
Seaweed Components as Potential Modulators of the Gut Microbiota



Abstract
:
1. Introduction
2. Discussion
2.1. Polysaccharides
2.1.1. Fucoidans
2.1.2. Laminarin
2.1.3. Alginate
2.1.4. Carrageenans
2.1.5. Porphyran
2.1.6. Ulvans
2.2. Gastrointestinal Digestion Studies with Seaweed Polysaccharides
2.3. Polyphenols
2.3.1. Bromophenols
2.3.2. Phlorotannins
2.4. In Vitro and In Vivo Gastrointestinal Digestion Studies with Seaweed Polyphenols
2.4.1. Prebiotic Function and Attenuation of Metabolic Syndrome by Phlorotannins
2.4.2. Impact of Digestion on Phlorotannin Bioactivity, Attenuation of DNA Damage, and Cancer Cell Proliferation In Vitro
2.5. Seaweed-Derived Peptides
2.6. Gastrointestinal Digestion Studies with Seaweed Peptides
Modulation of Intestinal Epithelial Cell Differentiation
2.7. Bioaccessibility and Bioavailability
2.7.1. In Vitro Bioaccessibility Methods
Solubility and Dialysability
Static and Dynamic Gastrointestinal Models
Cell Models
2.7.2. Ex Vivo Bioavailability Methods
2.7.3. In Vitro Fermentation Models
2.7.4. In Vivo Bioavailability Methods
2.7.5. Bioaccessibility of Seaweed Polysaccharides
2.7.6. Bioaccessibilty of Seaweed Polyphenols
2.7.7. Bioaccessibility of Seaweed Peptides
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Feng, W.; Hu, Y.; An, N.; Feng, Z.; Liu, J.; Mou, J.; Hu, T.; Guan, H.; Zhang, D.; Mao, Y. Alginate oligosaccharide alleviates monocrotaline-induced pulmonary hypertension via anti-oxidant and anti-inflammation pathways in rats. Int. Heart J. 2020, 61, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotas, J.; Marques, V.; Afonso, M.B.; Rodrigues, C.M.; Pereira, L. Antitumour potential of Gigartina pistillata carrageenans against colorectal cancer stem cell-enriched tumourspheres. Mar. Drugs 2020, 18, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, L.V.; Parada, J.; Pérez-Correa, J.R.; Mariotti-Celis, M.S.; Erpel, F.; Zambrano, A.; Palacios, M. Bioactive polyphenols from southern Chile seaweed as inhibitors of enzymes for starch digestion. Mar. Drugs 2020, 18, 353. [Google Scholar] [CrossRef]
- Lee, H.-G.; Lu, Y.A.; Li, X.; Hyun, J.-M.; Kim, H.-S.; Lee, J.J.; Kim, T.H.; Kim, H.M.; Kang, M.-C. Anti-obesity effects of Grateloupia elliptica, a red seaweed, in mice with high-fat diet-induced obesity via suppression of adipogenic factors in white adipose tissue and increased thermogenic factors in brown adipose tissue. Nutrients 2020, 12, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimentel, F.B.; Cermeño, M.; Kleekayai, T.; Harnedy, P.A.; FitzGerald, R.J.; Alves, R.C.; Oliveira, M.B.P. Effect of in vitro simulated gastrointestinal digestion on the antioxidant activity of the red seaweed Porphyra dioica. Food Res. Int. 2020, 136, 109309. [Google Scholar] [CrossRef] [PubMed]
- Irwin, Z.; McSorley, E.M.; Slevin, M.M.; Rowan, L.; McMillen, P.; McCullagh, D.; Magee, P.J.; Gill, C.I.; Cherry, P.; Crowe, W. The effect of a fibre extract from the red seaweed, Palmaria palmata, on lipid metabolism and inflammation in healthy adults. Proc. Nutr. Soc. 2020, 79. [Google Scholar] [CrossRef]
- Seca, A.M.; Pinto, D.C. Overview on the antihypertensive and anti-obesity effects of secondary metabolites from seaweeds. Mar. Drugs 2018, 16, 237. [Google Scholar] [CrossRef] [Green Version]
- Collins, K.G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Looking beyond the terrestrial: The potential of seaweed derived bioactives to treat non-communicable diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vaquero, M.; Mora, L.; Hayes, M. In vitro and in silico approaches to generating and identifying angiotensin-converting enzyme I inhibitory peptides from green macroalga Ulva lactuca. Mar. Drugs 2019, 17, 204. [Google Scholar] [CrossRef] [Green Version]
- Charoensiddhi, S.; Conlon, M.A.; Vuaran, M.S.; Franco, C.M.M.; Zhang, W. Polysaccharide and phlorotannin-enriched extracts of the brown seaweed Ecklonia radiata influence human gut microbiota and fermentation in vitro. J. Appl. Phycol. 2017, 29, 2407–2416. [Google Scholar] [CrossRef]
- Dobrinčić, A.; Balbino, S.; Zorić, Z.; Pedisić, S.; Bursać Kovačević, D.; Elez Garofulić, I.; Dragović-Uzelac, V. Advanced technologies for the extraction of marine brown algal polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Hao, T. Research advances on the chemical structures and medicinal values of seaweed polysaccharides. J. Anhui Agric. Sci. 2018, 2018, 14. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-AHNY201810014.htm (accessed on 11 May 2020).
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Biological activities and potential cosmeceutical applications of bioactive components from brown seaweeds: A review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current trends on seaweeds: Looking at chemical composition, phytopharmacology, and cosmetic applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pagès, A.K.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed secondary metabolites with beneficial health effects: An overview of successes in in vivo studies and clinical trials. Mar. Drugs 2019, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Marzullo, P.; Di Renzo, L.; Pugliese, G.; De Siena, M.; Barrea, L.; Muscogiuri, G.; Colao, A.; Savastano, S. From obesity through gut microbiota to cardiovascular diseases: A dangerous journey. Int. J. Obes. Suppl. 2020, 10, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Barko, P.C.; McMichael, M.A.; Swanson, K.S.; Williams, D.A. The gastrointestinal microbiome: A Review. J. Vet. Intern. Med. 2018, 32, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: Composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J. Chapter 6-Variability and stability of the human gut microbiome. In Why Gut Microbes Matter: Understanding Our Microbiome; Flint, H., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 63–79. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Özgün, C.O.U.; Knut, R.; Dzung, B.D. Modulation of the gut microbiota by prebiotic fibres and bacteriocins. Microb. Ecol. Health Dis. 2017, 28, 1348886. [Google Scholar] [CrossRef] [Green Version]
- Busnelli, M.; Manzini, S. The gut microbiota affects host pathophysiology as an endocrine organ: A focus on cardiovascular disease. Nutrients 2020, 12, 79. [Google Scholar] [CrossRef] [Green Version]
- Markowiak-Kopeć, P.; Śliżewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal relationship between diet-induced gut microbiota changes and diabetes: A novel strategy to transplant Faecalibacterium prausnitzii in preventing diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Rowin, J.; Xia, Y.; Jung, B.; Sun, J. Gut inflammation and dysbiosis in human motor neuron disease. Physiol. Rep. 2017, 5, e13443. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensiddhi, S.; Abraham, R.E.; Su, P.; Zhang, W. Chapter Four-Seaweed and seaweed-derived metabolites as prebiotics. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 91, pp. 97–156. [Google Scholar] [CrossRef]
- Hu, B.; Gong, Q.; Wang, Y.; Ma, Y.; Li, J.; Yu, W. Prebiotic effects of neoagaro-oligosaccharides prepared by enzymatic hydrolysis of agarose. Anaerobe 2006, 12, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Parra-Saldivar, R.; Bilal, M.; Afroze, C.A.; Ahmed, M.N.; Iqbal, H.M.N.; Xu, J. Algae-derived bioactive molecules for the potential treatment of SARS-CoV-2. Molecules 2021, 26, 2134. [Google Scholar] [CrossRef]
- Ajanth Praveen, M.; Karthika Parvathy, K.R.; Jayabalan, R.; Balasubramanian, P. Dietary fiber from Indian edible seaweeds and its in-vitro prebiotic effect on the gut microbiota. Food Hydrocoll. 2019, 96, 343–353. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Quigley, E.M.M. Prebiotics and probiotics in digestive health. Clin. Gastroenterol. Hepatol. 2019, 17, 333–344. [Google Scholar] [CrossRef]
- Coelho, M.C.; Ribeiro, T.B.; Oliveira, C.; Batista, P.; Castro, P.; Monforte, A.R.; Rodrigues, A.S.; Teixeira, J.; Pintado, M. In vitro gastrointestinal digestion impact on the bioaccessibility and antioxidant capacity of bioactive compounds from tomato flours obtained after conventional and ohmic heating extraction. Foods 2021, 10, 554. [Google Scholar] [CrossRef]
- Neal, M.J. Chap 3-Drug absorption, distribution and excretion In Medical Pharmacology at a Glance, 9th ed.; Neal, M.J., Ed.; John Wiley & Sons: Oxford, UK, 2020; Available online: https://www.wiley.com/en-au/Medical+Pharmacology+at+a+Glance%2C+9th+Edition-p-9781119548096 (accessed on 15 May 2020).
