
ARTP Mutagenesis of Schizochytrium sp. PKU#Mn4 and Clethodim-Based Mutant Screening for Enhanced Docosahexaenoic Acid Accumulation



 ,
,  and
and 
Abstract
:1. Introduction
2. Results
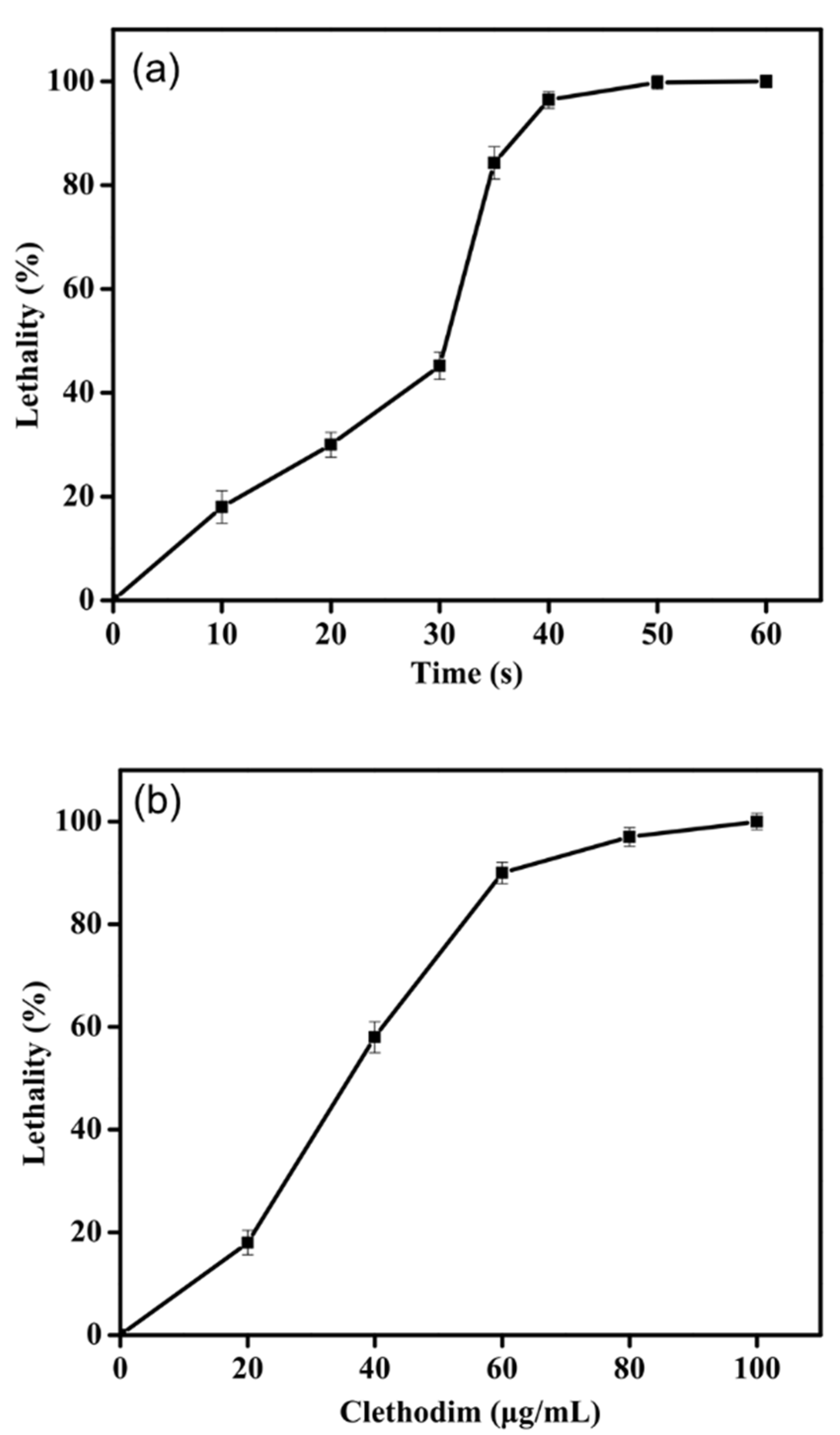
2.1. Optimal ARTP Exposure Time and Clethodim Concentration
2.2. Mutant Screening
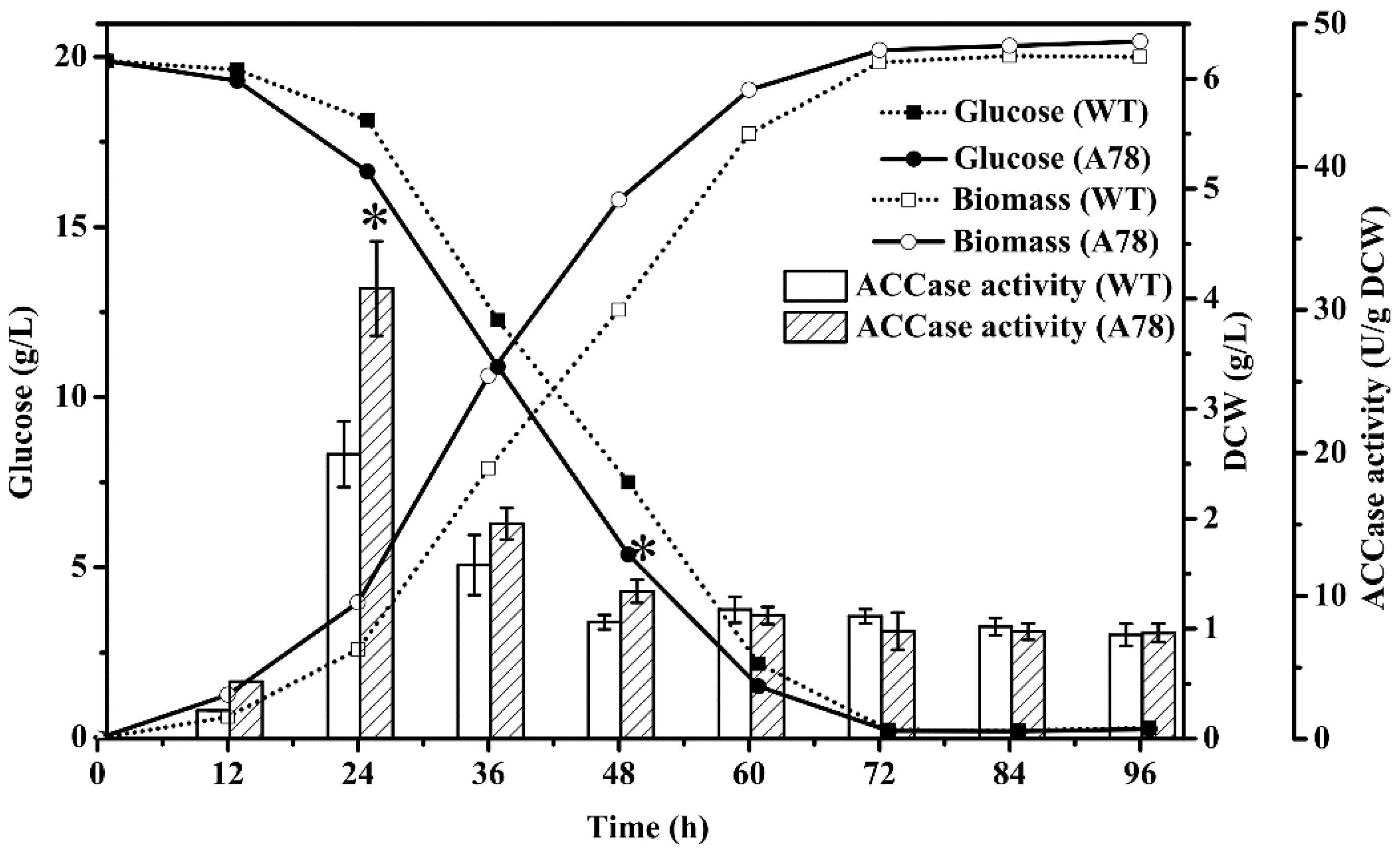
2.3. Dynamics of Growth, Glucose Consumption and ACCase Activity in Mutant A78
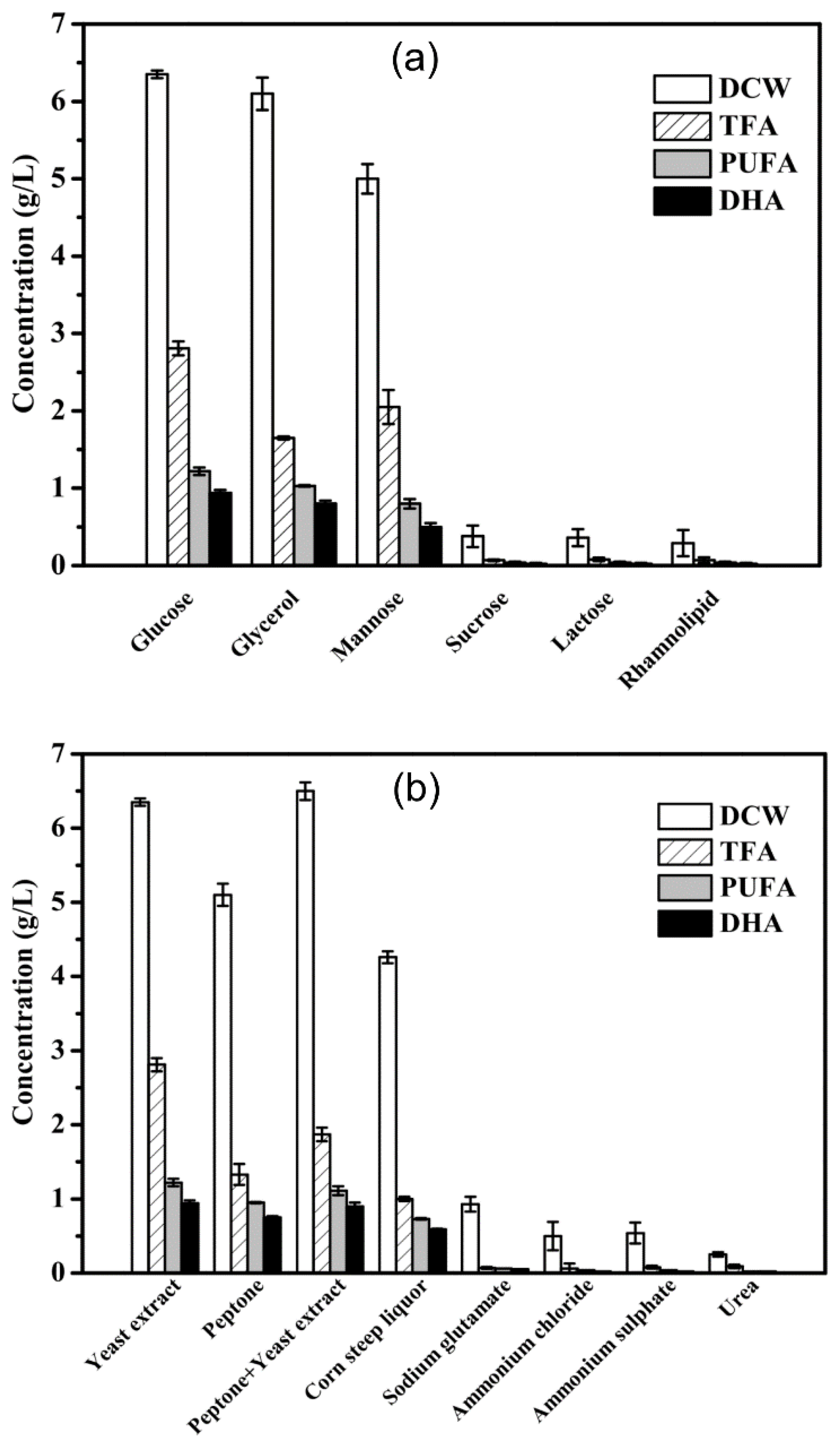
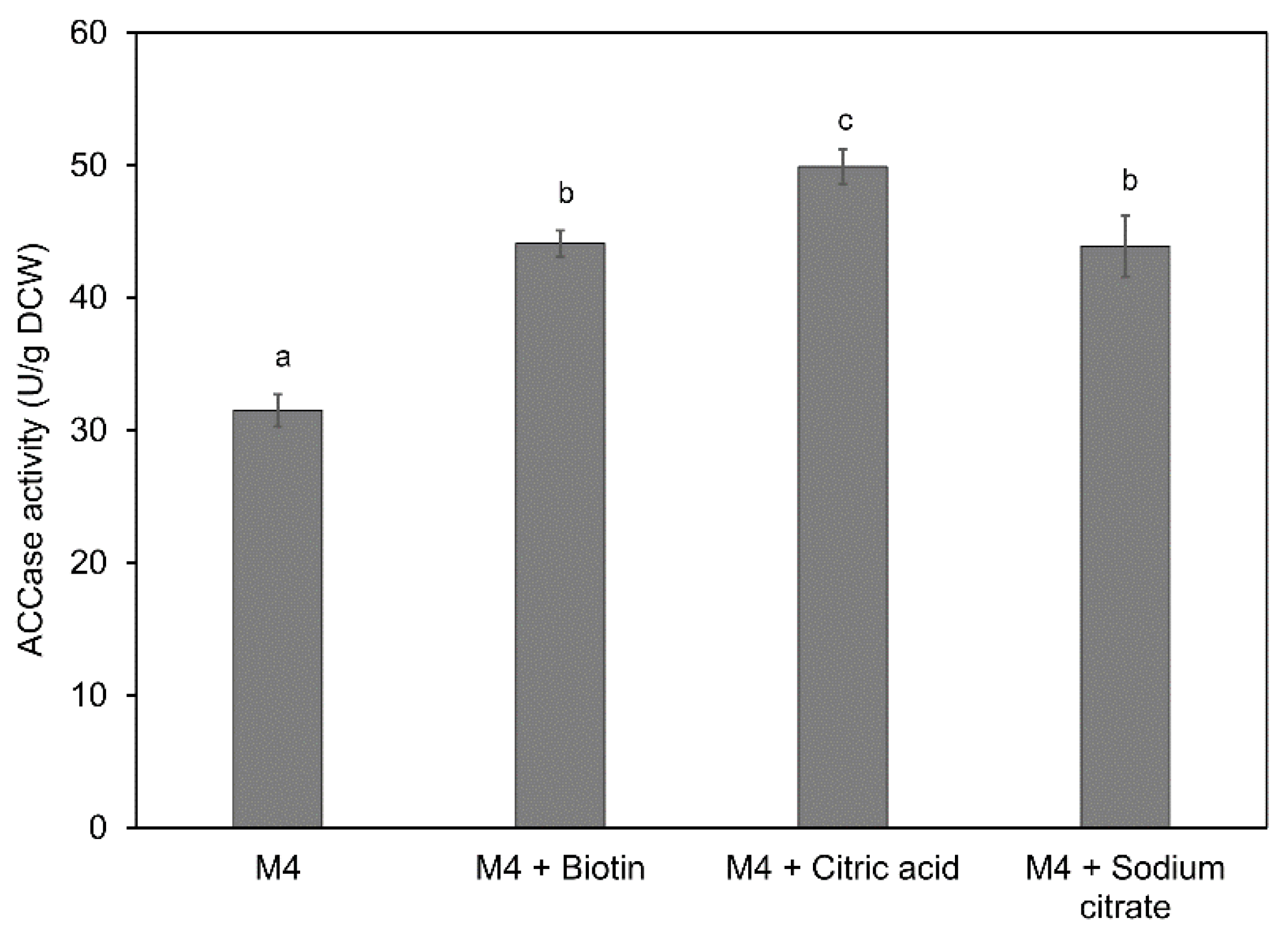
2.4. Effects of Media Components on DHA Accumulation in Mutant A78
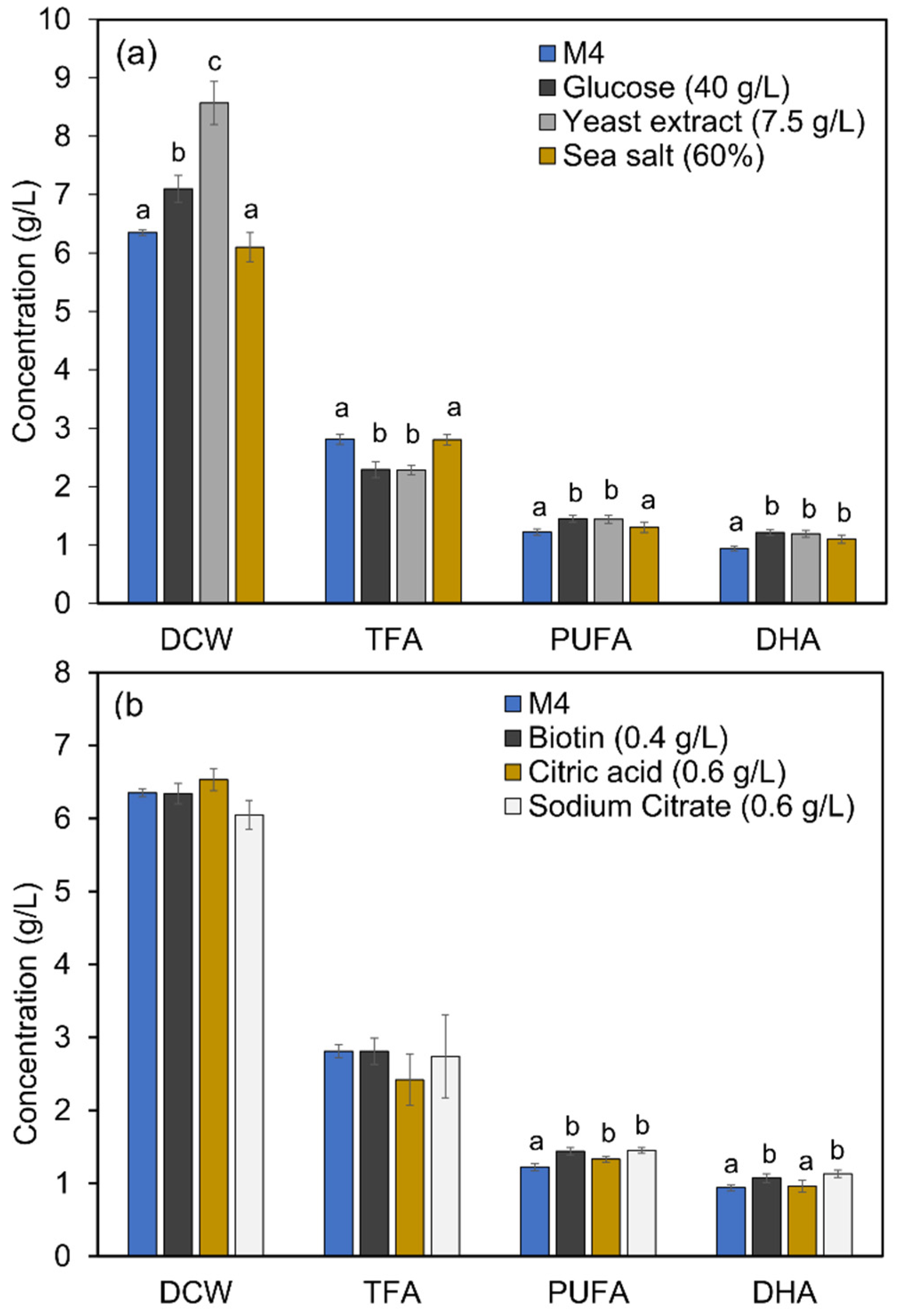
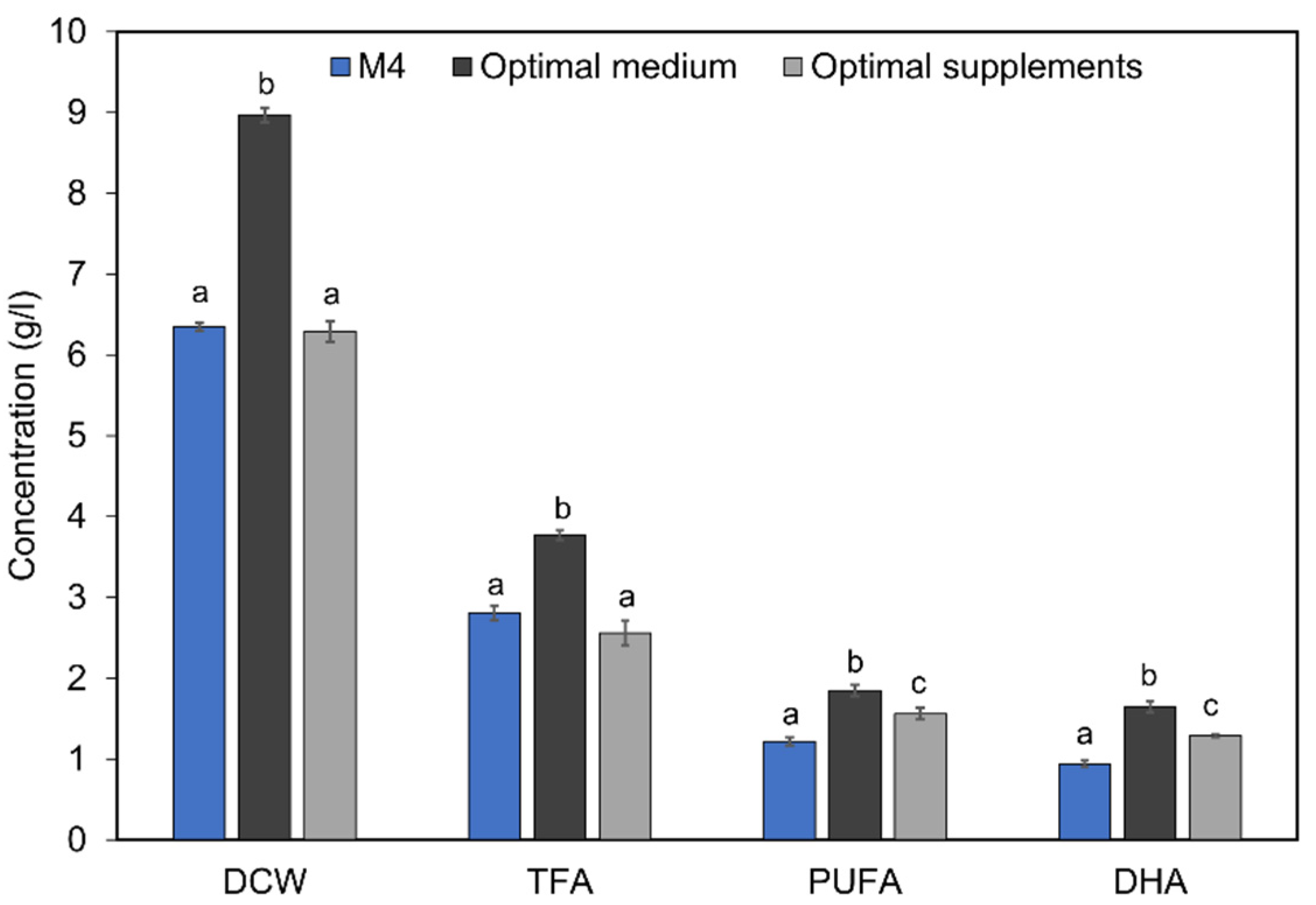
2.5. Improved DHA Accumulation in Mutant A78 through Media Optimization
3. Discussion
4. Materials and Methods
4.1. Strain and Seed Culture
4.2. Quantification of DCW, TFA and ACCase Activity
4.3. Sensitivity Assay of Schizochytrium sp. PKU#Mn4 to Clethodim
4.4. ARTP Mutagenesis and Clethodim-Based Screening
4.5. Genetic Stability of the Mutants
4.6. Medium Optimization
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Horrocks, L.A.; Yeo, Y.K. Health Benefits of Docosahexaenoic Acid (DHA). Pharmacol. Res. 1999, 40, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.A.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [Green Version]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Sargent, J.R. Fish oils and human diet. Br. J. Nutr. 1997, 78 (Suppl. 1), S5–S13. [Google Scholar] [CrossRef]
- Lenihan-Geels, G.; Bishop, K.S.; Ferguson, L.R. Alternative sources of omega-3 fats: Can we find a sustainable substitute for fish? Nutrients 2013, 5, 1301–1315. [Google Scholar] [CrossRef]
- Singh, P.; Liu, Y.; Li, L.; Wang, G. Ecological dynamics and biotechnological implications of thraustochytrids from marine habitats. Appl. Microbiol. Biotechnol. 2014, 98, 5789–5805. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ye, H.; Xie, Y.; He, Y.; Sen, B.; Wang, G. Culturable Diversity and Lipid Production Profile of Labyrinthulomycete Protists Isolated from Coastal Mangrove Habitats of China. Mar. Drugs 2019, 17, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Li, Y.; Ji, H.; Zhang, X.; Hu, X.; Zhu, G.; Lu, H.; Zhong, M. Optimization of cultivation conditions for DHA production by Schizochytrium sp. 20888. Microbiol. China 2016, 43, 1199–1209. [Google Scholar]
