
Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
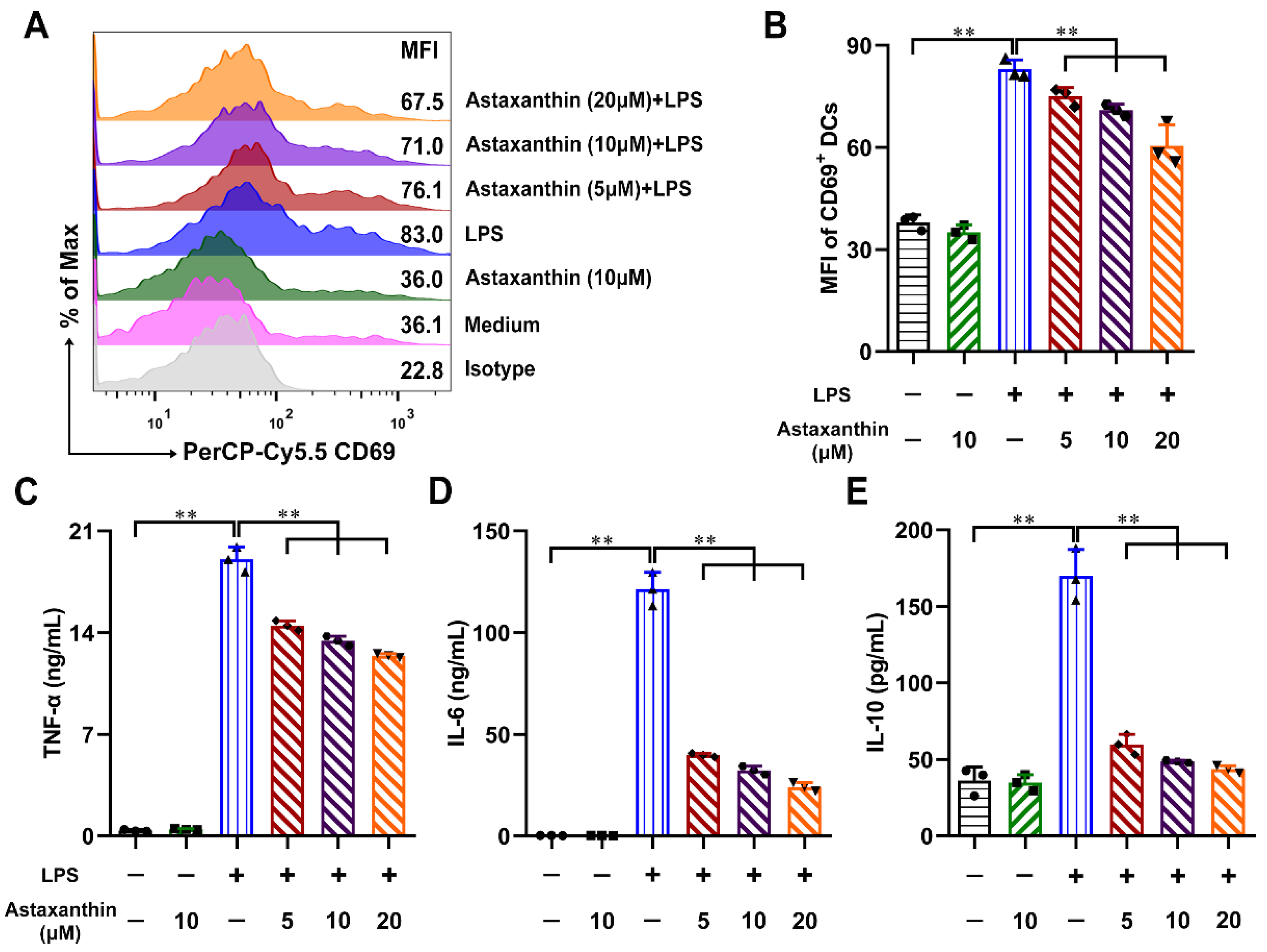
2.1. Astaxanthin Inhibited LPS-Induced Cytokine Production by DCs
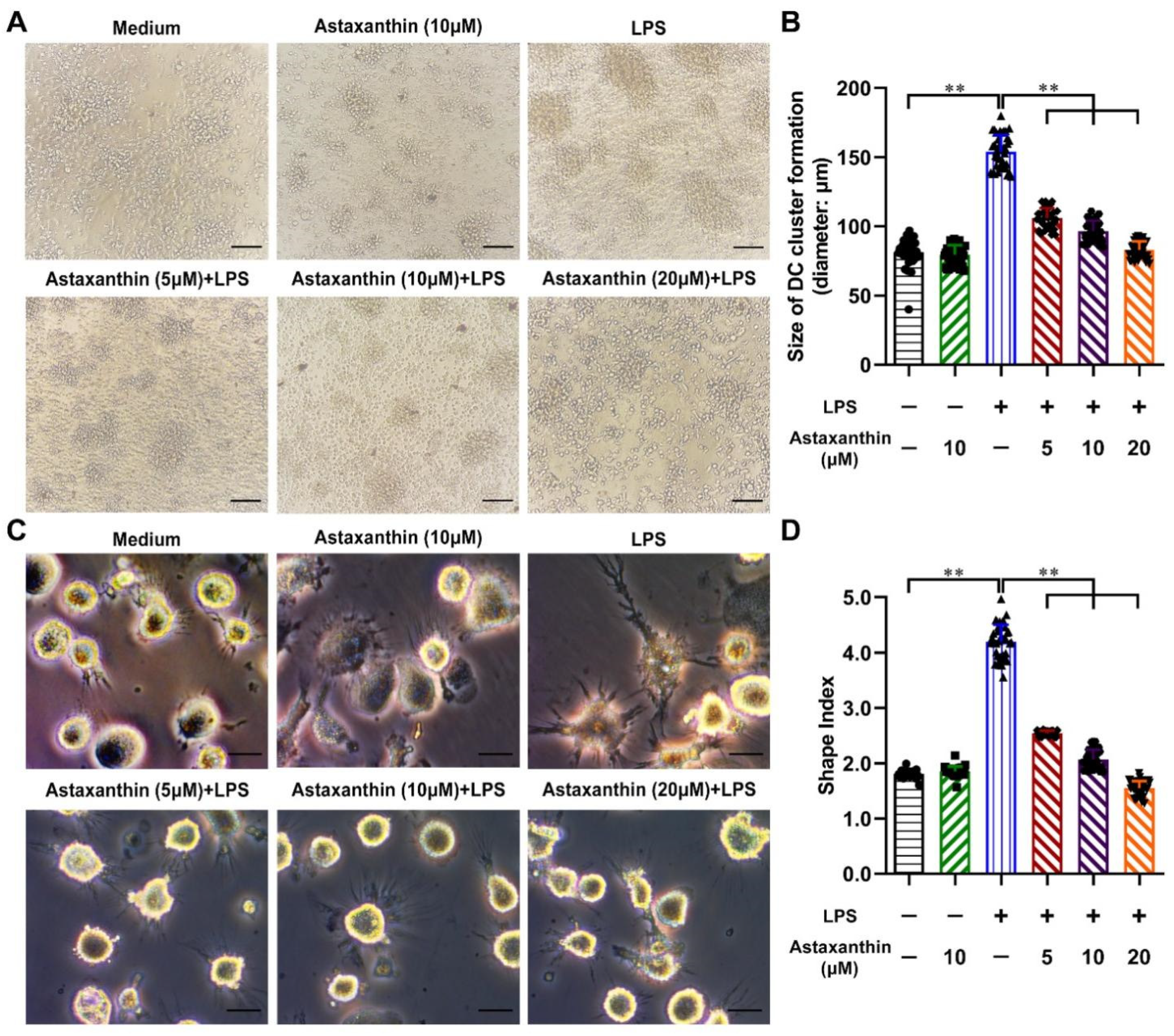
2.2. Astaxanthin Reversed the Morphological Changes in LPS-Activated DCs
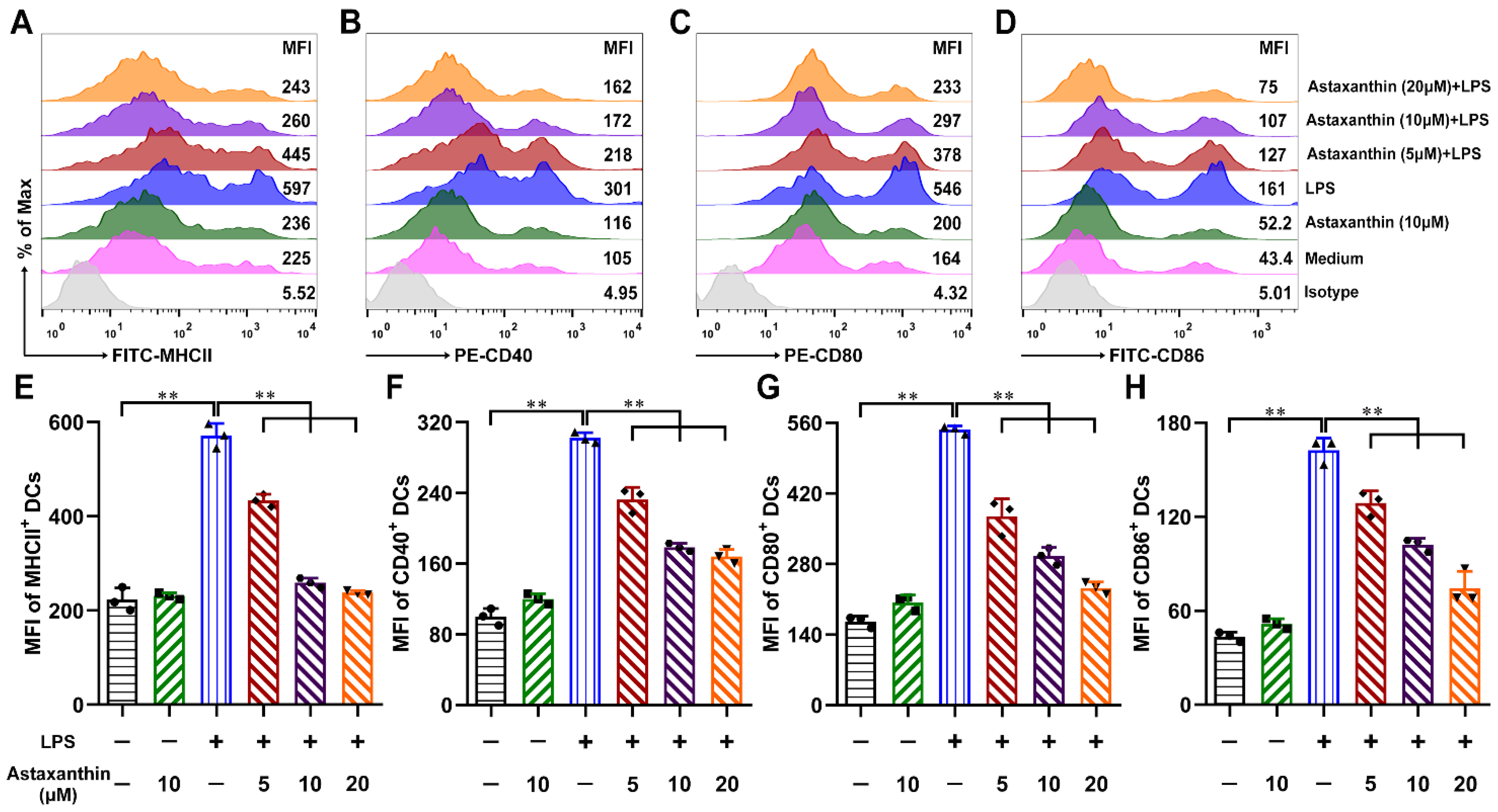
2.3. Astaxanthin Impaired the Phenotypic Maturation of LPS-Induced DCs
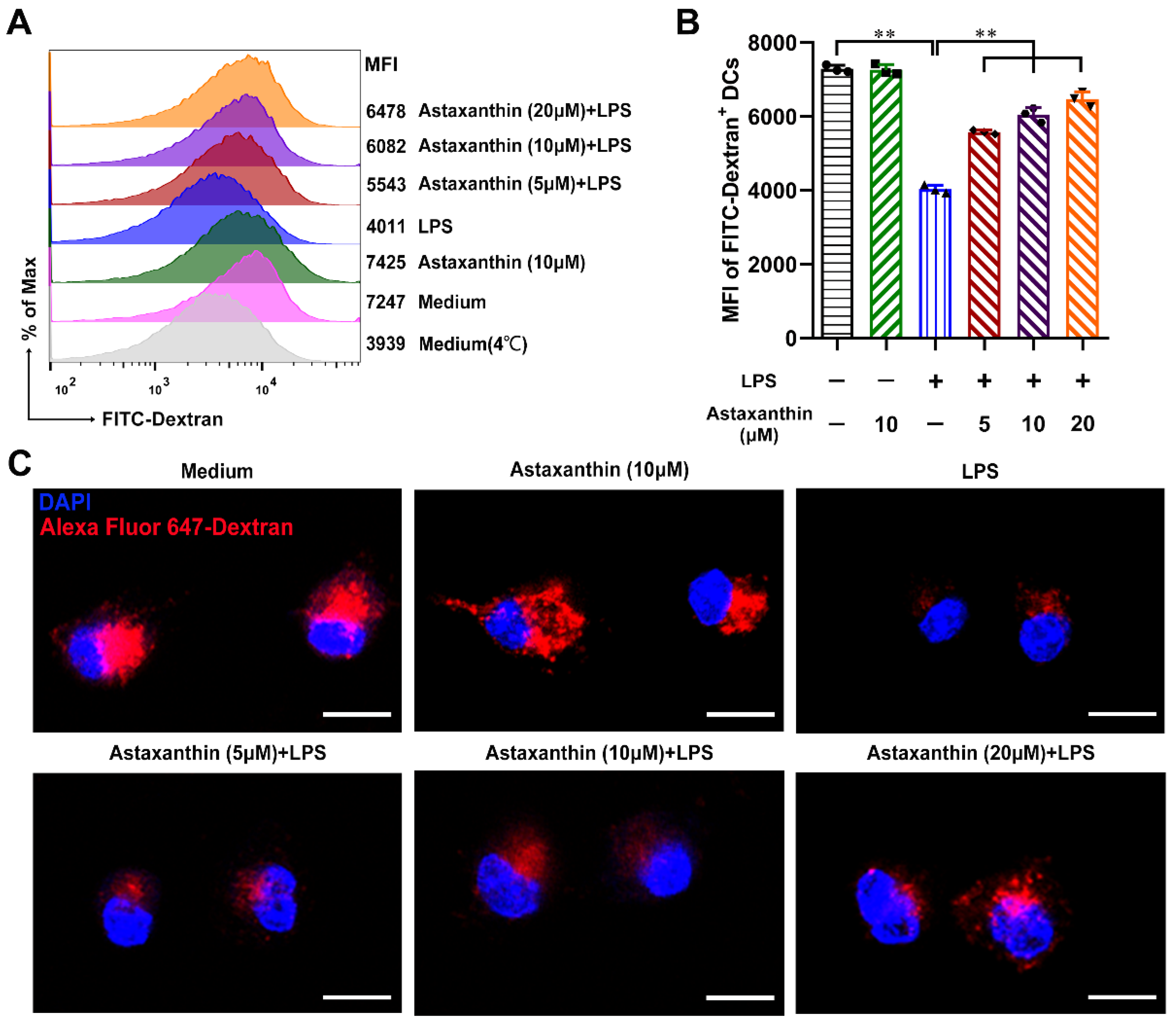
2.4. Astaxanthin Increased the Endocytosis Capability of LPS-Induced DCs
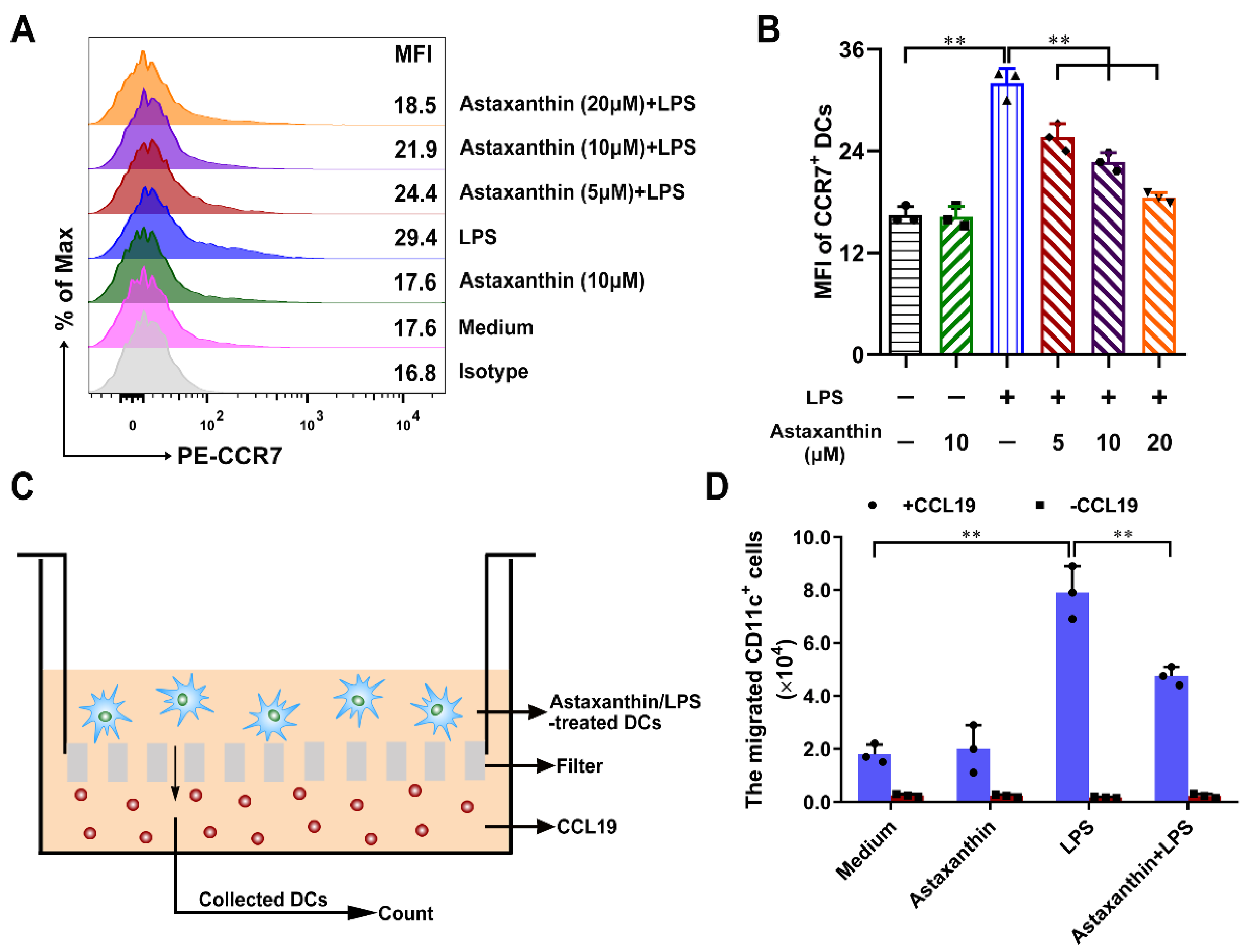
2.5. Astaxanthin Inhibited the Migration Capability of LPS-Induced DCs
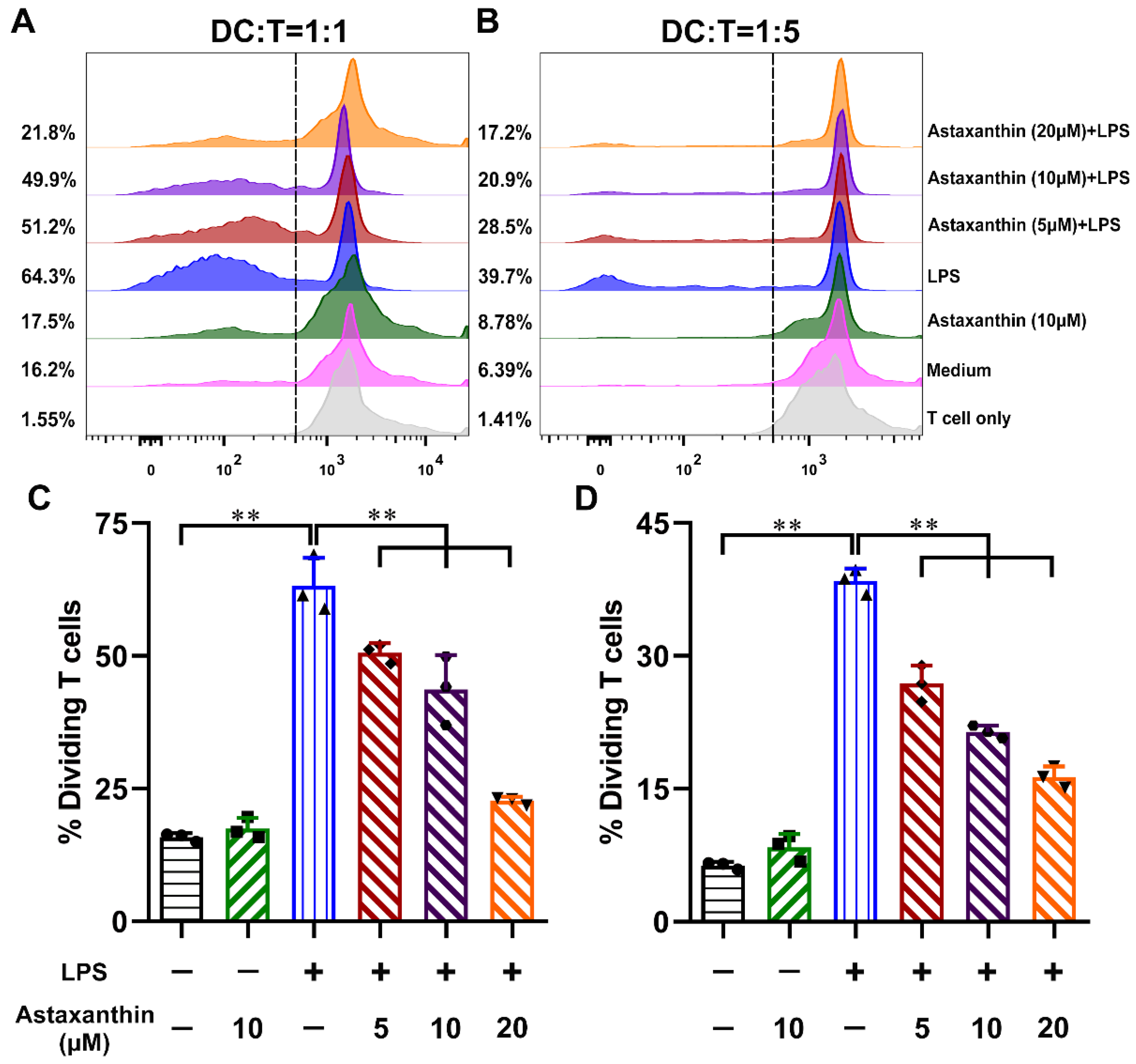
2.6. Astaxanthin Impaired the Allostimulatory Capacity of LPS-Induced DCs
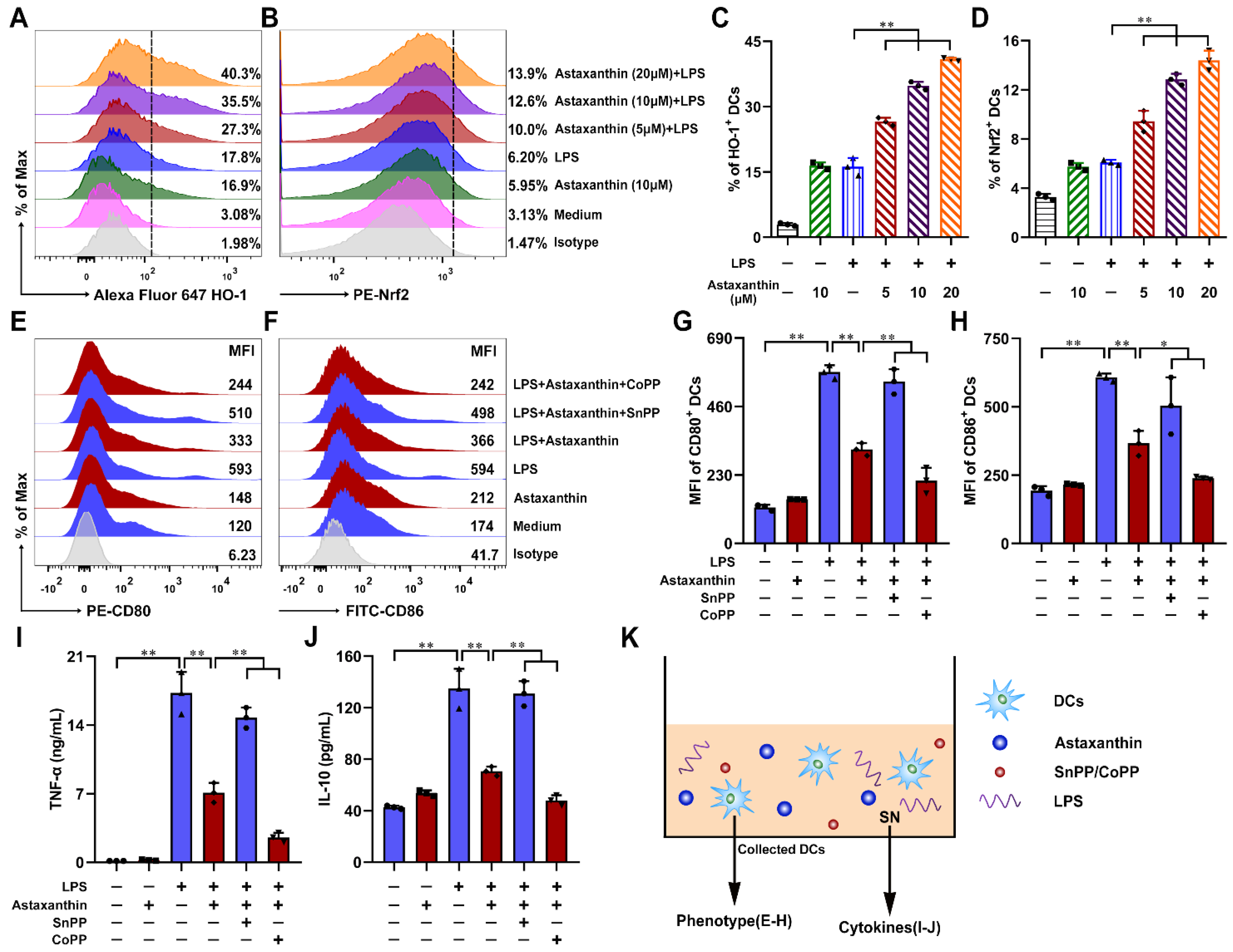
2.7. Astaxanthin Protected the LPS-Induced Immune Dysfunction of DCs via Activation of HO-1/Nrf2 Axis
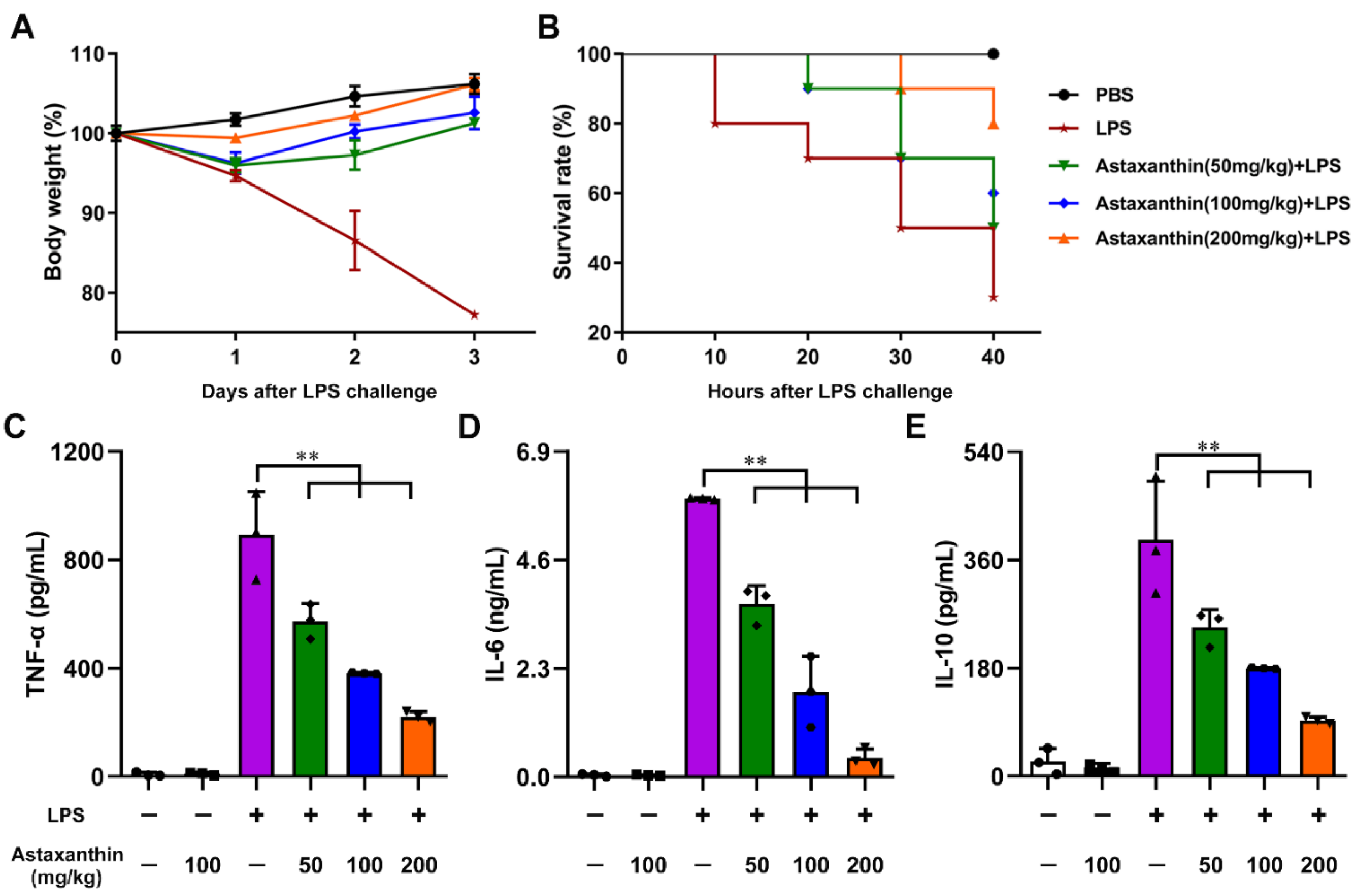
2.8. Astaxanthin Protected LPS-Induced Sepsis in Mice
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Reagents
4.3. Generation of DCs
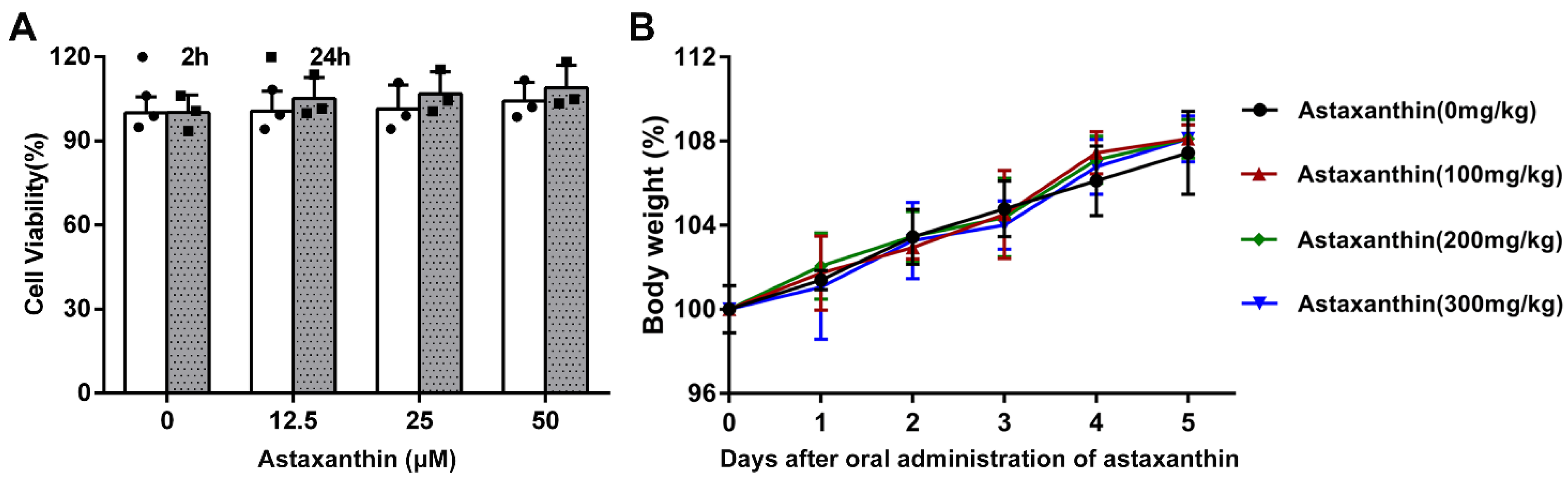
4.4. Cell Viability Assay
4.5. Cytokine Assay
4.6. Phenotype Assay
4.7. Endocytosis Assay
4.8. Migration Assay
4.9. Allogeneic Mixed Lymphocyte Reaction Assay
4.10. HO-1 and Nrf2 Protein Expression Assay
4.11. Body Weight Change Assay
4.12. Survival Rate and Cytokine Assay
4.13. Statistical Analysis
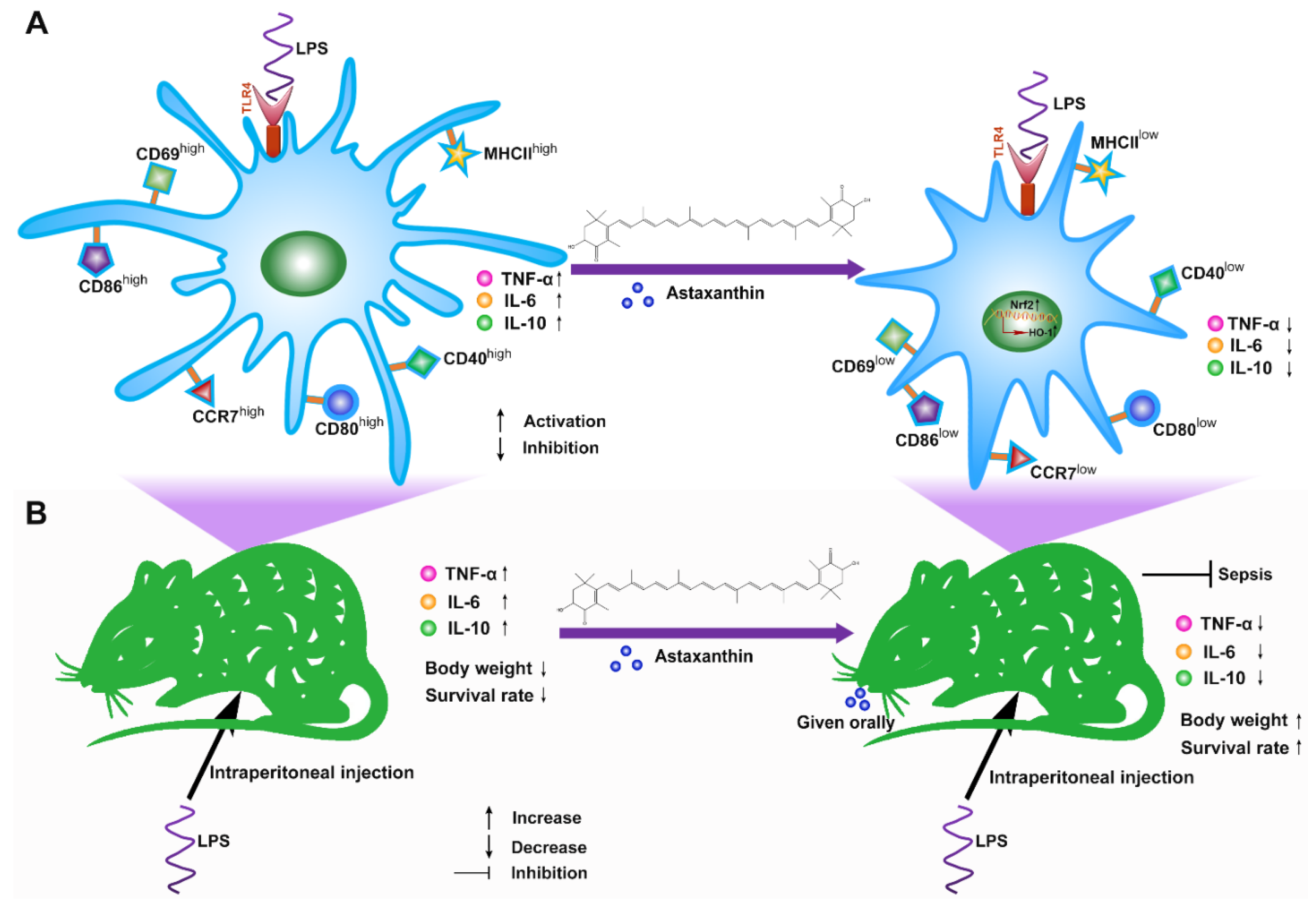
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Crimeen-Irwin, B.; Scalzo, K.; Gloster, S.; Mottram, P.L.; Plebanski, M. Failure of immune homeostasis—The consequences of under and over reactivity. Curr. Drug Targets Immune Endocr. Metab. Disord. 2005, 5, 413–422. [Google Scholar] [CrossRef]