- Srinivasan, V.S. Bioavailability of nutrients: A practical approach to in vitro demonstration of the availability of nutrients in multivitamin-mineral combination products. J. Nutr. 2001, 131, 1349S–1350S. [Google Scholar] [CrossRef]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. Chapter 2-Methods for determining bioavailability and bioaccessibility of bioactive compounds and nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Barba, F.J., Saraiva, J.M.A., Cravotto, G., Lorenzo, J.M., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 23–54. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and tissue distribution of fucoidan from Fucus vesiculosus after oral administration to rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, T.; Yokose, T.; Yamamoto, Y.; Yamaguchi, K.; Oda, T. Detection and pharmacokinetics of alginate oligosaccharides in mouse plasma and urine after oral administration by a liquid chromatography/tandem mass spectrometry (LC-MS/MS) method. Biosci. Biotechnol. Biochem. 2008, 72, 2184–2190. [Google Scholar] [CrossRef] [Green Version]
- Ventura, S.; Rodrigues, M.; Falcão, A.; Alves, G. Safety evidence on the administration of Fucus vesiculosus L. (bladderwrack) extract and lamotrigine: Data from pharmacokinetic studies in the rat. Drug Chem. Toxicol. 2020, 43, 560–566. [Google Scholar] [CrossRef]
- Zhang, E.; Chu, F.; Zhao, T.; Chai, Y.; Liang, H.; Song, S.; Ji, A. Determination of fucoidan in rat plasma by HPLC and its application in pharmacokinetics. Pak. J. Pharm. Sci. 2020, 33. [Google Scholar] [CrossRef]
- Lu, J.; Pan, Q.; Zhou, J.; Weng, Y.; Chen, K.; Shi, L.; Zhu, G.; Chen, C.; Li, L.; Geng, M.; et al. Pharmacokinetics, distribution, and excretion of sodium oligomannate, a recently approved anti-Alzheimer’s disease drug in China. J. Pharm. Anal. 2021. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Obluchinskaya, E.D.; Vuorela, H. The pharmacokinetics of fucoidan after topical application to rats. Mar. Drugs 2019, 17, 687. [Google Scholar] [CrossRef] [Green Version]
- Corino, C.; Di Giancamillo, A.; Modina, S.C.; Rossi, R. Prebiotic effects of seaweed polysaccharides in pigs. Animals 2021, 11, 1573. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of marine-derived drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; classification, chemical properties, and future perspective applications in fields of pharmacology and biological medicine (A review of current applications and upcoming potentialities). J. Polym. Environ. 2021. [Google Scholar] [CrossRef]
- Rasmussen, R.S.; Morrissey, M.T. Marine Biotechnology for Production of Food Ingredients. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2007; Volume 52, pp. 237–292. [Google Scholar] [CrossRef]
- Peñalver, R.; Lorenzo, J.M.; Ros, G.; Amarowicz, R.; Pateiro, M.; Nieto, G. Seaweeds as a functional ingredient for a healthy diet. Mar. Drugs 2020, 18, 301. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Cheung, P.C.K. Nutritional evaluation of some subtropical red and green seaweeds: Part I—Proximate composition, amino acid profiles and some physico-chemical properties. Food Chem. 2000, 71, 475–482. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Kraan, S. Chapter 22-Algal polysaccharides, novel applications and outlook. In Carbohydrates-Comprehensive Studies on Glycobiology and Glycotechnology; Chang, C.F., Ed.; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef] [Green Version]
- de Jesus Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Sanz-Pintos, N.; Pérez-Jiménez, J.; Buschmann, A.H.; Vergara-Salinas, J.R.; Pérez-Correa, J.R.; Saura-Calixto, F. Macromolecular antioxidants and dietary fiber in edible seaweeds. J. Food Sci. 2017, 82, 289–295. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Kim, K.-N.; Kim, D.; Jeon, Y.-J. Algal polysaccharides: Potential bioactive substances for cosmeceutical applications. Crit. Rev. Biotechnol. 2019, 39, 99–113. [Google Scholar] [CrossRef]
- Olsson, J.; Toth, G.B.; Albers, E. Biochemical composition of red, green and brown seaweeds on the Swedish west coast. J. Appl. Phycol. 2020. [Google Scholar] [CrossRef]
- Tannock, G.W.; Liu, Y. Guided dietary fibre intake as a means of directing short-chain fatty acid production by the gut microbiota. J. R. Soc. N. Z. 2020, 50, 434–455. [Google Scholar] [CrossRef]
- Hjorth, M.F.; Astrup, A. The role of viscous fiber for weight loss: Food for thought and gut bacteria. Am. J. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Shang, Q.; Li, G.; Wang, X.; Yu, G. Degradation of marine algae-derived carbohydrates by Bacteroidetes isolated from human gut microbiota. Mar. Drugs 2017, 15, 92. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Hemsworth, G.R.; Déjean, G.; Rogers, T.E.; Pudlo, N.A.; Urs, K.; Jain, N.; Davies, G.J.; Martens, E.C.; Brumer, H. Molecular mechanism by which prominent human gut bacteroidetes utilize mixed-linkage beta-glucans, major health-promoting cereal polysaccharides. Cell Rep. 2017, 21, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salyers, A.A.; Vercellotti, J.R.; West, S.E.; Wilkins, T.D. Fermentation of mucin and plant polysaccharides by strains of Bacteroides from the human colon. Appl. Environ. Microbiol. 1977, 33, 319–322. Available online: https://aem.asm.org/content/33/2/319.short (accessed on 11 August 2020). [CrossRef] [PubMed] [Green Version]
- Becker, S.; Tebben, J.; Coffinet, S.; Wiltshire, K.; Iversen, M.H.; Harder, T.; Hinrichs, K.-U.; Hehemann, J.-H. Laminarin is a major molecule in the marine carbon cycle. Proc. Natl. Acad. Sci. USA 2020, 117, 6599–6607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaharudin, N.; Tullin, M.; Pekmez, C.T.; Sloth, J.J.; Rasmussen, R.R.; Dragsted, L.O. Effects of brown seaweeds on postprandial glucose, insulin and appetite in humans–A randomized, 3-way, blinded, cross-over meal study. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Tamez-Hidalgo, P.; Cieplak, T.; Satessa, G.D.; Kot, W.; Søren, S.K.; Nielsen, M.O.; Nielsen, D.S.; Krych, L. Supplementation of a lacto-fermented rapeseed-seaweed blend promotes gut microbial- and gut immune-modulation in weaner piglets. bioRxiv 2020, 2020.09.22.308106. [Google Scholar] [CrossRef]
- Chen, L.; Xu, W.; Chen, D.; Chen, G.; Liu, J.; Zeng, X.; Shao, R.; Zhu, H. Digestibility of sulfated polysaccharide from the brown seaweed Ascophyllum nodosum and its effect on the human gut microbiota in vitro. Int. J. Biol. Macromol. 2018, 112, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Gong, Y.; Li, L.; Hu, X.; Brennan, C.; Kulikouskaya, V. Beneficial effects of three brown seaweed polysaccharides on gut microbiota and their structural characteristics: An overview. Int. J. Food Sci. Tech. 2020, 55, 1199–1206. [Google Scholar] [CrossRef]