- Jiang, X.; Zhang, J.; Zhao, J.; Gao, Z.Q.; Zhang, C.Z.; Chen, M. Regulation of lipid accumulation in Schizochytrium sp ATCC 20888 in response to different nitrogen sources. Eur. J. Lipid Sci. Tech. 2017, 119, 8. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, H.; Sen, B.; Xie, Y.; He, Y.; Park, S.; Wang, G. Improved production of docosahexaenoic acid in batch fermentation by newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. through bioprocess optimization. Synth. Sys. Biotechnol. 2018, 3, 121–129. [Google Scholar] [CrossRef]
- Ye, H.; He, Y.; Xie, Y.; Sen, B.; Wang, G. Fed-batch fermentation of mixed carbon source significantly enhances the production of docosahexaenoic acid in Thraustochytriidae sp. PKU#Mn16 by differentially regulating fatty acids biosynthetic pathways. Bioresour. Technol. 2020, 297, 122402. [Google Scholar]
- Chen, X.; He, Y.; Ye, H.; Xie, Y.; Sen, B.; Jiao, N.; Wang, G. Different carbon and nitrogen sources regulated docosahexaenoic acid (DHA) production of Thraustochytriidae sp. PKU#SW8 through a fully functional polyunsaturated fatty acid (PUFA) synthase gene (pfaB). Bioresour. Technol. 2020, 318, 124273. [Google Scholar]
- Yin, F.W.; Zhang, Y.T.; Jiang, J.Y.; Guo, D.S.; Gao, S.; Gao, Z. Efficient docosahexaenoic acid production by Schizochytrium sp. via a two-phase pH control strategy using ammonia and citric acid as pH regulators. Process Biochem. 2019, 77, 1–7. [Google Scholar] [CrossRef]
- Guo, D.S.; Ji, X.J.; Ren, L.J.; Li, G.L.; Yin, F.W.; Huang, H. Development of a real-time bioprocess monitoring method for docosahexaenoic acid production by Schizochytrium sp. Bioresour. Technol. 2016, 216, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.S.; Tong, L.L.; Ji, X.J.; Ren, L.J.; Ding, Q.Q. Development of a strategy to improve the stability of culture environment for docosahexaenoic acid fermentation by Schizochytrium sp. Appl. Biochem. Biotechnol. 2020, 192, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.W.; Zhu, S.Y.; Guo, D.S.; Ren, L.J.; Ji, X.J.; Huang, H.; Gao, Z. Development of a strategy for the production of docosahexaenoic acid by Schizochytrium sp from cane molasses and algae-residue. Bioresour. Technol. 2019, 271, 118–124. [Google Scholar] [CrossRef]
- Liang, Y.-M.; Liu, Y.; Li, J.-J.; Maurycy, D.; Cheng, J.J. Screening of high docosahexanoic acid yield mutants of Aurantiochytrium sp. PKU#SW7. Microbiol. China 2016, 43, 457–464. [Google Scholar]
- Fang, M.; Jin, L.; Zhang, C.; Tan, Y.; Jiang, P.; Ge, N.; Heping, L.; Xing, X. Rapid mutation of Spirulina platensis by a new mutagenesis system of atmospheric and room temperature plasmas (ARTP) and generation of a mutant library with diverse phenotypes. PLoS ONE 2013, 8, e77046. [Google Scholar] [CrossRef] [PubMed]