- Horwitz, D.A.; Fahmy, T.M.; Piccirillo, C.A.; La Cava, A. Rebalancing Immune Homeostasis to Treat Autoimmune Diseases. Trends Immunol. 2019, 40, 888–908. [Google Scholar] [CrossRef]
- Fleischmann, C.; Scherag, A.; Adhikari, N.K.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K.; International Forum of Acute Care Trialists. Assessment of Global Incidence and Mortality of Hospital-treated Sepsis. Current Estimates and Limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef]
- Liao, X.; Du, B.; Lu, M.; Wu, M.; Kang, Y. Current epidemiology of sepsis in mainland China. Ann. Transl. Med. 2016, 4, 324. [Google Scholar] [CrossRef] [Green Version]
- van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef]
- Baccala, R.; Gonzalez-Quintial, R.; Lawson, B.R.; Stern, M.E.; Kono, D.H.; Beutler, B.; Theofilopoulos, A.N. Sensors of the innate immune system: Their mode of action. Nat. Rev. Rheumatol. 2009, 5, 448–456. [Google Scholar] [CrossRef]
- Steinhagen, F.; Schmidt, S.V.; Schewe, J.C.; Peukert, K.; Klinman, D.M.; Bode, C. Immunotherapy in sepsis—Brake or accelerate? Pharm. Ther. 2020, 208, 107476. [Google Scholar] [CrossRef] [PubMed]
- Delano, M.J.; Ward, P.A. Sepsis-induced immune dysfunction: Can immune therapies reduce mortality? J. Clin. Investig. 2016, 126, 23–31. [Google Scholar] [CrossRef]
- Yin, Y.; Qin, T.; Yu, Q.; Yang, Q. Bursopentin (BP5) from chicken bursa of fabricius attenuates the immune function of dendritic cells. Amino Acids 2014, 46, 1763–1774. [Google Scholar] [CrossRef]
- Zhong, W.; Qian, K.; Xiong, J.; Ma, K.; Wang, A.; Zou, Y. Curcumin alleviates lipopolysaccharide induced sepsis and liver failure by suppression of oxidative stress-related inflammation via PI3K/AKT and NF-kappaB related signaling. Biomed. Pharm. 2016, 83, 302–313. [Google Scholar] [CrossRef]
