Food Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Enroth, C.; Eger, B.T.; Okamoto, K.; Nishino, T.; Pai, E. Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: Structure based mechanism of conversion. Proc. Natl. Acad. Sci. USA 2000, 97, 10723–10728. [Google Scholar] [CrossRef] [Green Version]
- Hadizadeh, M.; Keyhani, E.; Keyhani, J.; Khodadadi, C. Functional and structural alterations induced by copper in xanthine oxidase. Acta Bioch. Biophys. Sin. 2009, 41, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Zou, B.; Zeng, H.L.; Zhang, L.N.; Chen, M.; Fu, G.M. Inhibitory effect of verbascoside on xanthine oxidase activity. Int. J. Biol. Macromol. 2016, 93, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Young, J.L.; Boswell, R.B.; Nies, A.S. Severe allopurinol hypersensitivity: Association with thiazides and prior renal compromise. Arch. Intern. Med. 1974, 134, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Gaffo, A.L.; Saag, K.G. Febuxostat: The evidence for its use in the treatment of hyperuricemia and gout. Core Evid. 2009, 4, 25–36. [Google Scholar]
- Nongonierma, A.B.; Fitzgerald, R.J. Tryptophan-containing milk protein-derived dipeptides inhibit xanthine oxidase. Peptides 2012, 37, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.Y.; Kang, X.Y.; Shi, C.C.; Li, Y.J.; Majumder, K.; Ning, Z.X.; Ren, J.Y. Moderation of hyperuricemia in rats via consuming walnut protein hydrolysate diet and identification of new antihyperuricemic peptides. Food Funct. 2018, 9, 107–116. [Google Scholar] [CrossRef]
- Li, Y.J.; Kang, X.Y.; Li, Q.Y.; Shi, C.C.; Lian, Y.Y.; Yuan, E.D.; Zhou, M.; Ren, J.Y. Anti-hyperuricemic peptides derived from bonito hydrolysates based on in vivo hyperuricemic model and in vitro xanthine oxidase inhibitory activity. Peptides 2018, 107, 45–53. [Google Scholar] [CrossRef] [PubMed]
- He, W.W.; Su, G.W.; Sun-Waterhouse, D.X.; Waterhouse, G.I.N.; Zhao, M.M.; Liu, Y. In vivo anti-hyperuricemic and xanthine oxidase inhibitory properties of tuna protein hydrolysates and its isolated fractions. Food Chem. 2019, 272, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Su, G.W.; He, W.W.; Zhao, M.M.; Waterhouse, G.I.N.; Sun-Waterhouse, D.X. Effect of different buffer systems on the xanthine oxidase inhibitory activity of tuna (Katsuwonus pelamis) protein hydrolysate. Food Res. Int. 2018, 105, 556–562. [Google Scholar] [CrossRef]
- Murota, I.; Tamai, T.; Baba, T.; Sato, R.; Hashimoto, K.; Park, E.Y.; Nakamura, Y.; Sato, K. Uric acid lowering effect by ingestion of proteolytic digest of shark cartilage and its basic fraction. J. Food Biochem. 2010, 34, 182–194. [Google Scholar] [CrossRef]
- Murota, I.; Tamai, T.; Baba, T.; Sato, N.; Park, E.Y.; Nakamura, Y.; Sato, K. Moderation of oxonate-induced hyperuricemia in rats via the ingestion of an ethanol-soluble fraction of a shark cartilage proteolytic digest. J. Funct. Foods 2012, 4, 459–464. [Google Scholar] [CrossRef]
- Murota, I.; Taguchi, S.; Sato, N.; Park, E.Y.; Nakamura, Y.; Sato, K. Identification of antihyperuricemic peptides in the proteolytic digest of shark cartilage water extract using in vivo activity-guided fractionation. J. Agric. Food Chem. 2014, 62, 2392–2397. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, Y.T.; Dong, C.M.; Li, Z.L. Spatiotemporal distribution of Decapterus maruadsi in spring and autumn in response to environmental variation in the northern South China Sea. Reg. Stud. Mar. Sci. 2021, 45, 101811. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Compositions, functional properties and antioxidative activity of protein hydrolysates prepared from round scad (Decapterus maruadsi). Food Chem. 2007, 103, 1385–1394. [Google Scholar] [CrossRef]