- de Borba Gurpilhares, D.; Cinelli, L.P.; Simas, N.K.; Pessoa, A., Jr.; Sette, L.D. Marine prebiotics: Polysaccharides and oligosaccharides obtained by using microbial enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usov, A.; Zelinsky, N. Chapter 2-Chemical structures of algal polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 23–86. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Nutraceutical potential of seaweed polysaccharides: Structure, bioactivity, safety, and toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structure characterization and antioxidant activity of fucoidan isolated from Undaria pinnatifida grown in New Zealand. Carbohydr. Polym. 2019, 212, 178–185. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Skriptsova, A.V. Fucoidans of brown algae: Biosynthesis, localization, and physiological role in thallus. Russ. J. Mar. Biol. 2015, 41, 145–156. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Surits, V.V.; Shevchenko, N.M.; Thinh, P.D.; Zadorozhny, P.A.; Ermakova, S.P. Comparison of structure and in vitro anticancer activity of native and modified fucoidans from Sargassum feldmannii and S. duplicatum. Int. J. Biol. Macromol. 2019, 124, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Ngoc Hieu, B.T.; Lin, Y.C. Low-molecular-weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food Nutr. Res. 2016, 60. [Google Scholar] [CrossRef] [Green Version]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M. Pilot clinical study to evaluate the anticoagulant activity of fucoidan. Blood Coagul. Fibrinolysis 2009, 20, 607–610. [Google Scholar] [CrossRef]
- Tsai, H.-L.; Tai, C.-J.; Huang, C.-W.; Chang, F.-R.; Wang, J.-Y. Efficacy of low-molecular-weight fucoidan as a supplemental therapy in metastatic colorectal cancer patients: A double-blind randomized controlled trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, H.; Tamauchi, H.; Hashimoto, M.; Nakano, T. Antitumor activity and immune response of Mekabu fucoidan extracted from sporophyll of Undaria pinnatifida. In Vivo 2003, 17, 245–249. Available online: http://europepmc.org/abstract/MED/12929574 (accessed on 19 May 2020).
- Lin, H.-T.V.; Tsou, Y.-C.; Chen, Y.-T.; Lu, W.-J.; Hwang, P.-A. Effects of low-molecular-weight fucoidan and high stability fucoxanthin on glucose homeostasis, lipid metabolism, and liver function in a mouse model of type II diabetes. Mar. Drugs 2017, 15, 113. [Google Scholar] [CrossRef] [Green Version]
- Okolie, C.L.; Mason, B.; Mohan, A.; Pitts, N.; Udenigwe, C.C. The comparative influence of novel extraction technologies on in vitro prebiotic-inducing chemical properties of fucoidan extracts from Ascophyllum nodosum. Food Hydrocoll. 2019, 90, 462–471. [Google Scholar] [CrossRef]
- Kan, J.; Cheng, J.; Xu, L.; Hood, M.; Zhong, D.; Cheng, M.; Liu, Y.; Chen, L.; Du, J. The combination of wheat peptides and fucoidan protects against chronic superficial gastritis and alters gut microbiota: A double-blinded, placebo-controlled study. Eur. J. Nutr. 2020, 59, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Takahashi, K.; Abe, S.; Yamada, K.; Suzuki, M.; Masahisa, M.; Endo, M.; Abe, K.; Inoue, R.; Hoshi, H. Improvement of psoriasis by alteration of the gut environment by oral administration of fucoidan from Cladosiphon okamuranus. Mar. Drugs 2020, 18, 154. [Google Scholar] [CrossRef] [Green Version]
- Parnell, J.A.; Reimer, R.A. Prebiotic fiber modulation of the gut microbiota improves risk factors for obesity and the metabolic syndrome. Gut Microbes 2012, 3, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, Y.; Chen, X.-Q.; Aweya, J.J.; Cheong, K.-L. Catabolism of Saccharina japonica polysaccharides and oligosaccharides by human fecal microbiota. LWT 2020, 130, 109635. [Google Scholar] [CrossRef]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Olatunji, O. Aquatic Biopolymers: Understanding Their Industrial Significance and Environmental Implications; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Baweja, P.; Sahoo, D. Chapter 2-Classification of algae. In The Algae World; Sahoo, D., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 31–55. [Google Scholar] [CrossRef] [Green Version]
- Usman, A.; Khalid, S.; Usman, A.; Hussain, Z.; Wang, Y. Chapter 5-Algal polysaccharides, novel application, and outlook. In Algae Based Polymers, Blends, and Composites; Zia, K.M., Zuber, M., Ali, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 115–153. [Google Scholar] [CrossRef]
- Kadam, S.U.; O’Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound assisted extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Xu, L.; Zhang, W.; Cai, Y.; Jang, B.; Oh, J.; Jin, J.O. Laminarin promotes anti-cancer immunity by the maturation of dendritic cells. Oncotarget 2017, 8, 38554–38567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, H.-Q.; Elkin, M.; Aingorn, E.; Ishai-Michaeli, R.; Stein, C.A.; Vlodavsky, I. Inhibition of heparanase activity and tumor metastasis by laminarin sulfate and synthetic phosphorothioate oligodeoxynucleotides. Int. J. Cancer 1999, 83, 424–431. [Google Scholar] [CrossRef]
- Cuong, D.X. Laminarin (beta-glucan) of brown algae Sargassum mcclurei: Extraction, antioxidant activity, lipoxygenase inhibition activity, and physicochemistry properties. World J. Food Sci. Technol. 2020, 4, 31. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y.-J.; Kim, H.; Kim, Y.-S.; Park, W. Immunostimulatory effect of laminarin on RAW 264.7 mouse macrophages. Molecules 2012, 17, 5404–5411. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.; Sweeney, T.; Bahar, B.; O’Doherty, J. Effect of maternal seaweed extract supplementation on suckling piglet growth, humoral immunity, selected microflora, and immune response after an ex vivo lipopolysaccharide challenge. J. Anim. Sci. 2012, 90, 505–514. [Google Scholar] [CrossRef]