- Alsharif, K.F.; Almalki, A.A.; Al-Amer, O.; Mufti, A.H.; Theyab, A.; Lokman, M.S.; Ramadan, S.S.; Almeer, R.S.; Hafez, M.M.; Kassab, R.B.; et al. Oleuropein protects against lipopolysaccharide-induced sepsis and alleviates inflammatory responses in mice. IUBMB Life 2020, 72, 2121–2132. [Google Scholar] [CrossRef]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef]
- Gallo, P.M.; Gallucci, S. The dendritic cell response to classic, emerging, and homeostatic danger signals. Implications for autoimmunity. Front. Immunol. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Tisch, R. Immunogenic versus tolerogenic dendritic cells: A matter of maturation. Int. Rev. Immunol. 2010, 29, 111–118. [Google Scholar] [CrossRef]
- Wu, D.D.; Li, T.; Ji, X.Y. Dendritic Cells in Sepsis: Pathological Alterations and Therapeutic Implications. J. Immunol. Res. 2017, 2017, 3591248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Liu, Z.; Jin, H.; Yan, J.; Liang, H.P. Alterations of dendritic cells in sepsis: Featured role in immunoparalysis. Biomed. Res. Int. 2015, 2015, 903720. [Google Scholar] [CrossRef] [PubMed]
- Oberholzer, A.; Oberholzer, C.; Efron, P.A.; Scumpia, P.O.; Uchida, T.; Bahjat, K.; Ungaro, R.; Tannahill, C.L.; Murday, M.; Bahjat, F.R.; et al. Functional modification of dendritic cells with recombinant adenovirus encoding interleukin 10 for the treatment of sepsis. Shock 2005, 23, 507–515. [Google Scholar] [PubMed]
- Oberholzer, A.; Oberholzer, C.; Bahjat, K.S.; Ungaro, R.; Tannahill, C.L.; Murday, M.; Bahjat, F.R.; Abouhamze, Z.; Tsai, V.; LaFace, D.; et al. Increased survival in sepsis by in vivo adenovirus-induced expression of IL-10 in dendritic cells. J. Immunol. 2002, 168, 3412–3418. [Google Scholar] [CrossRef]
