
Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies



Abstract
:
1. Introduction
2. Health-Promoting Carotenoids from Microalgae and Their Biofunctions
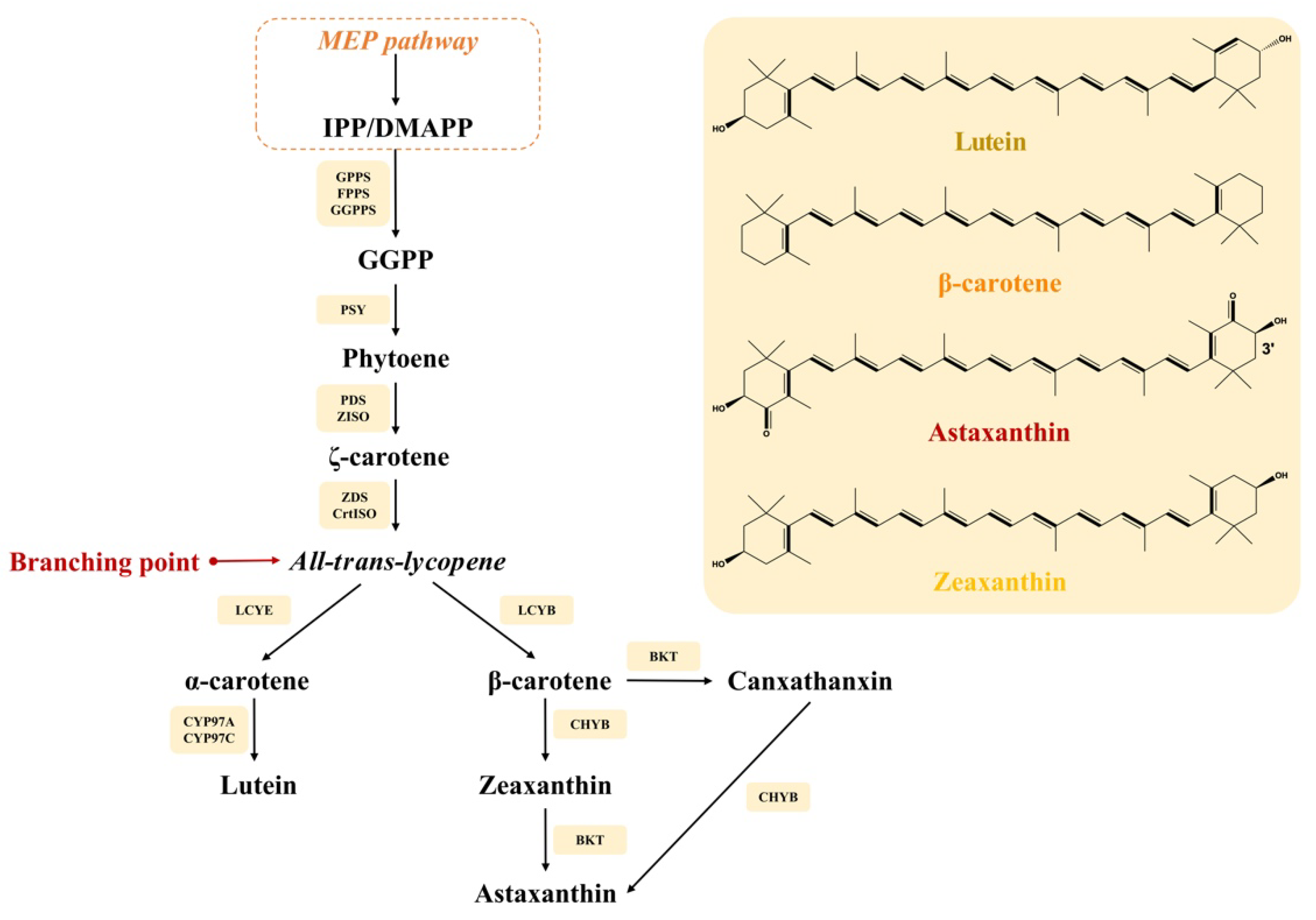
2.1. Carotenoid Biosynthesis in Microalgae
2.2. Health-Promoting Carotenoids and Their Production from Microalgae
2.2.1. Lutein
2.2.2. Astaxanthin
2.2.3. β-Carotene
2.2.4. Zeaxanthin
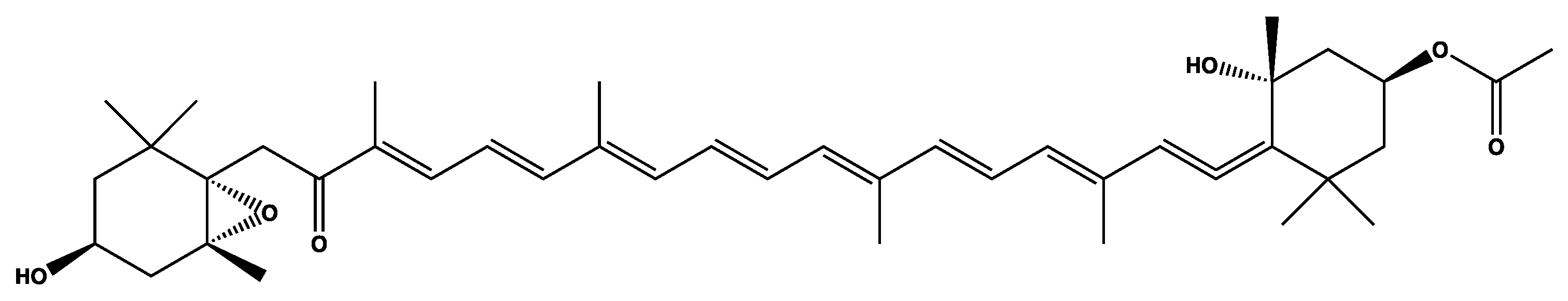
2.2.5. Fucoxanthin
3. Salt-Stress Treatment for Carotenoid Production from Microalgae
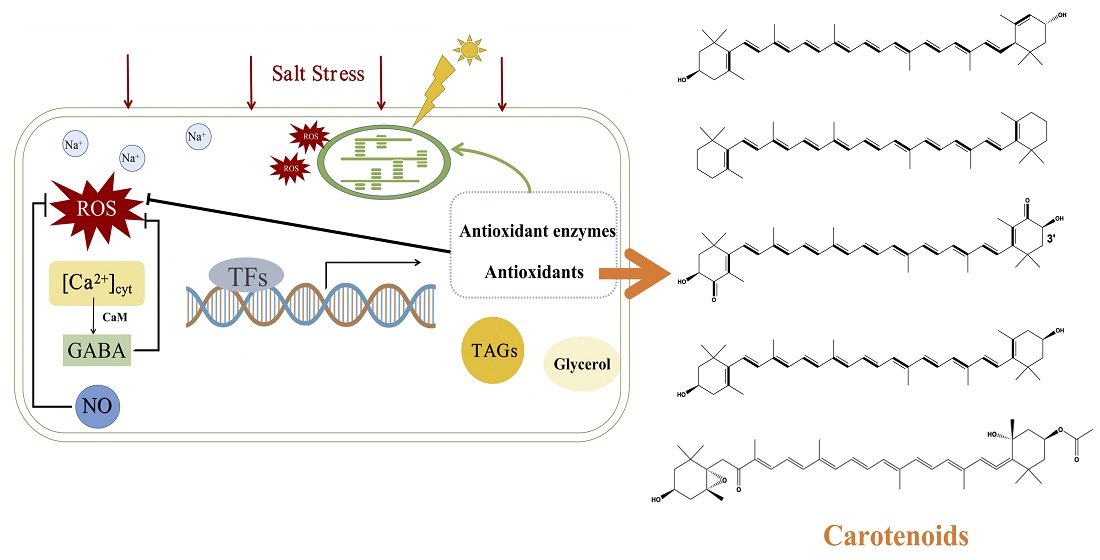
3.1. Microalgal Responses to Salt Stress
3.1.1. Early Signaling
3.1.2. Downstream Signaling
3.2. Salt Stress Strategies for Carotenoid Accumulation
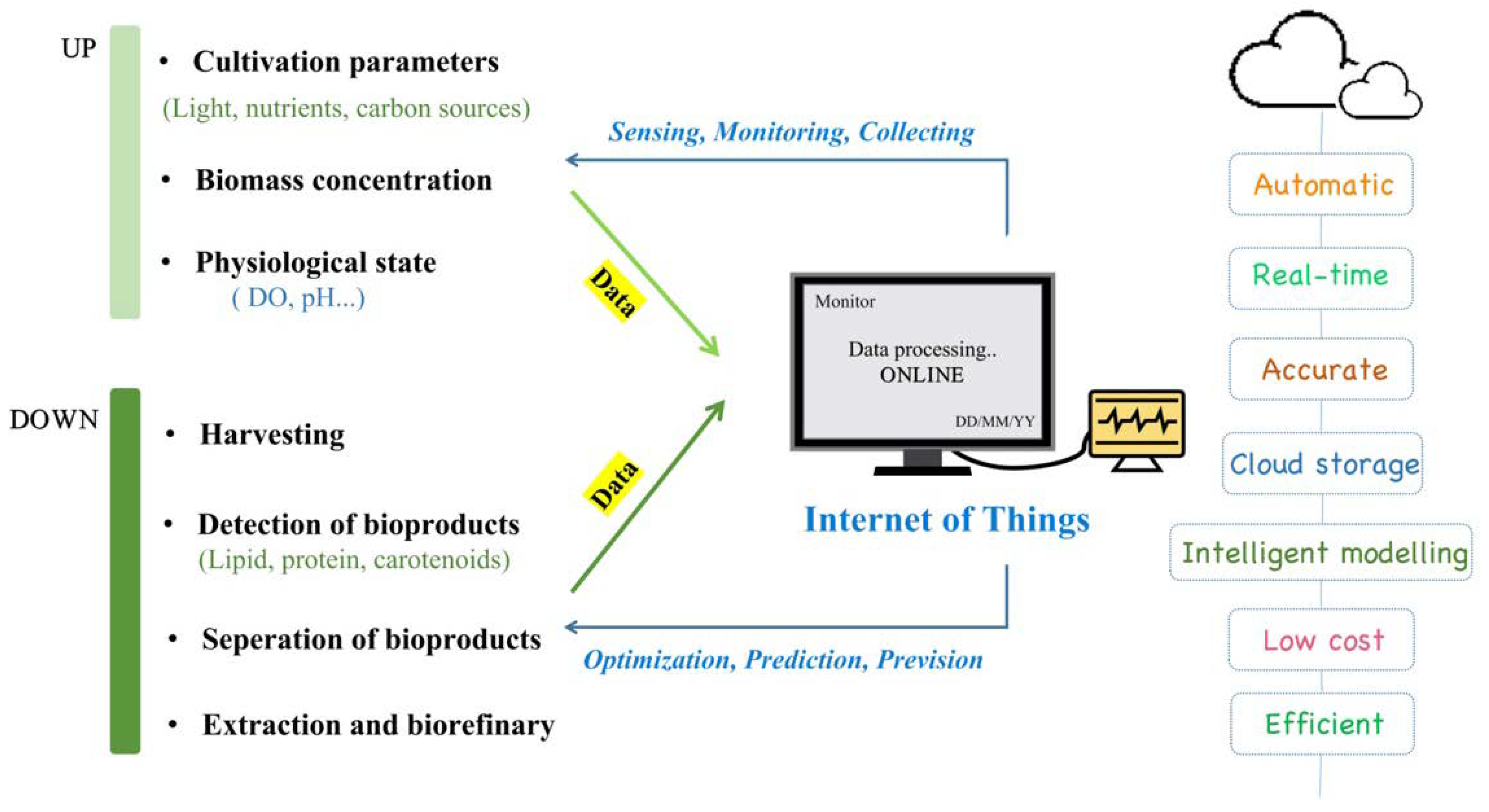
4. Potential Applications of Internet of Things (IoT) in Carotenoids Production
5. Future Prospects
5.1. Genetic Modifications of Microalgae for Salt Tolerance and Carotenoid Accmulation
5.2. Co-Production of Carotenoids with Value-Added Products from Microalgae
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.; Hu, B.; Cheng, Y.; Guo, Y.; Yao, W.; Qian, H. Carotenoids from fungi and microalgae: A review on their recent production, extraction, and developments. Bioresour. Technol. 2021, 337, 125398. [Google Scholar] [CrossRef]
- Qiang, S.; Su, A.P.; Li, Y.; Chen, Z.; Hu, C.Y.; Meng, Y.H. Elevated β-Carotene Synthesis by the Engineered Rhodobacter sphaeroides with Enhanced CrtY Expression. J. Agric. Food Chem. 2019, 67, 9560–9568. [Google Scholar] [CrossRef]
- Roberts, R.L.; Green, J.; Lewis, B. Lutein and zeaxanthin in eye and skin health. Clin. Dermatol. 2009, 27, 195–201. [Google Scholar] [CrossRef]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Kumar, L.; Bharadvaja, N. A review on microalgae biofuel and biorefinery: Challenges and way forward. Energy Sources Part A Recover. Util. Environ. Eff. 2020, 42, 1–24. [Google Scholar] [CrossRef]
- Liu, Z.; Berg, C.V.D.; Weusthuis, R.A.; Dragone, G.; Mussatto, S.I. Strategies for an improved extraction and separation of lipids and carotenoids from oleaginous yeast. Sep. Purif. Technol. 2020, 257, 117946. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef] [PubMed]
- Sinetova, M.A.; Sidorov, R.A.; Medvedeva, A.A.; Starikov, A.Y.; Markelova, A.G.; Allakhverdiev, S.I.; Los, D.A. Effect of salt stress on physiological parameters of microalgae Vischeria punctata strain IPPAS H-242, a superproducer of eicosapentaenoic acid. J. Biotechnol. 2021, 331, 63–73. [Google Scholar] [CrossRef]
- Ren, Y.; Deng, J.; Huang, J.; Wu, Z.; Yi, L.; Bi, Y.; Chen, F. Using green alga Haematococcus pluvialis for astaxanthin and lipid co-production: Advances and outlook. Bioresour. Technol. 2021, 340, 125736. [Google Scholar] [CrossRef] [PubMed]
- Liyanaarachchi, V.C.; Premaratne, M.; Ariyadasa, T.U.; Nimarshana, P.; Malik, A. Two-stage cultivation of microalgae for production of high-value compounds and biofuels: A review. Algal Res. 2021, 57, 102353. [Google Scholar] [CrossRef]
- Martin, L.J. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [Green Version]
- Bourdon, L.; Jensen, A.A.; Kavanagh, J.M.; McClure, D.D. Microalgal production of zeaxanthin. Algal Res. 2021, 55, 102266. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Zhang, Y.; Wang, X.; Liu, J. Novel insights into salinity-induced lipogenesis and carotenogenesis in the oleaginous astaxanthin-producing alga Chromochloris zofingiensis: A multi-omics study. Biotechnol. Biofuels 2020, 13, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Li, L. Toward the ‘golden’ era: The status in uncovering the regulatory control of carotenoid accumulation in plants. Plant Sci. 2019, 290, 110331. [Google Scholar] [CrossRef]