- Hlavova, M.; Turoczy, Z.; Bisova, K. Improving microalgae for biotechnology–from genetics to synthetic biology. Biotechnol. Adv. 2015, 33, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Tanadul, O.U.; Noochanong, W.; Jirakranwong, P.; Chanprame, S. EMS-induced mutation followed by quizalofop-screening increased lipid productivity in Chlorella sp. Bioprocess Biosyst. Eng. 2018, 41, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.S.; Li, S.; Zhang, L.L.; Zhang, C.; Han, Y.J.; Gao, G.; Sun, X.; Gong, X. Mutagenesis of Rhodobacter sphaeroides using atmospheric and room temperature plasma treatment for efficient production of coenzyme Q10. J. Biosci. Bioeng. 2019, 127, 698–702. [Google Scholar] [CrossRef]
- Ottenheim, C.; Nawrath, M.; Wu, J.C. Microbial mutagenesis by atmospheric and room-temperature plasma (ARTP): The latest development. Bioresour. Bioprocess. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, M.; Wang, S.; Fang, Y.; Wang, D.; Ren, W.; Zhao, G. The atmospheric and room-temperature plasma (ARTP) method on the dextranase activity and structure. Int. J. Biol. Macromol. 2014, 70, 284–291. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, H.; Chen, X.; Sun, B.; Du, G.; Zhou, Z.; Song, J.; Fan, Y.; Shen, W. Significantly improving the yield of recombinant proteins in Bacillus subtilis by a novel powerful mutagenesis tool (ARTP): Alkaline alpha-amylase as a case study. Protein Expr. Purif. 2015, 114, 82–88. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, L.-R.; Wei, L.; Li, H.-G.; Wang, L.; Zhou, Y.; Yu, X.-B. Mutation breeding of lycopene-producing strain Blakeslea Trispora by a novel atmospheric and room temperature plasma (ARTP). Appl. Biochem. Biotechnol. 2014, 174, 452–460. [Google Scholar]
- Choi, J.-I.; Yoon, M.; Joe, M.; Park, H.; Lee, S.G.; Han, S.J.; Lee, P.C. Development of microalga Scenedesmus dimorphus mutant with higher lipid content by radiation breeding. Bioprocess Biosyst. Eng. 2014, 37, 2437–2444. [Google Scholar] [CrossRef]
- Yang, M.; An, Y.; Zabed, H.M.; Guo, Q.; Yun, J.; Zhang, G.; Awad, F.N.; Sun, W.; Qi, X. Random mutagenesis of Clostridium butyricum strain and optimization of biosynthesis process for enhanced production of 1,3-propanediol. Bioresour. Technol. 2019, 284, 188–196. [Google Scholar] [CrossRef]