- Qin, T.; Yin, Y.; Yu, Q.; Yang, Q. Bursopentin (BP5) protects dendritic cells from lipopolysaccharide-induced oxidative stress for immunosuppression. PLoS ONE 2015, 10, e0117477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, Y.; Yoshida, H.; Kondo, K. Potential Anti-Atherosclerotic Properties of Astaxanthin. Mar. Drugs 2016, 14, 35. [Google Scholar] [CrossRef]
- Li, C.; Ma, B.; Chen, J.; Jeong, Y.; Xu, X. Astaxanthin Inhibits p70 S6 Kinase 1 Activity to Sensitize Insulin Signaling. Mar. Drugs 2020, 18, 495. [Google Scholar] [CrossRef]
- Augusti, P.R.; Quatrin, A.; Somacal, S.; Conterato, G.M.; Sobieski, R.; Ruviaro, A.R.; Maurer, L.H.; Duarte, M.M.; Roehrs, M.; Emanuelli, T. Astaxanthin prevents changes in the activities of thioredoxin reductase and paraoxonase in hypercholesterolemic rabbits. J. Clin. Biochem. Nutr. 2012, 51, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bae, M.; Kim, B.; Park, Y.K.; Koo, S.I.; Lee, J.Y. Astaxanthin prevents and reverses the activation of mouse primary hepatic stellate cells. J. Nutr. Biochem. 2016, 29, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, T.; Shimazawa, M.; Inoue, Y.; Nakano, Y.; Ojino, K.; Izawa, H.; Tsuruma, K.; Ishibashi, T.; Hara, H. Astaxanthin Protects Against Retinal Damage: Evidence from In Vivo and In Vitro Retinal Ischemia and Reperfusion Models. Curr. Eye Res. 2016, 41, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Farruggia, C.; Ku, C.S.; Pham, T.X.; Yang, Y.; Bae, M.; Wegner, C.J.; Farrell, N.J.; Harness, E.; Park, Y.K.; et al. Astaxanthin inhibits inflammation and fibrosis in the liver and adipose tissue of mouse models of diet-induced obesity and nonalcoholic steatohepatitis. J. Nutr. Biochem. 2017, 43, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Wu, C.; Kim, J.; Kim, B.; Lee, S.J. Astaxanthin reduces hepatic lipid accumulations in high-fat-fed C57BL/6J mice via activation of peroxisome proliferator-activated receptor (PPAR) alpha and inhibition of PPAR gamma and Akt. J. Nutr. Biochem. 