- Vigors, S.; O’Doherty, J.V.; Rattigan, R.; McDonnell, M.J.; Rajauria, G.; Sweeney, T. Effect of a laminarin rich macroalgal extract on the caecal and colonic microbiota in the post-weaned pig. Mar. Drugs 2020, 18, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattigan, R.; Sweeney, T.; Maher, S.; Thornton, K.; Rajauria, G.; O’Doherty, J.V. Laminarin-rich extract improves growth performance, small intestinal morphology, gene expression of nutrient transporters and the large intestinal microbial composition of piglets during the critical post-weaning period. Br. J. Nutr. 2019, 123, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.B.; Sweeney, T.; Callan, J.J.; O’Sullivan, J.T.; O’Doherty, J.V. The effect of dietary Laminaria-derived laminarin and fucoidan on nutrient digestibility, nitrogen utilisation, intestinal microflora and volatile fatty acid concentration in pigs. J. Sci. Food Agric. 2010, 90, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, N.; Yang, L.; Hong, Z.; Cai, B.; Le, Q.; Yang, T.; Shi, L.; He, J.; Cui, C.-B. Insoluble dietary fiber derived from brown seaweed Laminaria japonica ameliorate obesity-related features via modulating gut microbiota dysbiosis in high-fat diet-fed mice. Food Funct. 2020. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Cotas, J. Chapter 1-Introductory chapter: Alginates-a general overview. In Alginates-Recent Uses of This Natural Polymer; Pereira, L., Ed.; IntechOpen: Rijeka, Croatia, 2020. [Google Scholar] [CrossRef] [Green Version]
- Mei, X.; Chang, Y.; Shen, J.; Zhang, Y.; Xue, C. Expression and characterization of a novel alginate-binding protein: A promising tool for investigating alginate. Carbohydr. Polym. 2020, 246, 116645. [Google Scholar] [CrossRef]
- Ramos, P.E.; Silva, P.; Alario, M.M.; Pastrana, L.M.; Teixeira, J.A.; Cerqueira, M.A.; Vicente, A.A. Effect of alginate molecular weight and M/G ratio in beads properties foreseeing the protection of probiotics. Food Hydrocoll. 2018, 77, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Mancini, F.; Montanari, L.; Peressini, D.; Fantozzi, P. Influence of alginate concentration and molecular weight on functional properties of mayonnaise. LWT 2002, 35, 517–525. [Google Scholar] [CrossRef]
- Jönsson, M.; Allahgholi, L.; Sardari, R.R.; Hreggviðsson, G.O.; Nordberg Karlsson, E. Extraction and modification of macroalgal polysaccharides for current and next-generation applications. Molecules 2020, 25, 930. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Chen, H.; Zhu, L.; Liu, W.; Yu, H.D.; Wang, X.; Yin, Y. Comparative study on the in vitro effects of Pseudomonas aeruginosa and seaweed alginates on human gut microbiota. PLoS ONE 2017, 12, e0171576. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, G.; Shang, Q.; Chen, X.; Liu, W.; Pi, X.; Zhu, L.; Yin, Y.; Yu, G.; Wang, X. In vitro fermentation of alginate and its derivatives by human gut microbiota. Anaerobe 2016, 39, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Bamba, S.; Abe, N.; Sasaki, M. Effects of an alginate-containing variable-viscosity enteral nutrition formula on defecation, intestinal microbiota, and short-chain fatty acid production. J. Funct. Foods 2020, 67, 103852. [Google Scholar] [CrossRef]
- Georg-Jensen, M.; Pedersen, C.; Kristensen, M.; Frost, G.; Astrup, A. Efficacy of alginate supplementation in relation to appetite regulation and metabolic risk factors: Evidence from animal and human studies. Obes. Rev. 2013, 14, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Goff, H.D.; Xu, F.; Liu, F.; Ma, J.; Chen, M.; Zhong, F. The effect of sodium alginate on nutrient digestion and metabolic responses during both in vitro and in vivo digestion process. Food Hydrocoll. 2020, 107, 105304. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, Z.; Feng, W.; Hu, T.; Guan, H.; Mao, Y. AOS ameliorates monocrotaline-induced pulmonary hypertension by restraining the activation of P-selectin/p38MAPK/NF-κB pathway in rats. Biomed. Pharmacother. 2019, 109, 1319–1326. [Google Scholar] [CrossRef]
- Amimi, A.; Mouradi, A.; Bennasser, L.; Givernaud, T. Seasonal variations in thalli and carrageenan composition of Gigartina pistillata (Gmelin) Stackhouse (Rhodophyta, Gigartinales) harvested along the Atlantic coast of Morocco. Phycol. Res. 2007, 55, 143–149. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Oladzadabbasabadi, N.; Ebadi, S.; Nafchi, A.M.; Karim, A.; Kiahosseini, S.R. Functional properties of dually modified sago starch/κ-carrageenan films: An alternative to gelatin in pharmaceutical capsules. Carbohydr. Polym. 2017, 160, 43–51. [Google Scholar] [CrossRef]
- McKim, J.; Willoughby, J.; Blakemore, W.; Weiner, M. Clarifying the confusion between poligeenan, degraded carrageenan, and carrageenan: A review of the chemistry, nomenclature, and in vivo toxicology by the oral route. Crit. Rev. Food Sci. Nutr. 2018, 59, 1–70. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Omoto, T.; Goto, Y.; Asai, I.; Nakamura, M.; Maitani, T. Molecular weight distribution of carrageenans studied by a combined gel permeation/inductively coupled plasma (GPC/ICP) method. Food Addit. Contam. 2001, 18, 763–772. [Google Scholar] [CrossRef]
- Du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the red seaweed Sarconema filiforme attenuate symptoms of diet-induced metabolic syndrome in rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Cui, X.; Duan, M.; Ai, C.; Song, S.; Chen, X. In vitro fermentation of κ-carrageenan oligosaccharides by human gut microbiota and its inflammatory effect on HT29 cells. J. Func. Foods 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef]
- Xu, S.-Y.; Aweya, J.J.; Li, N.; Deng, R.-Y.; Chen, W.-Y.; Tang, J.; Cheong, K.-L. Microbial catabolism of Porphyra haitanensis polysaccharides by human gut microbiota. Food Chem. 2019, 289, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.-M.; Veeraperumal, S.; Lv, J.-H.; Wu, T.-C.; Zhang, Z.-P.; Zeng, Q.-K.; Liu, Y.; Chen, X.-Q.; Aweya, J.J.; Cheong, K.-L. Physicochemical properties and potential beneficial effects of porphyran from Porphyra haitanensis on intestinal epithelial cells. Carbohydr. Polym. 2020, 246, 116626. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, Q.; Qi, H.; Zhang, H.; Niu, X.; Xu, Z.; Li, Z. Degradation of porphyran from Porphyra haitanensis and the antioxidant activities of the degraded porphyrans with different molecular weight. Int. J. Biol. Macromol. 2006, 38, 45–50. [Google Scholar] [CrossRef]
- Bhatia, S.; Sharma, A.; Sharma, K.; Kavale, M.; Chaugule, B.; Dhalwal, K.; Namdeo, A.; Mahadik, K. Novel algal polysaccharides from marine source: Porphyran. Pharmacogn. Rev. 2008, 2, 271. Available online: https://www.phcogrev.com/sites/default/files/PhcogRev-2-4-271.pdf (accessed on 8 June 2020).
- He, D.; Wu, S.; Yan, L.; Zuo, J.; Cheng, Y.; Wang, H.; Liu, J.; Zhang, X.; Wu, M.; Choi, J.-I.; et al. Antitumor bioactivity of porphyran extracted from Pyropia yezoensis Chonsoo2 on human cancer cell lines. J. Sci. Food Agric. 2019, 99, 6722–6730. [Google Scholar] [CrossRef]
- Kwon, M.-J.; Nam, T.-J. Chromatographically purified porphyran from Porphyra yezoensis effectively inhibits proliferation of human cancer cells. Food Sci. Biotechnol. 2007, 16, 873–878. Available online: https://www.koreascience.or.kr/article/JAKO200709905796598.page (accessed on 13 July 2020).