- Ban, S.; Lin, W.; Luo, Z.; Luo, J. Improving hydrogen production of Chlamydomonas reinhardtii by reducing chlorophyll content via atmospheric and room temperature plasma. Bioresour. Technol. 2019, 275, 425–429. [Google Scholar] [CrossRef]
- Shu, L.; Si, X.G.; Yang, X.D.; Ma, W.Y.; Sun, J.L.; Zhang, J.; Xue, X.L.; Wang, D.P.; Gao, Q. Enhancement of acid protease activity of Aspergillus oryzae using atmospheric and room temperature plasma. Front. Microbiol. 2020, 11, 01418. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, Y.; Li, C.; Yang, H.; Wang, W. Enhancement of Schizochytrium DHA synthesis by plasma mutagenesis aided with malonic acid and zeocin screening. Appl. Microbiol. Biotechnol. 2018, 102, 2351–2361. [Google Scholar] [CrossRef]
- Li, X.; Liu, R.; Li, J.; Chang, M.; Liu, Y.; Jin, Q.; Wang, X. Enhanced arachidonic acid production from Mortierella alpina combining atmospheric and room temperature plasma (ARTP) and diethyl sulfate treatments. Bioresour. Technol. 2015, 177, 134–140. [Google Scholar] [CrossRef]
- Gao, M.; Song, X.; Feng, Y.; Li, W.; Cui, Q. Isolation and characterization of Aurantiochytrium species: High docosahexaenoic acid (DHA) production by the newly isolated microalga, Aurantiochytrium sp SD116. J. Oleo Sci. 2013, 62, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Diao, J.; Song, X.; Cui, J.; Liu, L.; Shi, M.; Wang, F.; Zhang, W. Rewiring metabolic network by chemical modulator based laboratory evolution doubles lipid production in Crypthecodinium cohnii. Metab. Eng. 2019, 51, 88–98. [Google Scholar] [CrossRef]
- Ma, C.; Ren, H.; Xing, D.; Xie, G.; Ren, N.; Liu, B. Mechanistic understanding towards the effective lipid production of a microalgal mutant strain Scenedesmus sp. Z-4 by the whole genome bioinformation. J. Hazard. Mater. 2019, 375, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Huerlimann, R.; Steinig, E.J.; Loxton, H.; Zenger, K.R.; Jerry, D.R.; Heimann, K. The effect of nitrogen limitation on acetyl-CoA carboxylase expression and fatty acid content in Chromera velia and Isochrysis aff. galbana (TISO). Gene 2014, 543, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Uppalapati, S.R.; Alamsjah, M.A.; Fujita, Y. Isolation of quizalofop-resistant mutants of Nannochloropsis oculata (Eustigmatophyceae) with high eicosapentaenoic acid following N-methyl-N-nitrosourea-induced random mutagenesis. J. Appl. Phycol. 2004, 16, 135–144. [Google Scholar] [CrossRef]
- Davis, M.S.; Solbiati, J.; Cronan, J.E. Overproduction of acetyl-CoA carboxylase activity increases the rate of fatty acid biosynthesis in Escherichia coli. J. Biol. Chem. 2000, 275, 28593–28598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Pei, G.; Diao, J.; Chen, Z.; Liu, L.; Chen, L.; Zhang, W. Screening and transcriptomic analysis of Crypthecodinium cohnii mutants with high growth and lipid content using the acetyl-CoA carboxylase inhibitor sethoxydim. Appl. Microbiol. Biotechnol. 2017, 101, 6179–6191. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Collavo, A.; Zheng, M.Q.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of acetyl-coenzyme a carboxylase mutations in resistant Lolium populations: Evaluation using clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef]
- Laroussi, M. Low temperature plasma-based sterilization: Overview and state-of-the-art. Plasma Process. Polym. 2005, 2, 391–400. [Google Scholar] [CrossRef]
- Cao, S.; Zhou, X.; Jin, W.; Wang, F.; Tu, R.; Han, S.; Chen, H.; Chen, C.; Xie, G.-J.; Ma, F. Improving of lipid productivity of the oleaginous microalgae Chlorella pyrenoidosa via atmospheric and room temperature plasma (ARTP). Bioresour. Technol. 2017, 244, 1400–1406. [Google Scholar] [CrossRef]
- An, J.; Gao, F.; Ma, Q.; Xiang, Y.; Ren, D.; Lu, J. Screening for enhanced astaxanthin accumulation among Spirulina platensis mutants generated by atmospheric and room temperature plasmas. Algal Res. 2017, 25, 464–472. [Google Scholar] [CrossRef]
- Liu, J.; Guo, T.; Luo, Y.; Chai, X.; Wu, J.; Zhao, W.; Jiao, P.; Luo, F.; Lin, Q. Enhancement of Monascus pigment productivity via a simultaneous fermentation process and separation system using immobilized-cell fermentation. Bioresour. Technol. 2019, 272, 552–560. [Google Scholar] [CrossRef]
- Wang, B.; Tan, F.; Yu, F.; Li, H.; Zhang, M. Efficient biorefinery of whole cassava for citrate production using Aspergillus niger mutated by atmospheric and room temperature plasma and enhanced co-saccharification strategy. J. Sci. Food Agric. 2021, 101, 4613–4620. [Google Scholar] [CrossRef] [PubMed]
- Bernaerts, T.M.M.; Gheysen, L.; Kyomugasho, C.; Jamsazzadeh Kermani, Z.; Vandionant, S.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Comparison of microalgal biomasses as functional food ingredients: Focus on the composition of cell wall related polysaccharides. Algal Res. 2018, 32, 150–161. [Google Scholar] [CrossRef]
- Darley, W.M.; Porter, D.; Fuller, M.S. Cell wall composition and synthesis via Golgi-directed scale formation in the marine eucaryote, Schizochytrium aggregatum, with a note on Thraustochytrium sp. Arch. Mikrobiol. 1973, 90, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Bahnweg, G.; Jckle, I. A new approach to taxonomy of the Thraustochytriales and Labyrinthulales. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 131–140. [Google Scholar]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Li, H.P.; Wang, L.Y.; Wang, S.; Zhao, H.X.; Sun, W.T.; Xing, X.H.; Bao, C.Y. Genetic effects of radio-frequency, atmospheric-pressure glow discharges with helium. Appl. Phys. Lett. 2008, 92, 3. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhao, B.; Sun, M.; Wang, W.; Yang, H. Rapid mutation breeding Schizochytrium strains producing high-yield docosahexenoic acid by atmospheric and room temperature plasmas (ARTP). Biotechnol. Bull. 2015, 31, 199–204. [Google Scholar]
- Gong, D.; Xiong, Y.; Sun, J.; Shi, Z. Mutation breeding Schizochytrium strains producing high-yield DHA via atmospheric and room temperature plasmas. China Oils Fats 2019, 44, 124–128. [Google Scholar]
- Lian, M.; Huang, H.; Ren, L.; Ji, X.; Zhu, J.; Jin, L. Increase of docosahexaenoic acid production by Schizochytrium sp through mutagenesis and enzyme assay. Appl. Biochem. Biotechnol. 2010, 162, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.R.; Sun, Z.J.; Cui, G.Z.; Song, X.; Cui, Q. A new strategy for strain improvement of Aurantiochytrium sp. based on heavy-ions mutagenesis and synergistic effects of cold stress and inhibitors of enoyl-ACP reductase. Enzym. Microb. Technol. 2016, 93–94, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Harwood, H.J. Acetyl-coenzyme A carboxylases: Versatile targets for drug discovery. J. Cell. Biochem. 2010, 99, 1476–1488. [Google Scholar]
- James, E.S.; Cronan, J.E. Expression of two Escherichia coli acetyl-CoA carboxylase subunits is autoregulated. J. Biol. Chem. 2004, 279, 2520–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Zeng, R.; Zhang, S.-X.; Yang, Z.-H.; Huang, H. Production of microalgal lipids as biodiesel feedstock with fixation of CO2 by Chlorella vulgaris. Food Technol. Biotech. 2014, 52, 285–291. [Google Scholar]