2016, 28, 9–18. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H. Multiple Mechanisms of Anti-Cancer Effects Exerted by Astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.R.; Gough, L.A.; Deb, S.K.; Sparks, S.A.; McNaughton, L.R. Astaxanthin in Exercise Metabolism, Performance and Recovery: A Review. Front. Nutr. 2017, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Fang, Q.; Guo, S.; Zhou, H.; Han, R.; Wu, P.; Han, C. Astaxanthin protects against early burn-wound progression in rats by attenuating oxidative stress-induced inflammation and mitochondria-related apoptosis. Sci. Rep. 2017, 7, 41440. [Google Scholar] [CrossRef]
- Ying, C.J.; Zhang, F.; Zhou, X.Y.; Hu, X.T.; Chen, J.; Wen, X.R.; Sun, Y.; Zheng, K.Y.; Tang, R.X.; Song, Y.J. Anti-inflammatory Effect of Astaxanthin on the Sickness Behavior Induced by Diabetes Mellitus. Cell. Mol. Neurobiol. 2015, 35, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Lu, J.W.; Liu, F.C.; Lee, C.H.; Lee, H.S.; Ho, Y.J.; Hsieh, T.H.; Wu, C.C.; Wang, C.C. Astaxanthin attenuates joint inflammation induced by monosodium urate crystals. FASEB J. 2020, 34, 11215–11226. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chen, Y.; Xie, X.; Yao, D.; Ding, C.; Chen, M. Astaxanthin prevents against lipopolysaccharide-induced acute lung injury and sepsis via inhibiting activation of MAPK/NF-κB. Am. J. Transl. Res. 2019, 11, 1884–1894. [Google Scholar]
- Zeng, X.; Wang, T.; Zhu, C.; Xing, X.; Ye, Y.; Lai, X.; Song, B.; Zeng, Y. Topographical and biological evidence revealed FTY720-mediated anergy-polarization of mouse bone marrow-derived dendritic cells in vitro. PLoS ONE 2012, 7, e34830. [Google Scholar] [CrossRef]