- Seong, H.; Bae, J.-H.; Seo, J.S.; Kim, S.-A.; Kim, T.-J.; Han, N.S. Comparative analysis of prebiotic effects of seaweed polysaccharides laminaran, porphyran, and ulvan using in vitro human fecal fermentation. J. Funct. Foods 2019, 57, 408–416. [Google Scholar] [CrossRef]
- Xu, S.-Y.; Chen, X.-Q.; Liu, Y.; Cheong, K.-L. Ultrasonic/microwave-assisted extraction, simulated digestion, and fermentation in vitro by human intestinal flora of polysaccharides from Porphyra haitanensis. Int. J. Biol. Macromol. 2020, 152, 748–756. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Rathgeber, B.; Stratton, G.; Thomas, N.; Evans, F.; Critchley, A.; Hafting, J.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poult. Sci. 2014, 93, 2991–3001. [Google Scholar] [CrossRef]
- Liu, J.; Kandasamy, S.; Zhang, J.; Kirby, C.W.; Karakach, T.; Hafting, J.; Critchley, A.T.; Evans, F.; Prithiviraj, B. Prebiotic effects of diet supplemented with the cultivated red seaweed Chondrus crispus or with fructo-oligo-saccharide on host immunity, colonic microbiota and gut microbial metabolites. BMC Complement. Altern. Med. 2015, 15, 279. [Google Scholar] [CrossRef]
- Balasubramanian, B.; Shanmugam, S.; Park, S.; Recharla, N.; Koo, J.S.; Andretta, I.; Kim, I.H. Supplemental impact of marine red seaweed (Halymenia palmata) on the growth performance, total tract nutrient digestibility, blood profiles, intestine histomorphology, meat quality, fecal gas emission, and microbial counts in broilers. Animals 2021, 11, 1244. [Google Scholar] [CrossRef]
- Lahaye, M. NMR spectroscopic characterisation of oligosaccharides from two Ulva rigida ulvan samples (Ulvales, Chlorophyta) degraded by a lyase. Carbohydr. Res. 1998, 314, 1–12. [Google Scholar] [CrossRef]
- Michel, G.; Czjzek, M. Chapter 16-Polysaccharide-degrading enzymes from marine bacteria. In Marine Enzymes for Biocatalysis: Sources, Biocatalytic Characteristics and Bioprocesses of Marine Enzymes; Trincone, A., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 429–464. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Adrien, A.; Bonnet, A.; Dufour, D.; Baudouin, S.; Maugard, T.; Bridiau, N. Anticoagulant activity of sulfated ulvan isolated from the green macroalga Ulva rigida. Mar. Drugs 2019, 17, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klongklaew, N.; Praiboon, J.; Tamtin, M.; Srisapoome, P. Antibacterial and antiviral activities of local Thai green macroalgae crude extracts in pacific white shrimp (Litopenaeus vannamei). Mar. Drugs 2020, 18, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, Y.; Zhang, M.; Wang, X.; Fu, X.; Guan, H.; Wang, P. Ulvan lyase assisted structural characterization of ulvan from Ulva pertusa and its antiviral activity against vesicular stomatitis virus. Int. J. Biol. Macromol. 2020, 157, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Cañedo-Castro, B.; Piñón-Gimate, A.; Carrillo, S.; Ramos, D.; Casas-Valdez, M. Prebiotic effect of Ulva rigida meal on the intestinal integrity and serum cholesterol and triglyceride content in broilers. J. Appl. Phycol. 2019, 31, 3265–3273. [Google Scholar] [CrossRef]
- Shalaby, M.; Amin, H. Potential using of ulvan polysaccharides from Ulva lactuca as a prebiotic in symbiotic yogurt production. J. Probiot. Health 2019, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Dong, S.; Gao, J.; Jiang, C. In vitro fermentation of sulfated polysaccharides from E. prolifera and L. japonica by human fecal microbiota. Int. J. Biol. Macromol. 2016, 91, 867–871. [Google Scholar] [CrossRef]
- Strain, C.R.; Collins, K.C.; Naughton, V.; McSorley, E.M.; Stanton, C.; Smyth, T.J.; Soler-Vila, A.; Rea, M.C.; Ross, P.R.; Cherry, P.; et al. Effects of a polysaccharide-rich extract derived from Irish-sourced Laminaria digitata on the composition and metabolic activity of the human gut microbiota using an in vitro colonic model. Eur. J. Nutr. 2020, 59, 309–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensiddhi, S.; Conlon, M.A.; Vuaran, M.S.; Franco, C.M.M.; Zhang, W. Impact of extraction processes on prebiotic potential of the brown seaweed Ecklonia radiata by in vitro human gut bacteria fermentation. J. Funct. Foods 2016, 24, 221–230. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Conlon, M.A.; Methacanon, P.; Franco, C.M.M.; Su, P.; Zhang, W. Gut health benefits of brown seaweed Ecklonia radiata and its polysaccharides demonstrated in vivo in a rat model. J. Funct. Foods 2017, 37, 676–684. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, G.; Peng, Y.; Rui, Y.; Zeng, X.; Ye, H. Simulated digestion and fermentation in vitro with human gut microbiota of polysaccharides from Coralline pilulifera. LWT 2019, 100, 167–174. [Google Scholar] [CrossRef]
- Cui, M.; Zhou, R.; Wang, Y.; Zhang, M.; Liu, K.; Ma, C. Beneficial effects of sulfated polysaccharides from the red seaweed Gelidium pacificum Okamura on mice with antibiotic-associated diarrhea. Food Funct. 2020, 11, 4625–4637. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Warren, F.J.; Gidley, M.J. Natural products for glycaemic control: Polyphenols as inhibitors of alpha-amylase. Trends Food Sci. Technol. 2019, 91, 262–273. [Google Scholar] [CrossRef]
- Mannino, A.M.; Micheli, C. Ecological function of phenolic compounds from Mediterranean fucoid algae and seagrasses: An overview on the genus Cystoseira sensu lato and Posidonia oceanica (L.) Delile. J. Mar. Sci. Eng. 2020, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–598. [Google Scholar] [CrossRef]
- Poole, J.; Diop, A.; Rainville, L.C.; Barnabé, S. Bioextracting polyphenols from the brown seaweed Ascophyllum nodosum from Québec’s north shore coastline. Ind. Biotechnol. 2019, 15, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Wekre, M.E.; Kåsin, K.; Underhaug, J.; Holmelid, B.; Jordheim, M. Quantification of polyphenols in seaweeds: A case study of Ulva intestinalis. Antioxidants 2019, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Freile-Pelegrin, Y.; Robledo, D. Chapter 6-Bioactive phenolic compounds from algae. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; Hernández-Ledesma, B., Herrero, M., Eds.; Wiley-Blackwell: Chichester, UK, 2014; pp. 113–129. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Cox, K.; Scholey, A.; Ryan, L.; Bonham, M.P. Twelve weeks’ treatment with a polyphenol-rich seaweed extract increased HDL cholesterol with no change in other biomarkers of chronic disease risk in overweight adults: A placebo-controlled randomised trial. J. Nutr. Biochem. 2021, 108777. [Google Scholar] [CrossRef] [PubMed]
- Haskell-Ramsay, C.F.; Jackson, P.A.; Dodd, F.L.; Forster, J.S.; Bérubé, J.; Levinton, C.; Kennedy, D.O. Acute post-prandial cognitive effects of brown seaweed extract in humans. Nutrients 2018, 10, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hata, Y.; Nakajima, K.; Uchida, J.-I.; Hidaka, H.; Nakano, T. Clinical effects of brown seaweed, Undaria pinnatifida (wakame), on blood pressure in hypertensive subjects. J. Clin. Biochem. Nutr. 2001, 30, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Derosa, G.; Pascuzzo, M.D.; D’Angelo, A.; Maffioli, P. Ascophyllum nodosum, Fucus vesiculosus and chromium picolinate nutraceutical composition can help to treat type 2 diabetic patients. Diabetes. Metab. Syndr. Obes. 2019, 12, 1861–1865. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. A single-dose of a polyphenol-rich Fucus vesiculosus extract is insufficient to blunt the elevated postprandial blood glucose responses exhibited by healthy adults in the evening: A randomised crossover trial. Antioxidants 2019, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Kumar Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar Rana, H.; Gupta, A.; Rosaria Lauro, M.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.F.; Maier, C.S. The chemistry of gut microbial metabolism of polyphenols. Phytochem. Rev. 2016, 15, 425–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food. Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Das, G.; Das, S.K. Roles of flavonoids in plants. Int. J. Pharm. Sci. Tech. 2011, 6, 12–35. Available online: https://www.researchgate.net/profile/Sanjoy_Das10/publication/279499208_Roles_of_flavonoids_in_Plants/links/5667098108ae8905db8b8b47.pdf (accessed on 27 July 2020).