- Bhat, I.; Mamatha, B.S. Genetic factors involved in modulating lutein bioavailability. Nutr. Res. 2021, 91, 36–43. [Google Scholar] [CrossRef]
- Fang, N.; Wang, C.; Liu, X.; Zhao, X.; Liu, Y.; Liu, X.; Du, Y.; Zhang, Z.; Zhang, H. De novo synthesis of astaxanthin: From organisms to genes. Trends Food Sci. Technol. 2019, 92, 162–171. [Google Scholar] [CrossRef]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the Provitamin A (β-Carotene) Biosynthetic Pathway into (Carotenoid-Free) Rice Endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [Green Version]
- Bouyahya, A.; El Omari, N.; Hakkur, M.; El Hachlafi, N.; Charfi, S.; Balahbib, A.; Guaouguaou, F.-E.; Rebezov, M.; Maksimiuk, N.; Shariati, M.A.; et al. Sources, health benefits, and biological properties of zeaxanthin. Trends Food Sci. Technol. 2021, 118, 519–538. [Google Scholar] [CrossRef]
- Muradian, K.; Vaiserman, A.; Min, K.-J.; Fraifeld, V. Fucoxanthin and lipid metabolism: A minireview. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Butnariu, M.; Rodino, S.; Petrache, P.; Butu, M. Determination and quantification of maize zeaxanthin stability. Dig. J. Nanomater. Bios. 2014, 9, 745–755. [Google Scholar]
- Lin, J.-H.; Lee, D.-J.; Chang, J.-S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2014, 184, 421–428. [Google Scholar] [CrossRef]
- Fernández-Sevilla, J.M.; Fernandez, F.G.A.; Grima, E.M. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, Y.; Li, S.; Nagarajan, D.; Varjani, S.; Lee, D.-J.; Chang, J.-S. Recent advances in lutein production from microalgae. Renew. Sustain. Energy Rev. 2021, 153, 111795. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Subramanian, G.; Dash, S.K.; Sen, R. Development of an optimal light-feeding strategy coupled with semi-continuous reactor operation for simultaneous improvement of microalgal photosynthetic efficiency, lutein production and CO2 sequestration. Biochem. Eng. J. 2016, 113, 47–56. [Google Scholar] [CrossRef]
- Chen, J.-H.; Chen, C.-Y.; Hasunuma, T.; Kondo, A.; Chang, C.-H.; Ng, I.-S.; Chang, J.-S. Enhancing lutein production with mixotrophic cultivation of Chlorella sorokiniana MB-1-M12 using different bioprocess operation strategies. Bioresour. Technol. 2019, 278, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, M.-M.; Sun, Z.; Liu, S.-F.; Qin, Z.-H.; Mou, J.; Zhou, Z.-G.; Lin, C.S.K. Sustainable lipid and lutein production from Chlorella mixotrophic fermentation by food waste hydrolysate. J. Hazard. Mater. 2020, 400, 123258. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhang, Z.; Ho, S.-H.; Ruan, C.; Li, J.; Xie, Y.; Shi, X.; Liu, L.; Chen, J. Two-stage bioprocess for hyper-production of lutein from microalga Chlorella sorokiniana FZU60: Effects of temperature, light intensity, and operation strategies. Algal Res. 2020, 52, 102119. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Sen, R.; Sarmah, A.K. Consolidated bioprocessing of wastewater cocktail in an algal biorefinery for enhanced biomass, lipid and lutein production coupled with efficient CO2 capture: An advanced optimization approach. J. Environ. Manag. 2019, 252, 109696. [Google Scholar] [CrossRef]
- Chen, J.-H.; Kato, Y.; Matsuda, M.; Chen, C.-Y.; Nagarajan, D.; Hasunuma, T.; Kondo, A.; Dong, C.-D.; Lee, D.-J.; Chang, J.-S. A novel process for the mixotrophic production of lutein with Chlorella sorokiniana MB-1-M12 using aquaculture wastewater. Bioresour. Technol. 2019, 290, 121786. [Google Scholar] [CrossRef]