- Platt, C.D.; Ma, J.K.; Chalouni, C.; Ebersold, M.; Bou-Reslan, H.; Carano, R.A.; Mellman, I.; Delamarre, L. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 4287–4292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagihara, S.; Komura, E.; Nagafune, J.; Watarai, H.; Yamaguchi, Y. EBI1/CCR7 is a new member of dendritic cell chemokine receptor that is up-regulated upon maturation. J. Immunol. 1998, 161, 3096–3102. [Google Scholar] [PubMed]
- Freudenthal, P.S.; Steinman, R.M. The distinct surface of human blood dendritic cells, as observed after an improved isolation method. Proc. Natl. Acad. Sci. USA 1990, 87, 7698–7702. [Google Scholar] [CrossRef] [Green Version]
- Bosmann, M.; Ward, P.A. The inflammatory response in sepsis. Trends Immunol. 2013, 34, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.Y.; Koyama-Nasu, R.; Yagi, R.; Nakayama, T. A new therapeutic target: The CD69-Myl9 system in immune responses. Semin. Immunopathol. 2019, 41, 349–358. [Google Scholar] [CrossRef]
- Alari-Pahissa, E.; Notario, L.; Lorente, E.; Vega-Ramos, J.; Justel, A.; Lopez, D.; Villadangos, J.A.; Lauzurica, P. CD69 does not affect the extent of T cell priming. PLoS ONE 2012, 7, e48593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Yang, W.; Shi, X.; Du, P.; Su, L.; Qin, Z.; Chen, J.; Deng, H. TNF receptor 1 mediates dendritic cell maturation and CD8 T cell response through two distinct mechanisms. J. Immunol. 2011, 187, 1184–1191. [Google Scholar] [CrossRef]
- Trevejo, J.M.; Marino, M.W.; Philpott, N.; Josien, R.; Richards, E.C.; Elkon, K.B.; Falck-Pedersen, E. TNF-alpha -dependent maturation of local dendritic cells is critical for activating the adaptive immune response to virus infection. Proc. Natl. Acad. Sci. USA 2001, 98, 12162–12167. [Google Scholar] [CrossRef] [Green Version]
- Hibbert, J.E.; Currie, A.; Strunk, T. Sepsis-Induced Immunosuppression in Neonates. Front. Pediatr. 2018, 6, 357. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.S.; Gad, M. Differential induction of inflammatory cytokines by dendritic cells treated with novel TLR-agonist and cytokine based cocktails: Targeting dendritic cells in autoimmunity. J. Inflamm. 2010, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Zhang, L.; Ling, F.; Wen, S.; Luo, Y.; Liu, H.; Liu, J.; Zheng, W.; Liang, M.; Sun, J.; et al. Effect of immune tolerance induced by immature dendritic cells and CTLA4-Ig on systemic lupus erythematosus: An in vivo study. Exp. Ther. Med. 2018, 15, 2499–2506. [Google Scholar] [CrossRef] [PubMed]
- Mok, M.Y. Tolerogenic dendritic cells: Role and therapeutic implications in systemic lupus erythematosus. Int. J. Rheum. Dis. 2015, 18, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, D.; Vollmann, E.H.; von Andrian, U.H. Mechanisms and consequences of dendritic cell migration. Immunity 2008, 29, 325–342. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yu, S.; Chen, J.D.; Kong, A.N. The nuclear cofactor RAC3/AIB1/SRC-3 enhances Nrf2 signaling by interacting with transactivation domains. Oncogene 2013, 32, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Otterbein, L.E.; Soares, M.P.; Yamashita, K.; Bach, F.H. Heme oxygenase-1: Unleashing the protective properties of heme. Trends Immunol. 2003, 24, 449–455. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef]
- Cohen, J. The immunopathogenesis of sepsis. Nature 2002, 420, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Toner, P.; McAuley, D.F.; Shyamsundar, M. Aspirin as a potential treatment in sepsis or acute respiratory distress syndrome. Crit. Care 2015, 19, 374. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Qin, T.; Wang, X.; Lin, J.; Yu, Q.; Yang, Q. CpG DNA assists the whole inactivated H9N2 influenza virus in crossing the intestinal epithelial barriers via transepithelial uptake of dendritic cell dendrites. Mucosal Immunol. 2015, 8, 799–814. [Google Scholar] [CrossRef] [Green Version]
- Lutz, M.B.; Assmann, C.U.; Girolomoni, G.; Ricciardi-Castagnoli, P. Different cytokines regulate antigen uptake and presentation of a precursor dendritic cell line. Eur. J. Immunol. 1996, 26, 586–594. [Google Scholar] [CrossRef]
- Qin, T.; Ma, S.; Miao, X.; Tang, Y.; Huangfu, D.; Wang, J.; Jiang, J.; Xu, N.; Yin, Y.; Chen, Y.; et al. Mucosal Vaccination for Influenza Protection Enhanced by Catalytic Immune-Adjuvant. Adv. Sci. 2020, 7, 2000771. [Google Scholar] [CrossRef]
- Marton, A.; Kolozsi, C.; Kusz, E.; Olah, Z.; Letoha, T.; Vizler, C.; Pecze, L. Propylene-glycol aggravates LPS-induced sepsis through production of TNF-alpha and IL-6. Iran. J. Immunol. 2014, 11, 113–122. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Y.; Xu, N.; Shi, Y.; Zhou, B.; Sun, D.; Ma, B.; Xu, Z.; Yang, J.; Li, C. Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction. Mar. Drugs 2021, 19, 346. https://doi.org/10.3390/md19060346
Yin Y, Xu N, Shi Y, Zhou B, Sun D, Ma B, Xu Z, Yang J, Li C. Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction. Marine Drugs. 2021; 19(6):346. https://doi.org/10.3390/md19060346
Chicago/Turabian StyleYin, Yinyan, Nuo Xu, Yi Shi, Bangyue Zhou, Dongrui Sun, Bixia Ma, Zhengzhong Xu, Jin Yang, and Chunmei Li. 2021. "Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction" Marine Drugs 19, no. 6: 346. https://doi.org/10.3390/md19060346
APA StyleYin, Y., Xu, N., Shi, Y., Zhou, B., Sun, D., Ma, B., Xu, Z., Yang, J., & Li, C. (2021). Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction. Marine Drugs, 19(6), 346. https://doi.org/10.3390/md19060346