- Lin, G.; Liu, X.; Yan, X.; Liu, D.; Yang, C.; Liu, B.; Huang, Y.; Zhao, C. Role of green macroalgae Enteromorpha prolifera polyphenols in the modulation of gene expression and intestinal microflora profiles in type 2 diabetic mice. Int. J. Mol. Sci. 2019, 20, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshie-Stark, Y.; Hsieh, Y.-P.; Suzuki, T. Distribution of flavonoids and related compounds from seaweeds in Japan. J. Tokyo Univ. Fish. 2003, 89, 1–6. Available online: https://www.researchgate.net/publication/285022048_Distribution_of_flavonoids_and_related_compounds_from_seaweeds_in_Japan (accessed on 3 July 2020).
- Culioli, G.; Ortalo-Magné, A.; Valls, R.; Hellio, C.; Clare, A.S.; Piovetti, L. Antifouling activity of meroditerpenoids from the marine brown alga Halidrys siliquosa. J. Nat. Prod. 2008, 71, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential role of seaweed polyphenols in cardiovascular-associated disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Dong, S.; Erik Hansen, P.; Stagos, D.; Lin, X.; Liu, M. Progress of bromophenols in marine algae from 2011 to 2020: Structure, bioactivities, and applications. Mar. Drugs 2020, 18, 411. [Google Scholar] [CrossRef]
- Shibata, T.; Miyasaki, T.; Miyake, H.; Tanaka, R.; Kawaguchi, S. The influence of phlorotannins and bromophenols on the feeding behavior of marine herbivorous gastropod Turbo cornutus. Am. J. Plant Sci. 2014, 5, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, B.V.; Maneein, S.; Farid, A.; Mahmud, M.; Milledge, J.J. The effects of halogenated compounds on the anaerobic digestion of macroalgae. Fermentation 2020, 6, 85. [Google Scholar] [CrossRef]
- Hay, M.E.; Fenical, W. Marine plant-herbivore interactions: The ecology of chemical defense. Annu. Rev. Ecol. Syst. 1988, 19, 111–145. [Google Scholar] [CrossRef]
- Whitfield, F.; Helidoniotis, F.; Drew, M. Effect of Diet and Environment on the Volatile Flavour Components of Crustaceans; CSIRO and Fisheries Research & Development Corporation: North Ryde, NSW, Australia, 1995. Available online: http://www.frdc.com.au/Archived-Reports/FRDC%20Projects/1992-075-DLD.pdf (accessed on 8 June 2020).
- Whitfield, F.B.; Helidoniotis, F.; Shaw, K.J.; Svoronos, D. Distribution of bromophenols in species of marine algae from eastern Australia. J. Agric. Food. Chem. 1999, 47, 2367–2373. [Google Scholar] [CrossRef]
- Luo, J.; Xu, Q.; Jiang, B.; Zhang, R.; Jia, X.; Li, X.; Wang, L.; Guo, C.; Wu, N.; Shi, D. Selectivity, cell permeability and oral availability studies of novel bromophenol derivative HPN as protein tyrosine phosphatase 1B inhibitor. Br. J. Pharmacol. 2018, 175, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Glukhov, E.; Yu, H.; Gerwick, L.; Dorrestein, P.; Gerwick, W. Monomeric and dimeric bromophenols from the red alga Ceramium sp. with antioxidant and anti-inflammatory activities. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Cherian, C.; Vennila, J.J.; Sharan, L. Marine bromophenols as an effective inhibitor of virulent proteins (peptidyl arginine deiminase, gingipain R and hemagglutinin A) in Porphyromas gingivalis. Arch. Oral Biol. 2019, 100, 119–128. [Google Scholar] [CrossRef]
- Shi, D.; Li, J.; Guo, S.; Su, H.; Fan, X. The antitumor effect of bromophenol derivatives in vitro and Leathesia nana extract in vivo. Chin. J. Oceanol. Limnol. 2009, 27, 277–282. [Google Scholar] [CrossRef]
- Shi, D.; Li, X.; Li, J.; Guo, S.; Su, H.; Fan, X. Antithrombotic effects of bromophenol, an alga-derived thrombin inhibitor. Chin. J. Oceanol. Limnol. 2010, 28, 96–98. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Nguyen, T.L.P.; Tran, T.V.A.; Do, A.D.; Kim, S.M. Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 2019, 9, 501. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, D.; Sun, Y.; Gu, Y.; Ming, Y.; Zheng, J.; Yu, C.; Chen, X.; Qi, H. Synergistic effects of UVA irradiation and phlorotannin extracts of Laminaria japonica on properties of grass carp myofibrillar protein gel. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef]
- Lemesheva, V.; Birkemeyer, C.; Garbary, D.; Tarakhovskaya, E. Vanadium-dependent haloperoxidase activity and phlorotannin incorporation into the cell wall during early embryogenesis of Fucus vesiculosus (Phaeophyceae). Eur. J. Phycol. 2020, 55, 275–284. [Google Scholar] [CrossRef]
- Gómez, I.; Huovinen, P. Induction of phlorotannins during UV exposure mitigates inhibition of photosynthesis and DNA damage in the kelp Lessonia nigrescens. Photochem. Photobiol. 2010, 86, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Arnold, T.M.; Targett, N.M. To grow and defend: Lack of tradeoffs for brown algal phlorotannins. Oikos 2003, 100, 406–408. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Vallejo, F.; Gil-Izquierdo, Á.; Andrade, P.B.; Valentão, P.; Pereira, D.M.; Ferreres, F. Profiling phlorotannins from Fucus spp. of the Northern Portuguese coastline: Chemical approach by HPLC-DAD-ESI/MSn and UPLC-ESI-QTOF/MS. Algal Res. 2018, 29, 113–120. [Google Scholar] [CrossRef]
- Sonani, R.; Rastogi, R.; Madamwar, D. Chapter 5-Natural Antioxidants From Algae: A Therapeutic Perspective. In Algal Green Chemistry: Recent Progress in Biotechnology; Rastogi, R.P., Madamwar, D., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 91–120. [Google Scholar] [CrossRef]
- Heffernan, N.; Brunton, N.P.; FitzGerald, R.J.; Smyth, T.J. Profiling of the molecular weight and structural isomer abundance of macroalgae-derived phlorotannins. Mar. Drugs 2015, 13, 509–528. [Google Scholar] [CrossRef]
- Steevensz, A.J.; MacKinnon, S.L.; Hankinson, R.; Craft, C.; Connan, S.; Stengel, D.B.; Melanson, J.E. Profiling phlorotannins in brown macroalgae by liquid chromatography–high resolution mass spectrometry. Phytochem. Anal. 2012, 23, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Barwell, C.J.; Blunden, G.; Manandhar, P.D. Isolation and characterization of brown algal polyphenols as inhibitors of α-amylase, lipase and trypsin. J. Appl. Phycol. 1989, 1, 319–323. [Google Scholar] [CrossRef]
- Geiselman, J.A.; McConnell, O.J. Polyphenols in brown algae Fucus vesiculosus and Ascophyllum nodosum: Chemical defenses against the marine herbivorous snail, Littorina littorea. J. Chem. Ecol. 1981, 7, 1115–1133. [Google Scholar] [CrossRef]