- Heo, J.; Shin, D.-S.; Cho, K.; Cho, D.-H.; Lee, Y.J.; Kim, H.-S. Indigenous microalga Parachlorella sp. JD-076 as a potential source for lutein production: Optimization of lutein productivity via regulation of light intensity and carbon source. Algal Res. 2018, 33, 1–7. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced High-Yield Production of Zeaxanthin, Lutein, and β-Carotene by a Mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Bassi, A. Investigation of Chlorella vulgaris UTEX 265 Cultivation under Light and Low Temperature Stressed Conditions for Lutein Production in Flasks and the Coiled Tree Photo-Bioreactor (CTPBR). Appl. Biochem. Biotechnol. 2017, 183, 652–671. [Google Scholar] [CrossRef]
- McClure, D.D.; Nightingale, J.K.; Luiz, A.; Black, S.; Zhu, J.; Kavanagh, J.M. Pilot-scale production of lutein using Chlorella vulgaris. Algal Res. 2019, 44, 101707. [Google Scholar] [CrossRef]
- Přibyl, P.; Pilný, J.; Cepák, V.; Kaštánek, P. The role of light and nitrogen in growth and carotenoid accumulation in Scenedesmus sp. Algal Res. 2016, 16, 69–75. [Google Scholar] [CrossRef]
- Wan, M.; Hou, D.; Li, Y.; Fan, J.; Huang, J.; Liang, S.; Wang, W.; Pan, R.; Wang, J.; Li, S. The effective photoinduction of Haematococcus pluvialis for accumulating astaxanthin with attached cultivation. Bioresour. Technol. 2014, 163, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Z.; Gerken, H.; Liu, Z.; Jiang, Y.; Chen, F. Chlorella zofingiensis as an Alternative Microalgal Producer of Astaxanthin: Biology and Industrial Potential. Mar. Drugs 2014, 12, 3487–3515. [Google Scholar] [CrossRef] [Green Version]
- Tharek, A.; Yahya, A.; Salleh, M.M.; Jamaluddin, H.; Yoshizaki, S.; Hara, H.; Iwamoto, K.; Suzuki, I.; Mohamad, S.E. Improvement and screening of astaxanthin producing mutants of newly isolated Coelastrum sp. using ethyl methane sulfonate induced mutagenesis technique. Biotechnol. Rep. 2021, 32, e00673. [Google Scholar] [CrossRef]
- Rammuni, M.; Ariyadasa, T.U.; Nimarshana, P.; Attalage, R. Comparative assessment on the extraction of carotenoids from microalgal sources: Astaxanthin from H. pluvialis and β-carotene from D. salina. Food Chem. 2018, 277, 128–134. [Google Scholar] [CrossRef]
- Mitra, M.; Mishra, S. A comparative analysis of different extraction solvent systems on the extractability of eicosapentaenoic acid from the marine eustigmatophyte Nannochloropsis oceanica. Algal Res. 2019, 38, 101387. [Google Scholar] [CrossRef]
- Pushpalatha, S.; Sangeetha, R.; Ariraman, S.; Ashokkumar, B.; Varalakshmi, P. Photocatalyst (TiO2) as an enhancer: An at-tempt to enhance the production of carotenoids and lipids with the combined oxidative stresses in Coelastrella sp. MClean Technol. Environ. Policy 2020, 23, 41–53. [Google Scholar] [CrossRef]
- Jin, E.; Feth, B.; Melis, A. A mutant of the green alga Dunaliella salina constitutively accumulates zeaxanthin under all growth conditions. Biotechnol. Bioeng. 2002, 81, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.Y.; Cha, K.H.; Song, D.-G.; Chung, D.; Pan, C.-H. Optimization of pressurized liquid extraction of zeaxanthin from Chlorella ellipsoidea. Environ. Boil. Fishes 2011, 24, 725–730. [Google Scholar] [CrossRef]
- Petrushkina, M.; Gusev, E.; Sorokin, B.; Zotko, N.; Mamaeva, A.; Filimonova, A.; Kulikovskiy, M.; Maltsev, Y.; Yampolsky, I.; Guglya, E.; et al. Fucoxanthin production by heterokont microalgae. Algal Res. 2017, 24, 387–393. [Google Scholar] [CrossRef]
- Li, Y.; Sun, H.; Wu, T.; Fu, Y.; He, Y.; Mao, X.; Chen, F. Storage carbon metabolism of Isochrysis zhangjiangensis under different light intensities and its application for co-production of fucoxanthin and stearidonic acid. Bioresour. Technol. 2019, 282, 94–102. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.-W.; Kwon, O.N.; Chung, D.; Pan, C.-H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Gao, F.; Cabanelas, I.I.T.; Wijffels, R.H.; Barbosa, M.J. Process optimization of fucoxanthin production with Tisochrysis lutea. Bioresour. Technol. 2020, 315, 123894. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.-J.; Kwon, O.-N.; Cha, K.H.; Um, B.-H.; Chung, D.; Pan, C.-H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Zhao, T.; Yan, X.; Sun, L.; Yang, T.; Hu, X.; He, Z.; Liu, F.; Liu, X. Research progress on extraction, biological activities and delivery systems of natural astaxanthin. Trends Food Sci. Technol. 2019, 91, 354–361. [Google Scholar] [CrossRef]