- Wan, M.; Liu, P.; Xia, J.; Rosenberg, J.N.; Oyler, G.A.; Betenbaugh, M.J.; Nie, Z.; Qiu, G. The effect of mixotrophy on microalgal growth, lipid content, and expression levels of three pathway genes in Chlorella sorokiniana. Appl. Microbiol. Biotechnol. 2011, 91, 835–844. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, D.; Zhang, J.; Chen, Y.; Liu, X.; Fan, C.; Wang, R.R.C.; Hou, Y.; Hu, Z. Overexpression of the transcription factor AtLEC1 significantly improved the lipid content of Chlorella ellipsoidea. Front. Bioeng. Biotechnol. 2021, 9, 626162. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.R.; Entcheva, P. Biotin in microbes, the genes involved in its biosynthesis, its biochemical role and perspectives for biotechnological production. Appl. Microbiol. Biotechnol. 2003, 61, 21–31. [Google Scholar] [CrossRef]
- Quilodran, B.; Hinzpeter, I.; Hormazabal, E.; Quiroz, A.; Shene, C. Docosahexaenoic acid (C22:6n-3, DHA) and astaxanthin production by Thraustochytriidae sp AS4-A1 a native strain with high similitude to Ulkenia sp.: Evaluation of liquid residues from food industry as nutrient sources. Enzym. Microb. Technol. 2010, 47, 24–30. [Google Scholar] [CrossRef]
- Ugalde, V.; Armenta, R.E.; Kermanshahi-pour, A.; Sun, Z.; Berryman, K.T.; Brooks, M.S. Improvement of culture conditions for cell biomass and fatty acid production by marine thraustochytrid F24-2. J. Appl. Phycol. 2018, 30, 329–339. [Google Scholar] [CrossRef]
- Lee Chang, K.J.; Dumsday, G.; Nichols, P.D.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A. High cell density cultivation of a novel Aurantiochytrium sp. strain TC 20 in a fed-batch system using glycerol to produce feedstock for biodiesel and omega-3 oils. Appl. Microbiol. Biotechnol. 2013, 97, 6907–6918. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-T.; Yu, S.-T.; Lin, L.-P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. S31. Process Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable Diversity and Biochemical Features of Thraustochytrids from Coastal Waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Wang, Q.; Sen, B.; Liu, X.; He, Y.; Xie, Y.; Wang, G. Enhanced saturated fatty acids accumulation in cultures of newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. for large-scale biodiesel production. Sci. Total Environ. 2018, 631–632, 994–1004. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Mutant | Biomass (g/L) | TFA (g/L) | DHA (g/L) | DHA (g/g) | DHA/TFA (%) |
|---|---|---|---|---|---|
| WT | 6.13 | 2.36 | 0.61 | 0.1 | 25.8 |
| A10 | 6.42 (4.7%) | 2.47 (4.7%) | 0.87 ** (42.6%) | 0.14 (40%) | 35.2 |
| A11 | 6.31 (2.9%) | 2.40 (1.7%) | 0.85 ** (39.3%) | 0.13 (30%) | 35.4 |
| A17 | 6.47 (5.5%) | 2.44 (3.4%) | 0.87 ** (42.6%) | 0.13 (30%) | 35.7 |
| A22 | 6.46 * (5.4%) | 2.66 (12.7%) | 0.92 ** (50.8%) | 0.14 (40%) | 34.6 |
| A23 | 6.07 (−1.0%) | 2.55 (8.1%) | 0.87 ** (42.6%) | 0.14 (40%) | 34.1 |
| A25 | 6.33 (3.3%) | 2.60 (10.2%) | 0.86 ** (41.0%) | 0.14 (40%) | 33.1 |
| A29 | 6.71 * (9.5%) | 2.66 (12.7%) | 0.95 ** (55.7%) | 0.14 (40%) | 35.7 |
| A36 | 6.49 * (5.9%) | 2.51 (6.4%) | 0.86 ** (41.0%) | 0.13 (30%) | 34.3 |
| A49 | 6.59 * (7.5%) | 2.40 (1.7%) | 0.91 ** (49.2%) | 0.14 (40%) | 37.9 |
| A54 | 6.28 * (2.4%) | 2.51 (6.4%) | 0.85 ** (39.3%) | 0.14 (40%) | 33.9 |
| A74 | 6.75 ** (10.1%) | 2.49 (5.5%) | 0.95 ** (55.7%) | 0.14 (40%) | 38.2 |
| A75 | 6.51 * (6.2%) | 2.74 (16.1%) | 0.87 ** (42.6%) | 0.13 (30%) | 31.8 |
| A78 | 6.35 (3.6%) | 2.81 * (19.1%) | 0.94 ** (54.1%) | 0.15 (50%) | 33.5 |
| A81 | 6.85 ** (11.7%) | 2.48 (5.1%) | 0.83 ** (36.1%) | 0.12 (20%) | 33.5 |
| A89 | 6.28 (2.4%) | 2.81 (19.1%) | 0.95 ** (55.7%) | 0.15 (50%) | 33.8 |
| A92 | 6.27 (2.3%) | 2.72 (15.3%) | 0.87 ** (42.6%) | 0.14 (40%) | 32.0 |
| A99 | 6.70 ** (9.3%) | 2.92 (23.7%) | 0.98 ** (60.7%) | 0.15 (50%) | 33.6 |
| Experiment | A Glucose (g/L) | B Yeast Extract (g/L) | C Sea Salt (%) * |
|---|---|---|---|
| 1 | 30 | 6.5 | 50 |
| 2 | 40 | 7.5 | 60 |
| 3 | 50 | 8.5 | 70 |
| Experiment | A Biotin (g/L) | B Citric Acid (g/L) | C Sodium Citrate (g/L) |
|---|---|---|---|
| 1 | 0.3 | 0.5 | 0.5 |
| 2 | 0.4 | 0.6 | 0.6 |
| 3 | 0.5 | 0.7 | 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Bai, M.; Zhang, S.; Li, J.; Liu, X.; Sen, B.; Wang, G. ARTP Mutagenesis of Schizochytrium sp. PKU#Mn4 and Clethodim-Based Mutant Screening for Enhanced Docosahexaenoic Acid Accumulation. Mar. Drugs 2021, 19, 564. https://doi.org/10.3390/md19100564
Liu L, Bai M, Zhang S, Li J, Liu X, Sen B, Wang G. ARTP Mutagenesis of Schizochytrium sp. PKU#Mn4 and Clethodim-Based Mutant Screening for Enhanced Docosahexaenoic Acid Accumulation. Marine Drugs. 2021; 19(10):564. https://doi.org/10.3390/md19100564
Chicago/Turabian StyleLiu, Lu, Mohan Bai, Sai Zhang, Jiantao Li, Xianhua Liu, Biswarup Sen, and Guangyi Wang. 2021. "ARTP Mutagenesis of Schizochytrium sp. PKU#Mn4 and Clethodim-Based Mutant Screening for Enhanced Docosahexaenoic Acid Accumulation" Marine Drugs 19, no. 10: 564. https://doi.org/10.3390/md19100564
APA StyleLiu, L., Bai, M., Zhang, S., Li, J., Liu, X., Sen, B., & Wang, G. (2021). ARTP Mutagenesis of Schizochytrium sp. PKU#Mn4 and Clethodim-Based Mutant Screening for Enhanced Docosahexaenoic Acid Accumulation. Marine Drugs, 19(10), 564. https://doi.org/10.3390/md19100564