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Gall, E.A. Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.W.; Jeon, J.-S.; Jung, Y.-J.; Kim, W.-R.; Kim, C.Y.; Um, B.-H. Determination of major phlorotannins in Eisenia bicyclis using hydrophilic interaction chromatography: Seasonal variation and extraction characteristics. Food Chem. 2013, 138, 2399–2406. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Yong-Li; Karadeniz, F.; Kim, M.-M.; Kim, S.-K. α-Glucosidase and α-amylase inhibitory activities of phloroglucinal derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food Agric. 2009, 89, 1552–1558. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Lajili, S.; Elkaibi, M.; Muller, C.; Majdoub, H.; Jamil, K.; Bouraoui, A. Optimized extraction, preliminary characterization and evaluation of the in vitro anticancer activity of phlorotannin-rich fraction from the brown seaweed, Cystoseira sedoides. J. Aquat. Food Prod. Technol. 2019, 28, 892–909. [Google Scholar] [CrossRef]
- Kim, H.-J.; Yong, H.I.; Lee, B.W.; Park, S.; Baek, K.H.; Kim, T.H.; Jo, C. Plasma-polymerized phlorotannins and their enhanced biological activities. J. Agric. Food. Chem. 2020, 68, 2357–2365. [Google Scholar] [CrossRef]
- Dong, X.; Bai, Y.; Xu, Z.; Shi, Y.; Sun, Y.; Janaswamy, S.; Yu, C.; Qi, H. Phlorotannins from Undaria pinnatifida sporophyll: Extraction, antioxidant, and anti-inflammatory activities. Mar. Drugs 2019, 17, 434. [Google Scholar] [CrossRef] [Green Version]
- Artan, M.; Li, Y.; Karadeniz, F.; Lee, S.-H.; Kim, M.-M.; Kim, S.-K. Anti-HIV-1 activity of phloroglucinol derivative, 6,6′-bieckol, from Ecklonia cava. Biorg. Med. Chem. 2008, 16, 7921–7926. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yi, M.; Ding, L.; He, S.; Yan, X. Isolation and Purification of a neuroprotective phlorotannin from the marine algae Ecklonia maxima by size exclusion and high-speed counter-current chromatography. Mar. Drugs 2019, 17, 212. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Wang, W.; Chu, W. Antimicrobial and anti-quorum sensing activities of phlorotannins from seaweed (Hizikia fusiforme). Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Ji, Y.; Anegboonlap, P.; Hotchkiss, S.; Gill, C.; Yaqoob, P.; Spencer, J.P.; Rowland, I. Gastrointestinal modifications and bioavailability of brown seaweed phlorotannins and effects on inflammatory markers. Br. J. Nutr. 2016, 115, 1240–1253. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Z.; Bach, S.J.; McAllister, T.A. Effects of phlorotannins from Ascophyllum nodosum (brown seaweed) on in vitro ruminal digestion of mixed forage or barley grain. Anim. Feed Sci. Technol. 2008, 145, 375–395. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C.; Chen, M.; Lv, X.; Liu, B.; Yi, L.; Cornara, L.; Wei, M.-C.; Yang, Y.-C.; Tundis, R.; et al. Regulatory efficacy of brown seaweed Lessonia nigrescens extract on the gene expression profile and intestinal microflora in type 2 diabetic mice. Mol. Nutr. Food Res. 2018, 62, 1700730. [Google Scholar] [CrossRef]
- Hirosumi, J.; Tuncman, G.; Chang, L.; Görgün, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central role for JNK in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-rich extracts from brown macroalgae Lessonia trabeculate attenuate hyperglycemia and modulate gut microbiota in high-fat diet and streptozotocin-induced diabetic rats. J. Agric. Food. Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, T.; Li, Y.; Yuan, C.; Ma, H.; Seeram, N.P.; Liu, F.; Mu, Y.; Huang, X.; Li, L. Hypoglycemic and hypolipidemic effects of triterpenoid-enriched Jamun (Eugenia jambolana Lam.) fruit extract in streptozotocin-induced type 1 diabetic mice. Food Funct. 2018, 9, 3330–3337. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.-B.; Lee, E.; Park, H.-Y. The effects of gelatinized wheat starch and high salt diet on gut microbiota and metabolic disorder. Nutrients 2020, 12, 301. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-R.; Lin, C.-S.; Chang, C.-J.; Lin, T.-L.; Martel, J.; Ko, Y.-F.; Ojcius, D.M.; Lu, C.-C.; Young, J.D.; Lai, H.-C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef]
- Corona, G.; Coman, M.; Guo, Y.; Hotchkiss, S.; Gill, C.; Yaqoob, P.; Spencer, J.; Rowland, I. Effect of simulated gastrointestinal digestion and fermentation on polyphenolic content and bioactivity of brown seaweed phlorotannin-rich extracts. Mol. Nutr. Food Res. 2017, 61, 1700223. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food. Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Purification and identification of dipeptidyl peptidase (DPP) IV inhibitory peptides from the macroalga Palmaria palmata. Food Chem. 2015, 172, 400–406. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Gallagher, E.; O’Connor, P.; Prieto, J.; Mora-Soler, L.; Grealy, M.; Hayes, M. Development of a seaweed derived platelet activating factor acetylhydrolase (PAF-AH) inhibitory hydrolysate, synthesis of inhibitory peptides and assessment of their toxicity using the Zebrafish larvae assay. Peptides 2013, 50, 119–124. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.; Yang, R.; Zhao, W. Identification of bioactive peptides with α-amylase inhibitory potential from enzymatic protein hydrolysates of red seaweed (Porphyra spp). J. Agric. Food. Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Cian, R.E.; Hernández-Chirlaque, C.; Gámez-Belmonte, R.; Drago, S.R.; Sánchez de Medina, F.; Martínez-Augustin, O. Green alga Ulva spp. hydrolysates and their peptide fractions regulate cytokine production in splenic macrophages and lymphocytes involving the TLR4-NFκB/MAPK pathways. Mar. Drugs 2018, 16, 235. [Google Scholar] [CrossRef] [Green Version]
- Minkova, K.M.; Toshkova, R.A.; Gardeva, E.G.; Tchorbadjieva, M.I.; Ivanova, N.J.; Yossifova, L.S.; Gigova, L.G. Antitumor activity of B-phycoerythrin from Porphyridium cruentum. J. Pharm. Res. 2011, 4, 1480–1482. Available online: https://www.researchgate.net/profile/Liliya_Yossifova/publication/228107475_Antitumor_activity_of_B-phycoerythrin_from_Porphyridium_cruentum/links/0fcfd50732fa2025d2000000.pdf (accessed on 16 September 2020).