- Shah, M.; Mahfuzur, R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [Green Version]
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel Insights into the Biotechnological Production of Haematococcus pluvialis-Derived Astaxanthin: Advances and Key Challenges to Allow Its Industrial Use as Novel Food Ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ren, Y.; Fan, Y.; Lu, X.; Zhao, W.; Chen, F. Systematic metabolic tools reveal underlying mechanism of product biosynthesis in Chromochloris zofingiensis. Bioresour. Technol. 2021, 337, 125406. [Google Scholar] [CrossRef]
- Gorgich, M.; Martins, A.A.; Mata, T.M.; Caetano, N.S. Composition, cultivation and potential applications of Chlorella zofingiensis—A comprehensive review. Algal Res. 2021, 60, 102508. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Jiang, H.; Mao, X. Biotechnology advances in β-carotene production by microorganisms. Trends Food Sci. Technol. 2021, 111, 322–332. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; De Morais, A.M.M.B.; Morais, R. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.; Bino, R.J.; Wijffels, R.H. Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol. 2008, 26, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Roso, G.R. The Bioeconomy of Microalgal Carotenoid-Rich Oleoresins Produced in Agroindustrial Biorefineries. J. Biosens. Bioelectron. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, D.B.; Mercadante, A.; Mariutti, L.R.B. Marigold carotenoids: Much more than lutein esters. Food Res. Int. 2018, 119, 653–664. [Google Scholar] [CrossRef]
- Matsuno, T. Aquatic animal carotenoids. Fish. Sci. 2001, 67, 771–783. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chen, C.-Y.; Varjani, S.; Chang, J.-S. Producing fucoxanthin from algae—Recent advances in cultivation strategies and downstream processing. Bioresour. Technol. 2021, 344, 126170. [Google Scholar] [CrossRef] [PubMed]
- Mohamadnia, S.; Tavakoli, O.; Faramarzi, M.A. Enhancing production of fucoxanthin by the optimization of culture media of the microalga Tisochrysis lutea. Aquaculture 2020, 533, 736074. [Google Scholar] [CrossRef]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.-H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Ozaki, Y.; Hashimoto, T.; Das, S.K.; Matsushita, S.; Hirano, M.; Okada, T.; Komoto, A.; Mori, N.; Nakatsuka, M. Commercial-scale Preparation of Biofunctional Fucoxanthin from Waste Parts of Brown Sea Algae Laminalia japonica. Food Sci. Technol. Res. 2008, 14, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Kajikawa, T.; Okumura, S.; Iwashita, T.; Kosumi, D.; Hashimoto, H.; Katsumura, S. Stereocontrolled Total Synthesis of Fucoxanthin and Its Polyene Chain-Modified Derivative. Org. Lett. 2012, 14, 808–811. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Kou, Y.; Liu, M.; Sun, P.; Dong, Z.; Liu, J. High light boosts salinity stress-induced biosynthesis of astaxanthin and lipids in the green alga Chromochloris zofingiensis. Algal Res. 2020, 50, 101976. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-G.; Toyota, M.; Kim, S.-H.; Hilleary, R.; Gilroy, S. Salt stress-induced Ca2+ waves are associated with rapid, long-distance root-to-shoot signaling in plants. Proc. Natl. Acad. Sci. USA 2014, 111, 6497–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, G.L.; Brownlee, C. Ca2+ signalling in plants and green algae—Changing channels. Trends Plant Sci. 2008, 13, 506–514. [Google Scholar] [CrossRef]
- Khanna, R.R.; Jahan, B.; Iqbal, N.; Khan, N.A.; AlAjmi, M.F.; Rehman, T.; Khan, M.I.R. GABA reverses salt-inhibited photosynthetic and growth responses through its influence on NO-mediated nitrogen-sulfur assimilation and antioxidant system in wheat. J. Biotechnol. 2020, 325, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.M.; Singh, N.K.; Cherry, J.H.; Locy, R.D. Nitrate uptake and utilization is modulated by exogenous γ-aminobutyric acid in Arabidopsis thaliana seedlings. Plant Physiol. Biochem. 2010, 48, 443–450. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of nitric oxide in tolerance of plants to abiotic stress. Protoplasma 2010, 248, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Procházková, D.; Haisel, D.; Wilhelmová, N.; Pavlíková, D.; Száková, J. Effects of exogenous nitric oxide on photosynthesis. Photosynthetica 2013, 51, 483–489. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.-P.; Han, B.; Yu, X. Coupling of abiotic stresses and phytohormones for the production of lipids and high-value by-products by microalgae: A review. Bioresour. Technol. 2018, 274, 549–556. [Google Scholar] [CrossRef]
- Li, Q.; You, J.; Qiao, T.; Zhong, D.-B.; Yu, X. Sodium chloride stimulates the biomass and astaxanthin production by Haematococcus pluvialis via a two-stage cultivation strategy. Bioresour. Technol. 2021, 344, 126214. [Google Scholar] [CrossRef] [PubMed]
- Elloumi, W.; Jebali, A.; Maalej, A.; Chamkha, M.; Sayadi, S. Effect of Mild Salinity Stress on the Growth, Fatty Acid and Carotenoid Compositions, and Biological Activities of the Thermal Freshwater Microalgae Scenedesmus sp. Biomolecules 2020, 10, 1515. [Google Scholar] [CrossRef]
- Rao, A.R.; Dayananda, C.; Sarada, R.; Shamala, T.; Ravishankar, G. Effect of salinity on growth of green alga Botryococcus braunii and its constituents. Bioresour. Technol. 2007, 98, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Kim, M.; Park, S.; Baek, K.; Oh, H.; Polle, J.E.; Jin, E. Comparative transcriptome analysis of short-term responses to salt and glycerol hyperosmotic stress in the green alga Dunaliella salina. Algal Res. 2020, 53, 102147. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Kim, Y.; Terng, E.L.; Peterson, L.; Cerutti, H. A multidomain enzyme, with glycerol-3-phosphate dehydrogenase and phosphatase activities, is involved in a chloroplastic pathway for glycerol synthesis in Chlamydomonas reinhardtii. Plant J. 2017, 90, 1079–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Ho, S.-H.; Vavricka, C.J.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Evolutionary engineering of salt-resistant Chlamydomonas sp. strains reveals salinity stress-activated starch-to-lipid biosynthesis switching. Bioresour. Technol. 2017, 245, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yuan, Y.; Cheng, D.; Gao, J.; Kong, L.; Zhao, Q.; Wei, W.; Sun, Y. Exploring stress tolerance mechanism of evolved freshwater strain Chlorella sp. S30 under 30 g/L salt. Bioresour. Technol. 2018, 250, 495–504. [Google Scholar] [CrossRef]
- Cheng, R.; Feng, J.; Zhang, B.-X.; Huang, Y.; Cheng, J.; Zhang, C.-X. Transcriptome and Gene Expression Analysis of an Oleaginous Diatom Under Different Salinity Conditions. BioEnergy Res. 2013, 7, 192–205. [Google Scholar] [CrossRef]
- Gao, Z.; Meng, C.; Chen, Y.C.; Ahmed, F.; Mangott, A.; Schenk, P.M.; Li, Y. Comparison of astaxanthin accumulation and biosynthesis gene expression of three Haematococcus pluvialis strains upon salinity stress. Environ. Boil. Fishes 2014, 27, 1853–1860. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.Á.; Rivas, J.; Guerrero, M.G. Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef]
- Bermejo, E.; Ruiz-Domínguez, M.C.; Cuaresma, M.; Vaquero, I.; Ramos-Merchante, A.; Vega, J.M.; Vílchez, C.; Garbayo, I. Production of lutein, and polyunsaturated fatty acids by the acidophilic eukaryotic microalga Coccomyxa onubensis under abiotic stress by salt or ultraviolet light. J. Biosci. Bioeng. 2018, 125, 669–675. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Y.; Ding, W.; Han, B.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Gamma-aminobutyric acid facilitates the simultaneous production of biomass, astaxanthin and lipids in Haematococcus pluvialis under salinity and high-light stress conditions. Bioresour. Technol. 2020, 320, 124418. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Zhao, Y.; Li, T.; Han, B.; Zhao, P.; Yu, X. Enhancing astaxanthin and lipid coproduction in Haematococcus pluvialis by the combined induction of plant growth regulators and multiple stresses. Bioresour. Technol. 2021, 344, 126225. [Google Scholar] [CrossRef] [PubMed]
- Khalili, Z.; Jalili, H.; Noroozi, M.; Amrane, A. Effect of linoleic acid and methyl jasmonate on astaxanthin content of Scenedesmus acutus and Chlorella sorokiniana under heterotrophic cultivation and salt shock conditions. Environ. Boil. Fishes 2019, 31, 2811–2822. [Google Scholar] [CrossRef]
- Maroli, A.; Narwane, V.S.; Gardas, B.B. Applications of IoT for achieving sustainability in agricultural sector: A comprehensive review. J. Environ. Manag. 2021, 298, 113488. [Google Scholar] [CrossRef] [PubMed]
- Kashani, M.H.; Madanipour, M.; Nikravan, M.; Asghari, P.; Mahdipour, E. A systematic review of IoT in healthcare: Applications, techniques, and trends. J. Netw. Comput. Appl. 2021, 192, 103164. [Google Scholar] [CrossRef]
- Akhter, R.; Sofi, S.A. Precision agriculture using IoT data analytics and machine learning. J. King Saud Univ.—Comput. Inf. Sci. 2021. [Google Scholar] [CrossRef]
- Wang, K.; Khoo, K.S.; Leong, H.Y.; Nagarajan, D.; Chew, K.W.; Ting, H.Y.; Selvarajoo, A.; Chang, J.-S.; Show, P.L. How does the Internet of Things (IoT) help in microalgae biorefinery? Biotechnol. Adv. 2021, 107819. [Google Scholar] [CrossRef]
- Tan, X.B.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Wong, C.Y.; Lee, K.T. Cultivation of microalgae for biodiesel production: A review on upstream and downstream processing. Chin. J. Chem. Eng. 2018, 26, 17–30. [Google Scholar] [CrossRef]
- Krujatz, F.; Fehse, K.; Jahnel, M.; Gommel, C.; Schurig, C.; Lindner, F.; Bley, T.; Weber, J.; Steingroewer, J. MicrOLED-photobioreactor: Design and characterization of a milliliter-scale Flat-Panel-Airlift-photobioreactor with optical process monitoring. Algal Res. 2016, 18, 225–234. [Google Scholar] [CrossRef]
- Barbosa, R.C.; Soares, J.; Martins, M.A. Low-cost and versatile sensor based on multi-wavelengths for real-time estimation of microalgal biomass concentration in open and closed cultivation systems. Comput. Electron. Agric. 2020, 176, 105641. [Google Scholar] [CrossRef]
- Haigh-Flórez, D.; Cano-Raya, C.; Bedoya, M.; Orellana, G. Rugged fibre-optic luminescent sensor for CO2 determination in microalgae photoreactors for biofuel production. Sens. Actuators B Chem. 2015, 221, 978–984. [Google Scholar] [CrossRef]
- Martin, J.; Dannenberg, A.; Detrell, G.; Ewald, S.F. Noninvasive process control of a microalgae-based system for automated treatment of polluted agricultural ground water transferred from the development of a biological Life Support Systems. In Proceedings of the International Conference on Environmental Systems; 2020. Available online: https://www.ices.space/conference-proceedings-2/ (accessed on 10 December 2021).
- Giannino, F.; Esposito, S.; Diano, M.; Cuomo, S.; Toraldo, G. A predictive Decision Support System (DSS) for a microalgae production plant based on Internet of Things paradigm. Concurr. Comput. Pract. Exp. 2018, 30, e4476. [Google Scholar] [CrossRef]
- Havlik, I.; Scheper, T.; Reardon, K.F. Monitoring of Microalgal Processes. In Microalgae Biotechnology; Posten, C., Feng Chen, S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 89–142. [Google Scholar]
- Benson, B.C.; Gutierrez-Wing, M.T.; Rusch, K.A. Optimization of the lighting system for a Hydraulically Integrated Serial Turbidostat Algal Reactor (HISTAR): Economic implications. Aquac. Eng. 2009, 40, 45–53. [Google Scholar] [CrossRef]
- Dufková, K.; Barták, M.; Morkusová, J.; Elster, J.; Hájek, J. Screening of growth phases of Antarctic algae and cyanobacteria cultivated on agar plates by chlorophyll fluorescence imaging. Czech Polar Rep. 2019, 9, 170–181. [Google Scholar] [CrossRef]
- Takahashi, T. Routine Management of Microalgae Using Autofluorescence from Chlorophyll. Molecules 2019, 24, 4441. [Google Scholar] [CrossRef] [Green Version]
- Mairet, F.; Moisan, M.; Bernard, O. Estimation of neutral lipid and carbohydrate quotas in microalgae using adaptive interval observers. Bioprocess Biosyst. Eng. 2013, 37, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrineau, M.M.; Zelzion, E.; Gross, J.; Price, D.C.; Boyd, J.; Bhattacharya, D. Evolution of salt tolerance in a laboratory reared population of Chlamydomonas reinhardtii. Environ. Microbiol. 2014, 16, 1755–1766. [Google Scholar] [CrossRef]
- Wang, L.; Xue, C.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Strain improvement of Chlorella sp. for phenol biodegradation by adaptive laboratory evolution. Bioresour. Technol. 2016, 205, 264–268. [Google Scholar] [CrossRef]
- Kamath, B.S.; Vidhyavathi, R.; Sarada, R.; Ravishankar, G. Enhancement of carotenoids by mutation and stress induced carotenogenic genes in Haematococcus pluvialis mutants. Bioresour. Technol. 2008, 99, 8667–8673. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, Y.; Sommerfeld, M.; Chen, F.; Hu, Q. Enhanced protection against oxidative stress in an astaxanthin-overproduction Haematococcus mutant (Chlorophyceae). Eur. J. Phycol. 2008, 43, 365–376. [Google Scholar] [CrossRef]
- Cheng, J.; Lu, H.; Huang, Y.; Li, K.; Huang, R.; Zhou, J.; Cen, K. Enhancing growth rate and lipid yield of Chlorella with nuclear irradiation under high salt and CO2 stress. Bioresour. Technol. 2016, 203, 220–227. [Google Scholar] [CrossRef]
- Muñoz, C.F.; Südfeld, C.; Naduthodi, M.I.; Weusthuis, R.A.; Barbosa, M.J.; Wijffels, R.H.; D’Adamo, S. Genetic engineering of microalgae for enhanced lipid production. Biotechnol. Adv. 2021, 52, 107836. [Google Scholar] [CrossRef]
- Cui, J.; Sun, T.; Chen, L.; Zhang, W. Engineering salt tolerance of photosynthetic cyanobacteria for seawater utilization. Biotechnol. Adv. 2020, 43, 107578. [Google Scholar] [CrossRef] [PubMed]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2020, 53, 102158. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Gerken, H.; Huang, J.; Jiang, Y.; Chen, F. Genetic engineering of the green alga Chlorella zofingiensis: A modified norflurazon-resistant phytoene desaturase gene as a dominant selectable marker. Appl. Microbiol. Biotechnol. 2014, 98, 5069–5079. [Google Scholar] [CrossRef] [PubMed]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the biosynthesis of fucoxanthin as targets for genetic engineering in Phaeodactylum tricornutum. Environ. Boil. Fishes 2015, 28, 123–129. [Google Scholar] [CrossRef]
- Chen, Y.; Bi, C.; Zhang, J.; Hou, H.; Gong, Z. Astaxanthin biosynthesis in transgenic Dunaliella salina (Chlorophyceae) enhanced tolerance to high irradiation stress. S. Afr. J. Bot. 2020, 133, 132–138. [Google Scholar] [CrossRef]
- Brar, A.; Kumar, M.; Soni, T.; Vivekanand, V.; Pareek, N. Insights into the genetic and metabolic engineering approaches to enhance the competence of microalgae as biofuel resource: A review. Bioresour. Technol. 2021, 339, 125597. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.; Yu, J.; Jeong, J.; Sim, S.J.; Bae, S.; Jin, E. Photoautotrophic production of macular pigment in a Chlamydomonas reinhardtii strain generated by using DNA-free CRISPR-Cas9 RNP-mediated mutagenesis. Biotechnol. Bioeng. 2017, 115, 719–728. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Han, D.; Sommerfeld, M.; Lu, Y.; Chen, F.; Hu, Q. Molecular mechanisms of the coordination between astaxanthin and fatty acid biosynthesis in Haematococcus pluvialis (Chlorophyceae). Plant J. 2014, 81, 95–107. [Google Scholar] [CrossRef]
- Guo, H.; Li, T.; Zhao, Y.; Yu, X. Role of copper in the enhancement of astaxanthin and lipid coaccumulation in Haematococcus pluvialis exposed to abiotic stress conditions. Bioresour. Technol. 2021, 335, 125265. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B. Are we consuming enough long chain omega-3 polyunsaturated fatty acids for optimal health? Prostaglandins Leukot. Essent. Fat. Acids 2011, 85, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Wang, X.; Liu, J. Screening of Isochrysis strains for simultaneous production of docosahexaenoic acid and fucoxanthin. Algal Res. 2019, 41, 101545. [Google Scholar] [CrossRef]
- Ye, Z.-W.; Jiang, J.-G.; Wu, G.-H. Biosynthesis and regulation of carotenoids in Dunaliella: Progresses and prospects. Biotechnol. Adv. 2008, 26, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Doan, Q.C.; Moheimani, N.R.; Mastrangelo, A.J.; Lewis, D.M. Microalgal biomass for bioethanol fermentation: Implications for hypersaline systems with an industrial focus. Biomass Bioenergy 2012, 46, 79–88. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoids | Natural Sources | Biofunctions | Recommended Dose | Ref. |
|---|---|---|---|---|
| Lutein | Marigold flower *; Yolk; Broccoli; Microalgae Orange-yellow fruits; Leafy green vegetables; | Antioxidant; Filter blue light; Prevent AMD; Prevent AD | 6 mg day−1 | [3,17] |
| Astaxanthin | Shrimp; Salmon; Crabs; Microalgae (Haematococcus pluvialis *) Phaffia rhodozyma | Antioxidant; Anti-aging; Anti-inflammatory; Anti-hypertensive; Anti-cancer; | 4–12 mg day−1 | [10,18] |
| β-carotene | Pumpkin; Mango; Carrots; Microalgae (Dunaliella salina *) | Vitamin A precursor; Antioxidant; Anti-cancer; Anti-cardiovascular; Immune enhancement | 600 μg RE 1/day | [19] |
| Zeaxanthin | Marigold flower *; Maize; Orange peppers; Microalgae; Scallions | Filter blue light; Improve visual acuity; Anti-cancer; Anti-inflammatory; Anti-allergy Against UV, skin redness | 2 mg day−1 | [13,20] |
| Fucoxanthin | Macroalgae *; Microalgae | Anti-cancer; Anti-hypertensive; Anti-inflammatory; Anti-obesity | − | [12,21] |
| Carotenoid | Microalgae | Content | Productivity/Yield | Ref. |
|---|---|---|---|---|
| Lutein | Chromochloris zofingiensis bkt1 (mutant) | 13.81 mg/g DW | 33.97 mg/L | [33] |
| Parachlorellasp. JD-076 | 11.87 mg/g DW | 25.0 mg/L/day | [32] | |
| Chlorella sorokiniana FZU60 | 11.22 mg/g DW | 8.25 mg/L/day | [29] | |
| Chlorella vulgaris UTEX 265 | 9.82 mg/g DW | 11.98 mg/g/day | [34] | |
| Chlorella vulgaris CS-41 | 9.0 mg/g DW | 1.56 mg/L/day | [35] | |
| Scenedesmus sp. | 7.47 mg/g DW | 19.70 mg/L/day | [36] | |
| Chlorella sp. GY-H4 | 8.9 mg/g DW | 10.50 mg/L/day | [28] | |
| Chlorella sorokiniana MB-1-M12 | 7.39 mg/g DW | 3.43 mg/L/day | [27] | |
| Chlorella minutissima MCC-27 | 7.05 mg/g DW | 6.34 mg/L/day | [26] | |
| Astaxanthin | Haematococcus pluvialis | 5% DW | 65.8 mg/m2/day | [37] |
| Chromochloris zofingiensis | 6.5 mg/g DW | 0.8 mg/L/day | [38] | |
| Coelastrum sp. G1-C1 (mutant) | − | 28.32 mg/L | [39] | |
| β-carotene | Dunaliellasalina * | 13% DW | − | [40] |
| Chromochloris zofingiensis bkt1 (mutant) | 7.18 mg/g DW | 34.64 ± 1.39 mg/L | [33] | |
| Zeaxanthin | Nannochloropsis oceanica CCNM 1081 * | 30.2 mg/g DW | − | [41] |
| Coelastrella sp. M60 | 13.15 mg/g DW | 0.72 mg/L/day | [42] | |
| Chromochloris zofingiensis bkt1 (mutant) | 7.00 mg/g DW | 36.79 ± 2.23 mg/L | [33] | |
| Dunaliellasalina zea1 (mutant) * | 5.9 mg/g DW | − | [43] | |
| Chlorella ellipsoidea * | 4.26 mg/g DW | − | [44] | |
| Fucoxanthin | Mallomonas sp. | 26.6 mg/g DW | − | [45] |
| Isochrysis zhanjiangensis * | 23.29 mg/g DW | 2.94 mg/L/day | [46] | |
| Odontella aurita * | 18.47 mg/g DW | 7.96 mg/L/day | [47] | |
| Isochrysis aff. Galbana * | 18.23 mg/g DW | − | [48] | |
| Tisochrysis lutea * | 16.39 mg/g DW | 9.81 mg/L/day | [49] | |
| Phaeodactylum tricornutum * | 16.33 mg/g DW | − | [50] |
| Responses | Low-Dose NaCl | High-Dose NaCl |
|---|---|---|
| Physiology | Growth ↑; Photosynthesis ↑ | Growth ↓; Chlorophyll content↓; Photosynthesis ↓ |
| Morphology | No significant changes | Cell size ↑; Cell wall ↑ Color change |
| Main carbon sinks | Carbohydrate ↑ | Carbohydrate (providing building blocks) ↓ TAGs ↑ |
| Gene expression |
(rbcL, rbcS) ↑ |
|
| Metabolites | Lutein ↑ | Secondary carotenoids ↑ TFA ↑ |
| Stress Conditions | Microalgae | Carotenoids | Fold Change | Ref. |
|---|---|---|---|---|
| 100–500 mM NaCl (Two-stage) | Coccomyxa onubensis | Lutein | 0.47-fold | [89] |
| 200 mM NaCl | C. zofingiensis CCAP 211/14 | Astaxanthin | 1.23-fold | [88] |
| 36.27 g/L NaCl | Tisochrysis lutea | Fucoxanthin | − | [64] |
| 2% NaCl (w/v) | Chromochloris zofingiensis bkt1 | Zeaxanthin | 1.38-fold | [33] |
| Lutein β-carotene | 0.22-fold | |||
| 0.36-fold | ||||
| High light + NaCl | Chromochloris zofingiensis | Astaxanthin | 7.53-fold | [70] |
| LA + NaCl (20%) | Chlorella sorokiniana | Astaxanthin | 1.25-fold | [92] |
| GABA + high light + NaCl | Haematococcus pluvialis | Astaxanthin | 3.24-fold | [90] |
| MT+ N-starvation + NaCl | Haematococcus pluvialis | Astaxanthin | 1.20-fold | [91] |
| TiO2 + N-starvation + NaCl | Coelastrella sp. | Zeaxanthin | 0.51-fold | [42] |
| Astaxanthin | 1.16-fold |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Sun, H.; Deng, J.; Huang, J.; Chen, F. Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Mar. Drugs 2021, 19, 713. https://doi.org/10.3390/md19120713
Ren Y, Sun H, Deng J, Huang J, Chen F. Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Marine Drugs. 2021; 19(12):713. https://doi.org/10.3390/md19120713
Chicago/Turabian StyleRen, Yuanyuan, Han Sun, Jinquan Deng, Junchao Huang, and Feng Chen. 2021. "Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies" Marine Drugs 19, no. 12: 713. https://doi.org/10.3390/md19120713
APA StyleRen, Y., Sun, H., Deng, J., Huang, J., & Chen, F. (2021). Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Marine Drugs, 19(12), 713. https://doi.org/10.3390/md19120713