- Chen, H.H.; Wang, S.; Zhou, A.; Miao, J.Y.; Liu, J.Y.; Benjakul, S. A novel antioxidant peptide purified from defatted round scad (Decapterus maruadsi) protein hydrolysate extends lifespan in Caenorhabditis elegans. J. Funct. Foods 2020, 68, 103907. [Google Scholar] [CrossRef]
- Jiang, H.P.; Tong, T.Z.; Sun, J.H.; Xu, Y.J.; Zhao, Z.X.; Liao, D.K. Purification and characterization of antioxidative peptides from round scad (Decapterus maruadsi) muscle protein hydrolysate. Food Chem. 2014, 154, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Su, G.W.; Zhao, T.T.; Wang, S.G.; Sun, B.G.; Zheng, L.; Zhao, M.M. The memory improving effects of round scad (Decapterus maruadsi) hydrolysates on sleep deprivation-induced memory deficits in rats via antioxidant and neurotrophic pathways. Food Funct. 2019, 10, 7733–7744. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yang, X.Q.; Wang, T.T.; Li, L.H.; Wu, Y.Y.; Zhou, Y.; You, L.J. Purification and identification of antioxidant peptides from round scad (Decapterus maruadsi) hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. Toxicol. 2019, 135, 110882. [Google Scholar] [CrossRef]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Harnedy, P.A.; Li, B.F.; Hou, H.; Zhang, Z.H.; Zhao, X.; FitzGerald, R.J. Food protein-derived chelating peptides: Biofunctional ingredients for dietary mineral bioavailability enhancement. Trends Food Sci. Technol. 2014, 37, 92–105. [Google Scholar] [CrossRef]
- Liu, P.R.; Lan, X.D.; Yaseed, M.; Chai, K.G.; Zhou, L.Q.; Sun, J.H.; Lan, P.; Tong, Z.F.; Liao, D.K. Immobilized metal affinity chromatography matrix modified by poly (ethylene glycol) methyl ether for purification of angiotensin I-converting enzyme inhibitory peptide from casein hydrolysate. J. Chromatogr. B 2020, 1143, 122042. [Google Scholar] [CrossRef]
- Peng, W.; Ding, F.; Jiang, Y.T.; Sun, Y.; Peng, Y.K. Evaluation of the biointeraction of colorant flavazin with human serum albumin: Insights from multiple spectroscopic studies, in silico docking and molecular dynamics simulation. Food Funct. 2014, 5, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; Fitzgerald, R.J. Inhibition of dipeptidyl peptidase IV and xanthine oxidase by amino acids and dipeptides. Food Chem. 2013, 141, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Zeng, N.; Zhang, G.W.; Hu, X.; Pan, J.H.; Gong, D.M. Mechanism of fisetin suppressing superoxide anion and xanthine oxidase activity. J. Funct. Foods 2019, 58, 1–10. [Google Scholar] [CrossRef]
- Yang, J.; Chang, R.R.; Ge, S.J.; Zhao, M.; Liang, C.F.; Xiong, L.; Sun, Q.J. The inhibition effect of starch nanoparticles on tyrosinase activity and its mechanism. Food Funct. 2016, 7, 4804–4815. [Google Scholar] [CrossRef]
- Zeng, N.; Zhang, G.W.; Hu, X.; Pan, J.H.; Zhou, Z.S.; Gong, D.M. Inhibition mechanism of baicalein and baicalin on xanthine oxidase and their synergistic effect with allopurinol. J. Funct. Foods 2018, 50, 172–182. [Google Scholar] [CrossRef]
- Tavani, C.; Bianchi, L.; Palma, A.D.; Passeri, G.I.; Punzi, G.; Pierri, C.L.; Lovece, A.; Cavalluzzi, M.M.; Franchini, C.; Lentini, G.; et al. Nitro-substituted tetrahydroindolizines and homologs: Design, kinetics, and mechanism of α-glucosidase inhibition. Bioorg. Med. Chem. Lett. 2017, 27, 3980–3986. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Kwon, S.H.; Hwang, S.H.; Kang, Y.H.; Lee, J.Y.; Lim, S.S. Competitive binding experiments can reduce the false positive results of affinity-based ultrafiltration-HPLC: A case study for identification of potent xanthine oxidase inhibitors from Perilla frutescens extract. J. Chromatogr. B 2017, 1048, 30–37. [Google Scholar] [CrossRef]
- Nishino, T.; Okamoto, K.; Eger, B.T.; Pai, E.F.; Nishino, T. Mammalian xanthine oxidoreductase-mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS J. 2008, 275, 3278–3289. [Google Scholar] [CrossRef]
- Okamoto, K.; Kusano, T.; Nishino, T. Chemical nature and reaction mechanisms of the molybdenum cofactor of xanthine oxidoreductase. Curr. Pharm. Des. 2013, 19, 2606–2614. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.Y.; Shi, C.C.; Wang, M.; Zhou, M.; Liang, M.; Zhang, T.; Yuan, E.D.; Wang, Z.; Yao, M.J.; Ren, J.Y. Tryptophan residue enhances in vitro walnut protein-derived peptides exerting xanthine oxidase inhibition and antioxidant activities. J. Funct. Foods 2019, 53, 276–285. [Google Scholar] [CrossRef]
- Jayaraj, P.; Mathew, B.; Parimaladevi, B.; Ramani, V.A.; Govindarajan, R. Isolation of a bioactive flavonoid from Spilanthes calva D.C. in vitro xanthine oxidase assay and in silico study. Biomed. Prev. Nutr. 2014, 4, 481–484. [Google Scholar] [CrossRef]
- Masuoka, N.; Nihei, K.I.; Maeta, A.; Yamagiwa, Y.; Kubo, I. Inhibitory effects of cardols and related compounds on superoxide anion generation by xanthine oxidase. Food Chem. 2015, 166, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.M.; He, L.C.; Liang, Y.H.; Yue, L.L.; Peng, W.M.; Jin, G.F.; Ma, M.H. Preparation process optimization of pig bone collagen peptide-calcium chelate using response surface methodology and its structural characterization and stability analysis. Food Chem. 2019, 284, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.; Zhang, G.N.; Tian, Y.; Kong, F.B.; Xiong, S.; Zhao, S.B.; Jia, D.; Manyande, A.; Du, H.Y. Identification of novel antioxidant peptides from snakehead (Channa argus) soup generated during gastrointestinal digestion and insights into the anti-oxidation mechanisms. Food Chem. 2021, 337, 12791. [Google Scholar] [CrossRef]
- Lv, Y.; Liu, Q.; Bao, X.L.; Tang, W.X.; Yang, B.C.; Guo, S.T. Identification and characteristics of iron-chelating peptides from soybean protein hydrolysates using IMAC-Fe3+. J. Agric. Food Chem. 2009, 57, 4593–4597. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | MW (Da) | Structure Formula | XO Inhibitory Activity (%) |
|---|---|---|---|
 | 447.53 |  | 5.43 ± 0.20 c |
 | 448.51 |  | 22.61 ± 1.81 a |
 | 506.59 |  | 20.09 ± 2.41 a |
 | 629.66 |  | 16.21 ± 0.78 b |
| Peptides | T (K) | Ksv (×104 M−1) | Kq (×1012 M−1 s−1) | R2 |
|---|---|---|---|---|
| FPSV | 298 | 0.0542 | 0.0542 | 0.9898 |
| FPFP | 298 | 0.5608 | 0.5608 | 0.9999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Zhou, Y.; Zhou, S.; Chen, S.; Wu, Y.; Li, L.; Yang, X. Purification and Identification of Novel Xanthine Oxidase Inhibitory Peptides Derived from Round Scad (Decapterus maruadsi) Protein Hydrolysates. Mar. Drugs 2021, 19, 538. https://doi.org/10.3390/md19100538
Hu X, Zhou Y, Zhou S, Chen S, Wu Y, Li L, Yang X. Purification and Identification of Novel Xanthine Oxidase Inhibitory Peptides Derived from Round Scad (Decapterus maruadsi) Protein Hydrolysates. Marine Drugs. 2021; 19(10):538. https://doi.org/10.3390/md19100538
Chicago/Turabian StyleHu, Xiao, Ya Zhou, Shaobo Zhou, Shengjun Chen, Yanyan Wu, Laihao Li, and Xianqing Yang. 2021. "Purification and Identification of Novel Xanthine Oxidase Inhibitory Peptides Derived from Round Scad (Decapterus maruadsi) Protein Hydrolysates" Marine Drugs 19, no. 10: 538. https://doi.org/10.3390/md19100538
APA StyleHu, X., Zhou, Y., Zhou, S., Chen, S., Wu, Y., Li, L., & Yang, X. (2021). Purification and Identification of Novel Xanthine Oxidase Inhibitory Peptides Derived from Round Scad (Decapterus maruadsi) Protein Hydrolysates. Marine Drugs, 19(10), 538. https://doi.org/10.3390/md19100538