- Venkatraman, K.L.; Syed, A.A.; Indumathi, P.; Mehta, A. VITPOR AI, a coagulation factor XIIa inhibitor from Porphyra yezoensis: In vivo mode of action and assessment of platelet function analysis. Protein Pept. Lett. 2020, 27, 243–250. [Google Scholar] [CrossRef]
- McLaughlin, C.M.; Sharkey, S.J.; Harnedy-Rothwell, P.; Parthsarathy, V.; Allsopp, P.J.; McSorley, E.M.; FitzGerald, R.J.; O’Harte, F.P.M. Twice daily oral administration of Palmaria palmata protein hydrolysate reduces food intake in streptozotocin induced diabetic mice, improving glycaemic control and lipid profiles. J. Funct. Foods 2020, 73, 104101. [Google Scholar] [CrossRef]
- Dave, L.A.; Hayes, M.; Mora, L.; Rutherfurd, S.M.; Montoya, C.A.; Moughan, P.J. Bioactive peptides originating from gastrointestinal endogenous proteins in the growing pig: In vivo identification. Curr. Pharm. Des. 2021, 27, 1382–1395. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Aluko, R.E.; Hossain, M.; Rai, D.K.; Hayes, M. Potential of a renin inhibitory peptide from the red seaweed Palmaria palmata as a functional food ingredient following confirmation and characterization of a hypotensive effect in spontaneously hypertensive rats. J. Agric. Food. Chem. 2014, 62, 8352–8356. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, P.; Crowe, W.; Bahar, B.; Harnedy, P.A.; Brown, E.S.; Taylor, S.S.; Smyth, T.J.; Soler-Vila, A.; Magee, P.J.; Gill, C.I.R.; et al. The effect of consuming Palmaria palmata-enriched bread on inflammatory markers, antioxidant status, lipid profile and thyroid function in a randomised placebo-controlled intervention trial in healthy adults. Eur. J. Nutr. 2016, 55, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-converting enzyme inhibitory peptides derived from Wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food. Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Fractionation and identification of antioxidant peptides from an enzymatically hydrolysed Palmaria palmata protein isolate. Food Res. Int. 2017, 100, 416–422. [Google Scholar] [CrossRef] [Green Version]
- Amaretti, A.; Gozzoli, C.; Simone, M.; Raimondi, S.; Righini, L.; Pérez-Brocal, V.; García-López, R.; Moya, A.; Rossi, M. Profiling of protein degraders in cultures of human gut microbiota. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial fermentation of dietary protein: An important factor in diet-microbe-host interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of dietary protein and peptides by intestinal microbes and their impacts on gut. Curr. Protein Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hetzel, M.; Boiangiu, C.D.; Buckel, W. Dehydration of (R)-2-hydroxyacyl-CoA to enoyl-CoA in the fermentation of α-amino acids by anaerobic bacteria. FEMS Microbiol. Rev. 2004, 28, 455–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachier, F.; Mariotti, F.; Huneau, J.F.; Tomé, D. Effects of amino acid-derived luminal metabolites on the colonic epithelium and physiopathological consequences. Amino Acids 2007, 33, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Ao, H.; Peng, C. Gut microbiota, short-chain fatty acids, and herbal medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Portune, K.J.; Beaumont, M.; Davila, A.-M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57 Pt B, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review article: Insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharmacol. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpela, K. Diet, microbiota, and metabolic health: Trade-off between saccharolytic and proteolytic fermentation. Annu. Rev. Food Sci. Technol. 2018, 9, 65–84. [Google Scholar] [CrossRef]
- Wang, X.; Gibson, G.R.; Costabile, A.; Sailer, M.; Theis, S.; Rastall, R.A. Prebiotic supplementation of in vitro fecal fermentations inhibits proteolysis by gut bacteria, and host diet shapes gut bacterial metabolism and response to intervention. Appl. Environ. Microbiol. 2019, 85, e02749-18. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Kim, I.H.; Choi, Y.H.; Nam, T.J. A peptide from Porphyra yezoensis stimulates the proliferation of IEC-6 cells by activating the insulin-like growth factor I receptor signaling pathway. Int. J. Mol. Med. 2015, 35, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Remacle-Bonnet, M.; Garrouste, F.; Baillat, G.; Andre, F.; Marvaldi, J.; Pommier, G. Membrane rafts segregate pro- from anti-apoptotic insulin-like growth factor-I receptor signaling in colon carcinoma cells stimulated by members of the tumor necrosis factor superfamily. Am. J. Pathol. 2005, 167, 761–773. [Google Scholar] [CrossRef] [Green Version]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A comprehensive review on MAPK: A promising therapeutic target in cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-K.; Kim, I.-H.; Choi, Y.-H.; Choi, J.-W.; Kim, Y.-M.; Nam, T.-J. The proliferative effects of Pyropia yezoensis peptide on IEC-6 cells are mediated through the epidermal growth factor receptor signaling pathway. Int. J. Mol. Med. 2015, 35, 909–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, M.; Amit, I.; Yarden, Y. Regulation of MAPKs by growth factors and receptor tyrosine kinases. Biochim. Biophys. Acta Bioenerg. 2007, 1773, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhao, G.D.; Shi, Z.; Qi, L.L.; Zhou, L.Y.; Fu, Z.X. The Ras/Raf/MEK/ERK signaling pathway and its role in the occurrence and development of HCC. Oncol. Lett. 2016, 12, 3045–3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopfleisch, R.; Gruber, A.D. Differential expression of cell cycle regulators p21, p27 and p53 in metastasizing canine mammary adenocarcinomas versus normal mammary glands. Res. Vet. Sci. 2009, 87, 91–96. [Google Scholar] [CrossRef]
- Paunovic, B.; Khomenko, T.; Deng, X.; Xiong, X.; Sandor, Z.; Szabo, S. Overexpression of cyclin-dependent kinase (CDK) inhibitors p21 and p27 is a common mechanism of experimental duodenal ulcer and ulcerative colitis. FASEB J. 2010, 24, 1027.4. [Google Scholar] [CrossRef]
- Abdelhedi, O.; Nasri, M. Basic and recent advances in marine antihypertensive peptides: Production, structure-activity relationship and bioavailability. Trends Food Sci. Technol. 2019, 88, 543–557. [Google Scholar] [CrossRef]
- Samarakoon, K.; Jeon, Y.-J. Bio-functionalities of proteins derived from marine algae: A review. Food Res. Int. 2012, 48, 948–960. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Galafat, A.; Sáez, M.I.; Martínez, T.F.; Alarcón, F.J. Partial characterization of protease inhibitors of Ulva ohnoi and their effect on digestive proteases of marine fish. Mar. Drugs 2020, 18, 319. [Google Scholar] [CrossRef]
- Mahomoodally, M.F.; Bibi Sadeer, N.; Zengin, G.; Cziáky, Z.; Jekő, J.; Diuzheva, A.; Sinan, K.I.; Palaniveloo, K.; Kim, D.H.; Rengasamy, K.R.R. In vitro enzyme inhibitory properties, secondary metabolite profiles and multivariate analysis of five seaweeds. Mar. Drugs 2020, 18, 198. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Wang, S.; Jing, L.; Yao, D. Purification and characterisation of a novel angiotensin-I converting enzyme (ACE)-inhibitory peptide derived from the enzymatic hydrolysate of Enteromorpha clathrata protein. Food Chem. 2016, 211, 423–430. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Nova, P.; Pimenta-Martins, A.; Laranjeira Silva, J.; Silva, A.M.; Gomes, A.M.; Freitas, A.C. Health benefits and bioavailability of marine resources components that contribute to health–what’s new? Crit. Rev. Food Sci. Nutr. 2020, 1–13. [Google Scholar] [CrossRef]
- Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366–380. [Google Scholar] [CrossRef]
- Alegría, A.; Garcia-Llatas, G.; Cilla, A. Chapter 1-Static digestion models: General introduction. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 3–12. [Google Scholar] [CrossRef] [Green Version]
- Plank, D.W. In Vitro Method for Estimating In Vivo Protein Digestibility; General Mills Inc.: Minneapolis, MN, USA, 2017; Available online: https://patents.google.com/patent/US9738920B2/en (accessed on 19 July 2020).
- Bohn, T.; Carrière, F.; Day, L.; Deglaire, A.; Egger, L.; Freitas, D.; Golding, M.; Lefeunteun, S.; Macierzanka, A.; Ménard, O.; et al. Correlation between in vitro and in vivo data on food digestion. What can we predict with static in vitro digestion models? Crit. Rev. Food Sci. Nutr. 2017, 58, 2239–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dima, C.; Assadpour, E.; Dima, S.; Jafari, S.M. Bioavailability and bioaccessibility of food bioactive compounds; overview and assessment by in vitro methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2862–2884. [Google Scholar] [CrossRef] [PubMed]
- Alminger, M.; Aura, A.-M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; McDougall, G.J.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef] [Green Version]
- Egger, L.; Ménard, O.; Delgado-Andrade, C.; Alvito, P.; