
Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression


 , , ,
, , , 
Abstract
:1. Introduction
2. Results
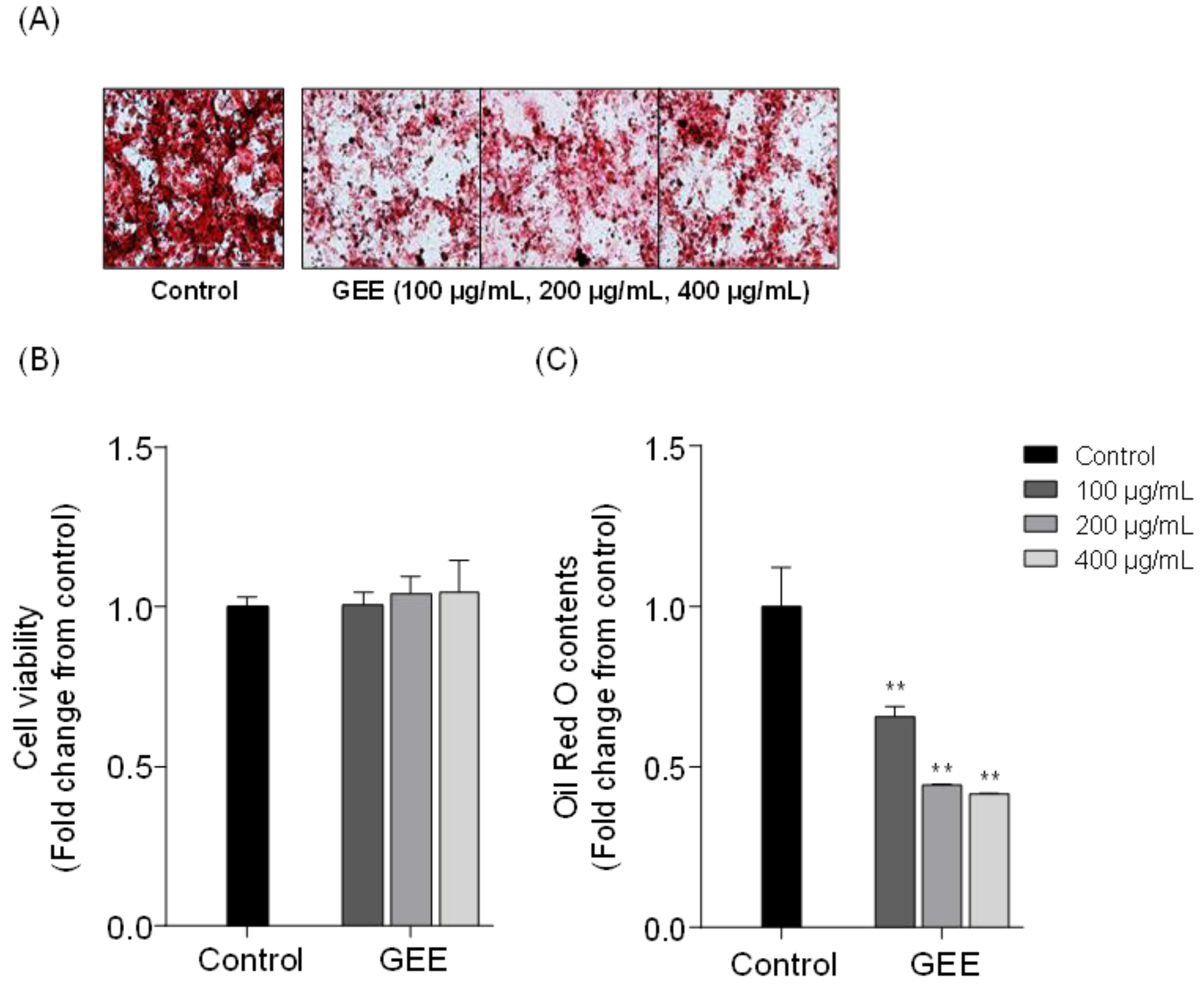
2.1. Effects of Red Seaweed Extract on Lipid Accumulation in 3T3-L1 Cells
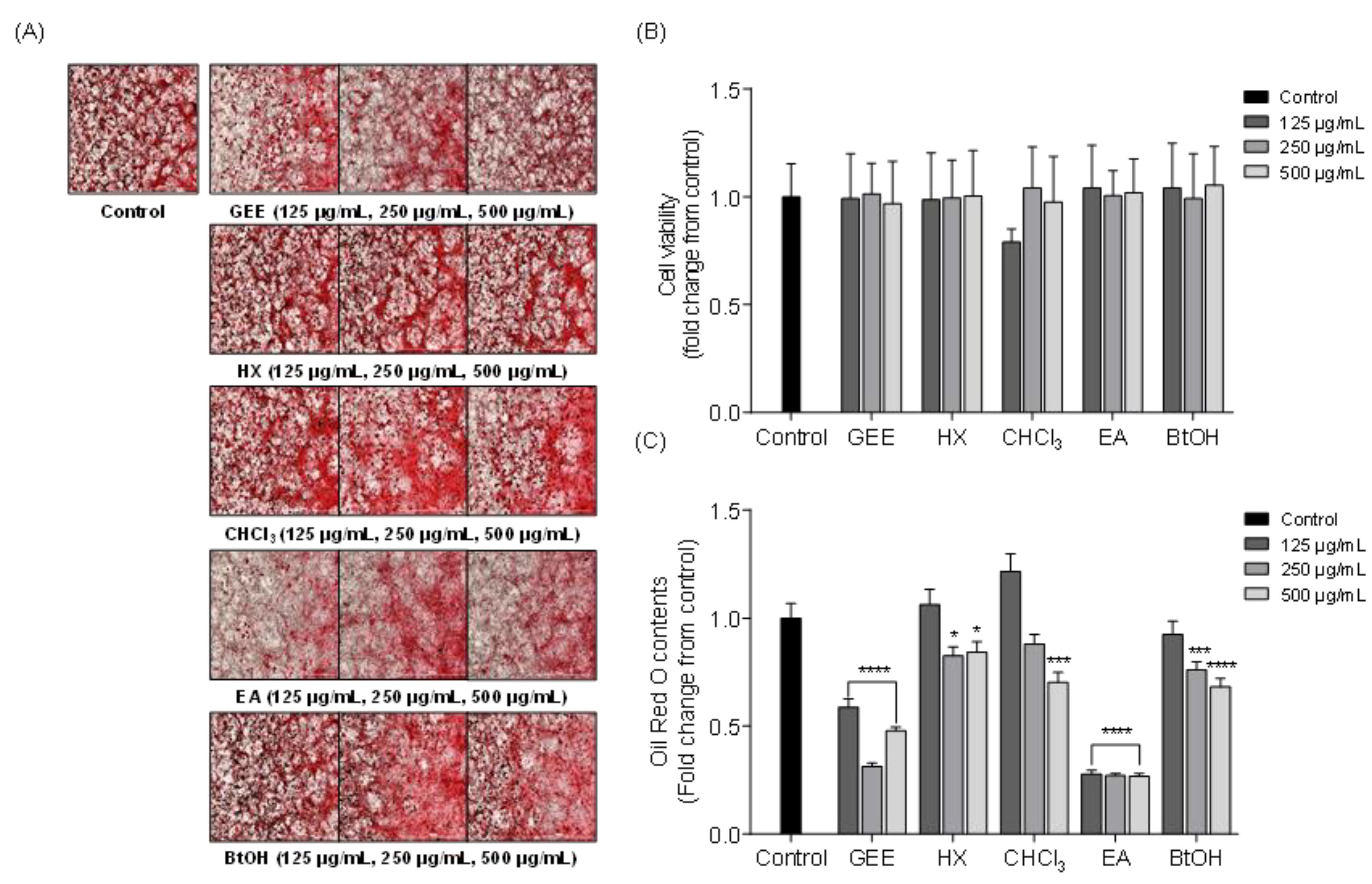
2.2. Effects of Solvent Fractions from GEE on Intracellular Lipid Accumulation in 3T3-L1 Cells
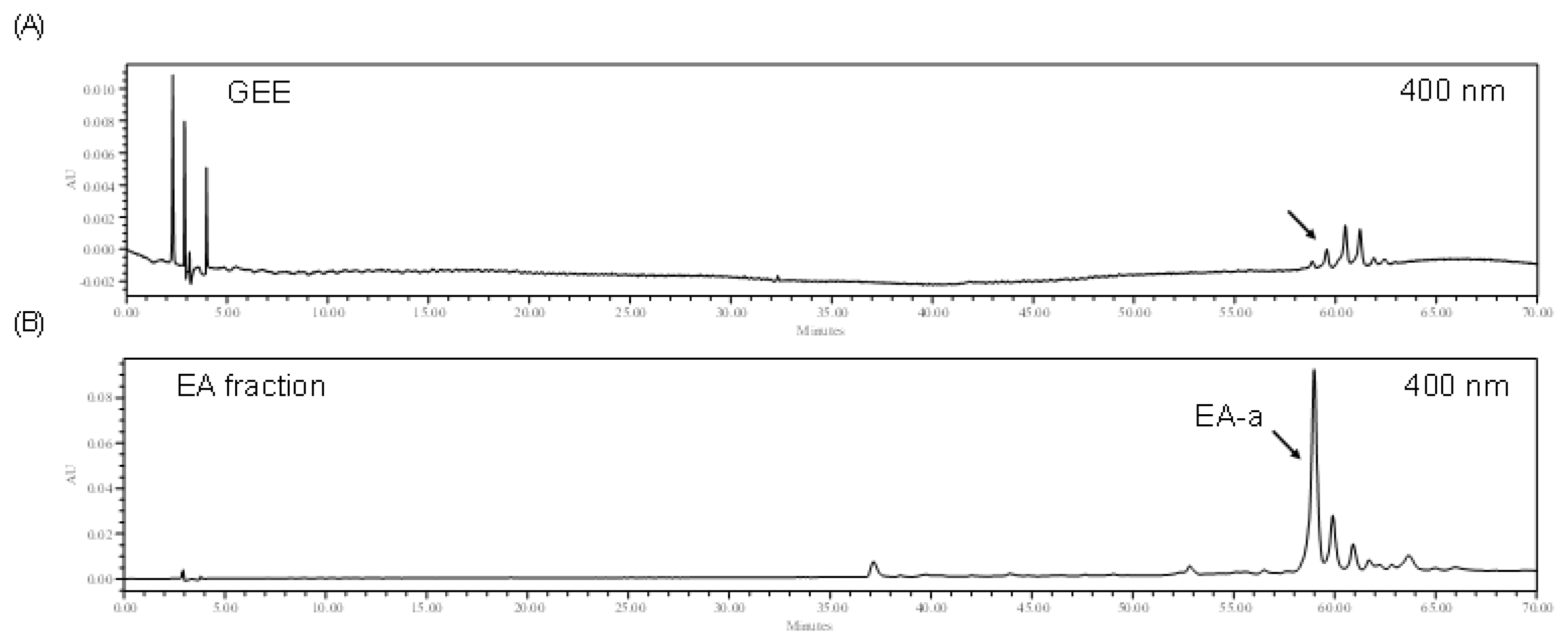
2.3. High Performance Liquid Chromatography Analysis of the EA Fraction and GEE
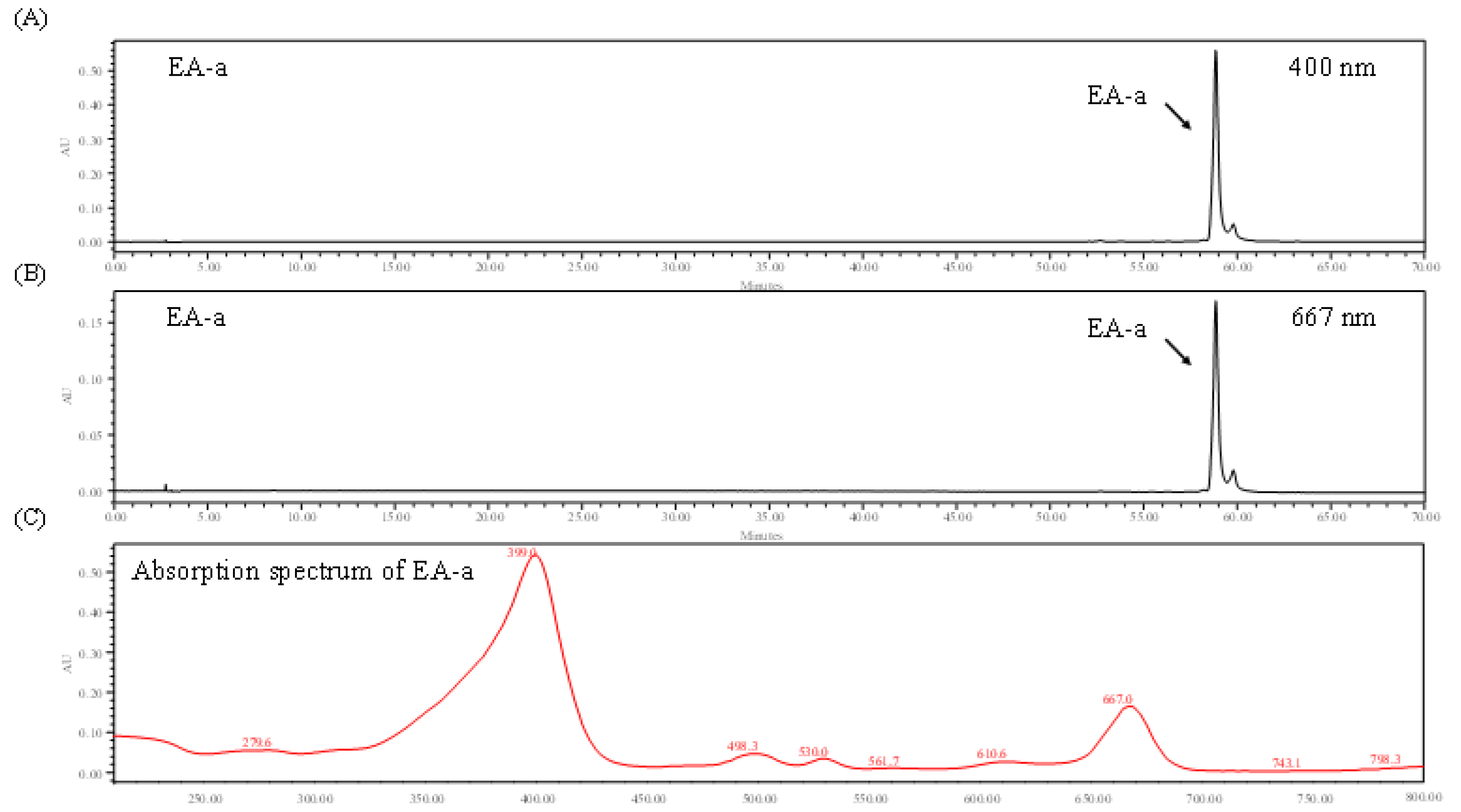
2.4. Identification of Chlorophyll Derivative via HPLC and Absorption Spectrum
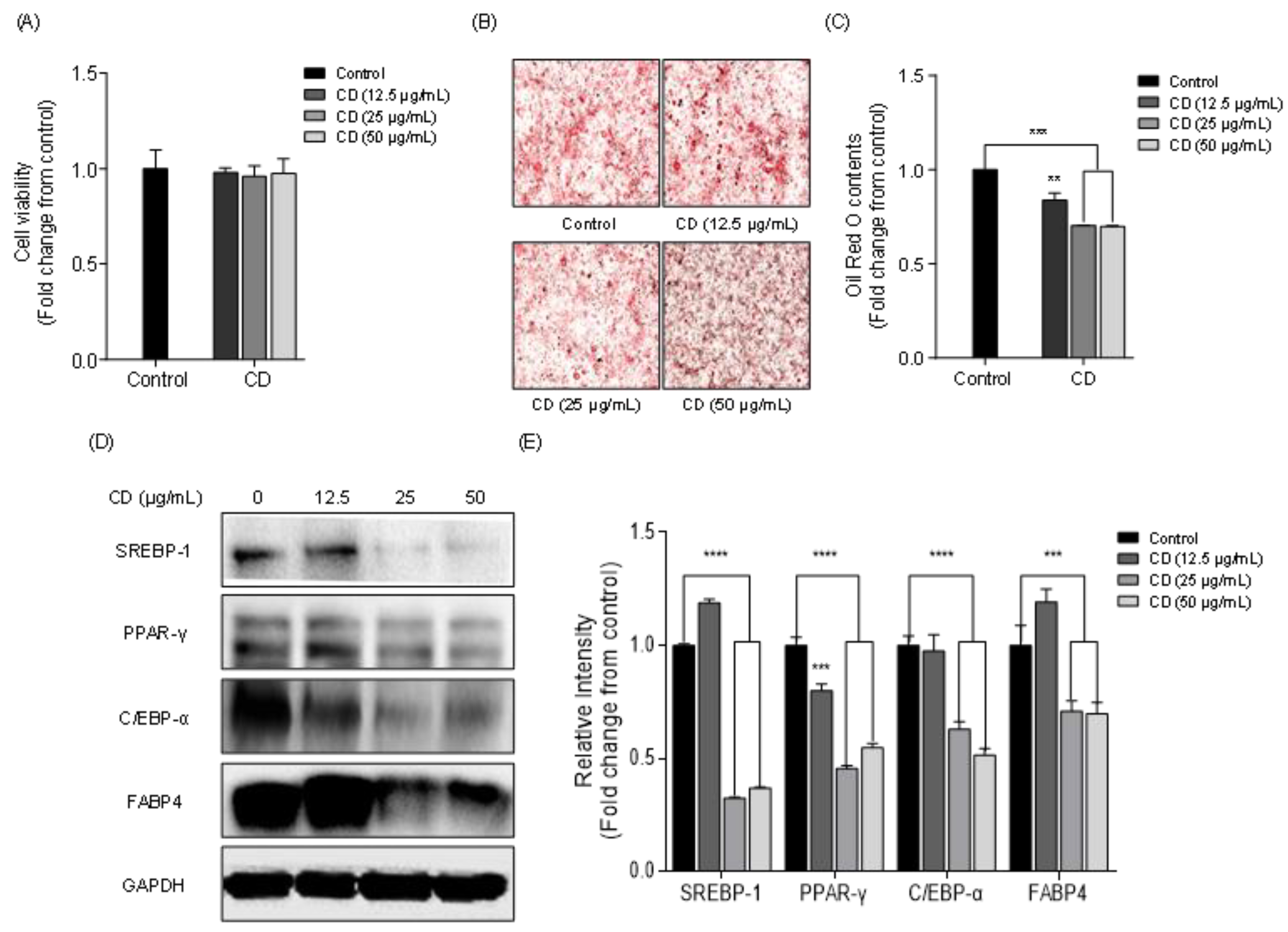
2.5. Effect of Chlorophyll Derivative on Lipid Accumulation and the Expression of Adipogenic Proteins in 3T3-L1 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Extraction and Solvent Fractionation of 60% of Red Seaweeds Ethanol Extract
4.3. HPLC Analysis
4.4. Cell Culture
4.5. Cytotoxicity
4.6. Oil Red O Staining
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| BS | bovine serum |
| DMSO | dimethyl sulfoxide |
| DMEM | dulbecco’s modified Eagle’s medium |
| FABP4 | fatty acid-binding protein 4 |
| FBS | fetal bovine serum |
| GCE | garcinia cambogia extract |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GEE | Grateloupia elliptica ethanol extract |
| H&E | hematoxylin and eosin |
| HFD | high-fat diet |
| ORO | Oil Red O |
| PPAR-γ | peroxisome proliferator-activated receptor gamma |
| SREBP-1 | sterol regulatory element-binding protein 1 |
| TC | total cholesterol; TG, triglyceride |
| CD | chlorophyll derivative |
References
- James, W.P.T. WHO recognition of the global obesity epidemic. Int. J. Obes. 2008, 32, S120–S126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef]
- McDonald, R.; Kuhn, K.; Bradford, A.P.; Santoro, N.J.F. Sterility, Cell type specific effects of hyperlipidemia and hyperinsulinemia, characteristic of reprometabolic syndrome, on pituitary function. Fertil. Steril. 2019, 112, e103. [Google Scholar] [CrossRef]
- Perez-Leighton, C.E.; Lockridge, A.D.; Teske, J.A.; Alejandro, E.U.; Klotz, C.M. Rat Models of Obesity, Metabolic Syndrome, and Diabetes. In The Laboratory Rat; Elsevier: Amsterdam, The Netherlands, 2020; pp. 987–1002. [Google Scholar]
- Bray, G.A.; Popkin, B.M. Dietary fat intake does affect obesity! Am. J. Clin. Nutr. 1998, 68, 1157–1173. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, P.G. Obesity as a medical problem. Nat. Cell Biol. 2000, 404, 635–643. [Google Scholar] [CrossRef]
- Zumoff, B.J.A.M.S. Hormonal abnormalities in obesity. Acta Med. Scand. 1987, 222, 153–160. [Google Scholar] [CrossRef]
- Haluzik, M.; Parizkova, J.; Haluzik, M.M. Adiponectin and its role in the obesity-induced insulin resistance and related complications. Physiol. Res. 2004, 53, 123–129. [Google Scholar]
- Rasouli, N.; Kern, P.A. Adipocytokines and the Metabolic Complications of Obesity. J. Clin. Endocrinol. Metab. 2008, 93, s64–s73. [Google Scholar] [CrossRef] [Green Version]
- Aggoun, Y. Obesity, Metabolic Syndrome, and Cardiovascular Disease. Pediatr. Res. 2007, 61, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, T47–T59. [Google Scholar] [CrossRef] [Green Version]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.; Hamilton, D.; Zderic, T.J.M.S.; Disorders, R. Role of Low Energy Expenditure and Sitting in Obesity, Metabolic Syndrome, Type 2 Diabetes, and Cardiovascular Disease. Diabetes 2007, 56, 2655–2667. [Google Scholar] [CrossRef] [Green Version]
- Chaput, J.P.; St-Pierre, S.; Tremblay, A. Currently available drugs for the treatment of obesity: Sibutramine and orlistat. Mini Rev. Med. Chem. 2007, 7, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Scheen, A.J.; Ernest, P. New antiobesity agents in type 2 diabetes: Overview of clinical trials with sibutramine and orlistat. Diabetes Metab. 2002, 28, 437–445. [Google Scholar] [PubMed]
- Guerciolini, R. Mode of action of orlistat. Int. J. Obes. Relat. Metab. Disord. 1997, 21, S12–S23. [Google Scholar]
- Luque, C.A.; Rey, J.A. Sibutramine: A Serotonin–Norepinephrine Reuptake-Inhibitor for the Treatment of Obesity. Ann. Pharmacother. 1999, 33, 968–978. [Google Scholar] [CrossRef]
- Luque, C.A.; Rey, J.A. The discovery and status of sibutramine as an anti-obesity drug. Eur. J. Pharmacol. 2002, 440, 119–128. [Google Scholar] [CrossRef]
- Abraham, B.; Sellin, J.H. Drug-induced diarrhea. Curr. Gastroenterol. Rep. 2007, 9, 365–372. [Google Scholar] [CrossRef]
- Douglas, I.J.; Langham, J.; Bhaskaran, K.; Brauer, R.; Smeeth, L. Orlistat and the risk of acute liver injury: Self controlled case series study in UK Clinical Practice Research Datalink. BMJ 2013, 346, f1936. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Lee, E.H.; Hwang, J.C.; Jeung, J.H.; Kim, D.Y.; Cheong, J.Y.; Cho, S.W.; Kim, Y.B. A case of acute cholestatic hepatitis associated with Orlistat. Taehan Kan Hakhoe Chi 2002, 8, 317–320. [Google Scholar]
- Montero, J.L.; Muntané, J.; Fraga, E.; Delgado, M.; Costán, G.; Serrano, M.; Padillo, J.; De La Mata, M.; Miño, G. Orlistat associated subacute hepatic failure. J. Hepatol. 2001, 34, 173. [Google Scholar] [CrossRef]
- Sall, D.; Wang, J.; Rashkin, M.; Welch, M.; Droege, C.; Schauer, D. Orlistat-induced fulminant hepatic failure. Clin. Obes. 2014, 4, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Umemura, T.; Ichijo, T.; Matsumoto, A.; Kiyosawa, K. Severe Hepatic Injury Caused by Orlistat. Am. J. Med. 2006, 119, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, W.P.; Caterson, I.D.; Coutinho, W.; Finer, N.; Van Gaal, L.F.; Maggioni, A.P.; Torp-Pedersen, C.; Sharma, A.M.; Shepherd, G.M.; Rode, R.A.; et al. Effect of Sibutramine on Cardiovascular Outcomes in Overweight and Obese Subjects. N. Engl. J. Med. 2010, 363, 905–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, Y.X.; Mi, Y.; Cao, W.X.; Lim, P.E.; Xue, C.H.; Tang, Q.J. A Pilot Study on Anti-Obesity Mechanisms of KappaphycusAlvarezii: The Role of Native κ-Carrageenan and the Leftover Sans-Carrageenan Fraction. Nutrients 2019, 11, 1133. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-C.; Kang, N.; Kim, S.-Y.; Lima, I.S.; Ko, S.-C.; Kim, Y.-T.; Kim, Y.-B.; Jeung, H.-D.; Choi, K.-S.; Jeon, Y.-J. Popular edible seaweed, Gelidium amansii prevents against diet-induced obesity. Food Chem. Toxicol. 2016, 90, 181–187. [Google Scholar] [CrossRef]
- Kang, M.-C.; Kang, N.; Ko, S.-C.; Kim, Y.-B.; Jeon, Y.-J. Anti-obesity effects of seaweeds of Jeju Island on the differentiation of 3T3-L1 preadipocytes and obese mice fed a high-fat diet. Food Chem. Toxicol. 2016, 90, 36–44. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-I.; Kim, H.-G.; Geraldino, P.J.L.; Hwang, I.-K.; Boo, S.-M. Molecular Classification of the Genus Grateloupia (Halymeniaceae, Rhodophyta) in Korea. Algae 2009, 24, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Nam, K.; Kurihara, H.; Kim, S. Potent α-glucosidase inhibitors purified from the red alga Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.I.; Kim, S.C.; Han, S.C.; Hong, H.J.; Jeon, Y.J.; Kim, B.; Koh, Y.S.; Yoo, E.S.; Kang, H.K. Hair-Loss Preventing Effect of Grateloupia elliptica. Biomol. Ther. 2012, 20, 118–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.; Park, G.M.; Kim, S.N.; Amna, T.; Lee, S.; Shin, W.S. Glioblastoma-Specific Anticancer Activity of Pheophorbide a from the Edible Red Seaweed Grateloupia elliptica. J. Microbiol. Biotechnol. 2014, 24, 346–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almela, L.; Fernandez-Lopez, J.A.; Roca, M.J. High-performance liquid chromatographic screening of chlorophyll derivatives produced during fruit storage. J. Chromatogr. A 2000, 870, 483–489. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4. 3.1–F4. 3.8. [Google Scholar] [CrossRef]
- Han, T.S.; Lean, M.E. A clinical perspective of obesity, metabolic syndrome and cardiovascular disease. JRSM Cardiovasc. Dis. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Qin, B.; Poti, J.; Sokol, R.; Gordon-Larsen, P. Epidemiology of Obesity in Adults: Latest Trends. Curr. Obes. Rep. 2018, 7, 276–288. [Google Scholar] [CrossRef]
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The obesity transition: Stages of the global epidemic. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Castro, M.; Preto, M.; Vasconcelos, V.; Urbatzka, R. Obesity: The Metabolic Disease, Advances on Drug Discovery and Natural Product Research. Curr. Top. Med. Chem. 2016, 16, 2577–2604. [Google Scholar] [CrossRef]
- Panwar, B.; Hanks, L.J.; Tanner, R.M.; Muntner, P.; Kramer, H.; McClellan, W.M.; Warnock, D.G.; Judd, S.E.; Gutierrez, O.M. Obesity, metabolic health, and the risk of end-stage renal disease. Kidney Int. 2015, 87, 1216–1222. [Google Scholar] [CrossRef] [Green Version]
- Sakai, N.S.; Taylor, S.A.; Chouhan, M.D. Obesity, metabolic disease and the pancreas—Quantitative imaging of pancreatic fat. Br. J. Radiol. 2018, 91. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Tao, N.; Wang, X.; Xiao, J.; Wang, M. Marine-derived bioactive compounds with anti-obesity effect: A review. J. Funct. Foods 2016, 21, 372–387. [Google Scholar] [CrossRef]
- Awang, A.N.; Ng, J.L.; Matanjun, P.; Sulaiman, M.R.; Tan, T.S.; Ooi, Y.B.H. Anti-obesity property of the brown seaweed, Sargassum polycystum using an in vivo animal model. J. Appl. Phycol. 2014, 26, 1043–1048. [Google Scholar] [CrossRef]
- Ben Abdallah Kolsi, R.; Ben Gara, A.; Chaaben, R.; El Feki, A.; Paolo Patti, F.; El Feki, L.; Belghith, K. Anti-obesity and lipid lowering effects of Cymodocea nodosa sulphated polysaccharide on high cholesterol-fed-rats. Arch. Physiol. Biochem. 2015, 121, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Choung, S.Y. Antiobesity Effects of the Ethanol Extract of Laminaria japonica Areshoung in High-Fat-Diet-Induced Obese Rat. Evid. Based Complement. Altern. Med. 2013, 2013, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-G.; Lu, Y.A.; Li, X.; Hyun, J.-M.; Kim, H.-S.; Lee, J.J.; Kim, T.H.; Kim, H.M.; Kang, M.-C. Anti-Obesity Effects of Grateloupia elliptica, a Red Seaweed, in Mice with High-Fat Diet-Induced Obesity via Suppression of Adipogenic Factors in White Adipose Tissue and Increased Thermogenic Factors in Brown Adipose Tissue. Nutrients 2020, 12, 308. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Antiobesity Effect of Fucoxanthin from Edible Seaweeds and Its Multibiological Functions; ACS Publications: Washington, DC, USA, 2008. [Google Scholar]
- Okada, T.; Mizuno, Y.; Sibayama, S.; Hosokawa, M.; Miyashita, K. Antiobesity Effects of Undaria Lipid Capsules Prepared with Scallop Phospholipids. J. Food Sci. 2011, 76, H2–H6. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine Algae as a Potential Source for Anti-Obesity Agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Chuyen, H.V.; Eun, J.-B. Marine carotenoids: Bioactivities and potential benefits to human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Marine Carotenoids against Oxidative Stress: Effects on Human Health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef]
- Miyashita, K. Function of marine carotenoids. In Food Factors for Health Promotion; Karger Publishers: Basel, Switzerland, 2009; Volume 61, pp. 136–146. [Google Scholar]
- Vílchez, C.; Forján, E.; Cuaresma, M.; Bédmar, F.; Garbayo, I.; Vega, J.M. Marine Carotenoids: Biological Functions and Commercial Applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mounien, L.; Tourniaire, F.; Landrier, J.-F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef] [Green Version]
- Gammone, M.A.; D’Orazio, N. Anti-Obesity Activity of the Marine Carotenoid Fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef]
- Gille, A.; Stojnic, B.; Derwenskus, F.; Trautmann, A.; Schmid-Staiger, U.; Posten, C.; Briviba, K.; Palou, A.; Bonet, M.L.; Ribot, J. A lipophilic fucoxanthin-rich phaeodactylum tricornutum extract ameliorates effects of diet-Induced obesity in C57BL/6J mice. Nutrients 2019, 11, 796. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Takahashi, N.; Kawada, T.; Miyashita, K. Fucoxanthin and its metabolite, fucoxanthinol, suppress adipocyte differentiation in 3T3-L1 cells. Int. J. Mol. Med. 2006, 18, 147–152. [Google Scholar] [CrossRef]
- Freitas, S.; Silva, N.G.; Sousa, M.L.; Ribeiro, T.; Rosa, F.; Leão, P.N.; Vasconcelos, V.; Reis, M.A.; Urbatzka, R. Chlorophyll Derivatives from Marine Cyanobacteria with Lipid-Reducing Activities. Mar. Drugs 2019, 17, 229. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.-J.; Kim, K.-J.; Choi, J.; Koh, E.-J.; Lee, B.-Y. Spirulina maxima extract reduces obesity through suppression of adipogenesis and activation of browning in 3T3-L1 cells and high-fat diet-induced obese mice. Nutrients 2018, 10, 712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christodoulides, C.; Vidal-Puig, A. PPARs and adipocyte function. Mol. Cell. Endocrinol. 2010, 318, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, K.-A.; Hwang, Y.-J.; Im, P.R.; Hwang, H.-J.; Song, J.; Kim, Y.-J. Platycodon grandiflorum Extract Reduces High-Fat Diet-Induced Obesity through Regulation of Adipogenesis and Lipogenesis Pathways in Mice. J. Med. Food 2019, 22, 993–999. [Google Scholar] [CrossRef]
- Ryu, H.-J.; Um, M.Y.; Ahn, J.-Y.; Jung, C.H.; Huh, D.; Kim, T.-W.; Ha, T.-Y. Anti-obesity Effect of Hypsizigus marmoreus in High Fat-fed Mice. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1708–1714. [Google Scholar] [CrossRef]
- Jemai, R.; Drira, R.; Makni, M.; Fetoui, H.; Sakamoto, K. Colocynth (Citrullus colocynthis) seed extracts attenuate adipogenesis by down-regulating PPARγ/SREBP-1c and C/EBPα in 3T3-L1 cells. Food Biosci. 2020, 33, 100491. [Google Scholar] [CrossRef]
- Spiegelman, B.M.; Flier, J.S. Obesity and the Regulation of Energy Balance. Cell 2001, 104, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [PubMed]
- Loftus, T.M.; Lane, M.D. Modulating the transcriptional control of adipogenesis. Curr. Opin. Genet. Dev. 1997, 7, 603–608. [Google Scholar] [CrossRef]
- Jeon, T.; Hwang, S.G.; Hirai, S.; Matsui, T.; Yano, H.; Kawada, T.; Lim, B.O.; Park, D.K. Red yeast rice extracts suppress adipogenesis by down-regulating adipogenic transcription factors and gene expression in 3T3-L1 cells. Life Sci. 2004, 75, 3195–3203. [Google Scholar] [CrossRef]
- Liang, Y.-C.; Yang, M.-T.; Lin, C.-J.; Chang, C.L.-T.; Yang, W.-C. Bidens pilosa and its active compound inhibit adipogenesis and lipid accumulation via down-modulation of the C/EBP and PPARγ pathways. Sci. Rep. 2016, 6, 24285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.-Q.; Zhang, J.-W.; Lane, M.D. Sequential gene promoter interactions of C/EBPβ, C/EBPα, and PPARγ during adipogenesis. Biochem. Biophys. Res. Commun. 2004, 319, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Shah, N.A.; Warrington, J.A.; Anderson, N.N.; Park, S.W.; Brown, M.S.; Goldstein, J.L. Combined analysis of oligonucleotide microarray data from transgenic and knockout mice identifies direct SREBP target genes. Proc. Natl. Acad. Sci. USA 2003, 100, 12027–12032. [Google Scholar] [CrossRef] [Green Version]
- Fajas, L.; Schoonjans, K.; Gelman, L.; Kim, J.B.; Najib, J.; Martin, G.; Fruchart, J.-C.; Briggs, M.; Spiegelman, B.M.; Auwerx, J. Regulation of Peroxisome Proliferator-Activated Receptor γ Expression by Adipocyte Differentiation and Determination Factor 1/Sterol Regulatory Element Binding Protein 1: Implications for Adipocyte Differentiation and Metabolism. Mol. Cell. Biol. 1999, 19, 5495–5503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.B.; Wright, H.M.; Wright, M.; Spiegelman, B.M. ADD1/SREBP1 activates PPARγ through the production of endogenous ligand. Proc. Natl. Acad. Sci. USA 1998, 95, 4333–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, I.S.; Sanjeewa, K.A.; Kim, H.S.; Wang, L.; Lee, W.W.; Jeon, Y.J. Apoptotic and antiproliferative properties of 3β-hydroxy-Δ5-steroidal congeners from a partially purified column fraction of Dendronephthya gigantea against HL-60 and MCF-7 cancer cells. J. Appl. Toxicol. 2018, 38, 527–536. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Scientific Name |
|---|---|
| 1. | Grateloupia elliptica (GE) |
| 2. | Gracilaria vermiculophylla (GVP) |
| 3. | Grateloupia lanceolata (GL) |
| 4. | Gracilaria verrucosa (GV) |
| 5. | Grateloupia asiatica (GA) |
| 6. | Gloiopeltis furcata (GF) |
| 7. | Lomentaria catenata (LC) |
| 8. | Chondrus nipponicus (CN) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-G.; Lu, Y.-A.; Je, J.-G.; Jayawardena, T.U.; Kang, M.-C.; Lee, S.-H.; Kim, T.-H.; Lee, D.-S.; Lee, J.-M.; Yim, M.-J.; et al. Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression. Mar. Drugs 2021, 19, 91. https://doi.org/10.3390/md19020091
Lee H-G, Lu Y-A, Je J-G, Jayawardena TU, Kang M-C, Lee S-H, Kim T-H, Lee D-S, Lee J-M, Yim M-J, et al. Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression. Marine Drugs. 2021; 19(2):91. https://doi.org/10.3390/md19020091
Chicago/Turabian StyleLee, Hyo-Geun, Yu-An Lu, Jun-Geon Je, Thilina U. Jayawardena, Min-Cheol Kang, Seung-Hong Lee, Tae-Hee Kim, Dae-Sung Lee, Jeong-Min Lee, Mi-Jin Yim, and et al. 2021. "Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression" Marine Drugs 19, no. 2: 91. https://doi.org/10.3390/md19020091
APA StyleLee, H. -G., Lu, Y. -A., Je, J. -G., Jayawardena, T. U., Kang, M. -C., Lee, S. -H., Kim, T. -H., Lee, D. -S., Lee, J. -M., Yim, M. -J., Kim, H. -S., & Jeon, Y. -J. (2021). Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression. Marine Drugs, 19(2), 91. https://doi.org/10.3390/md19020091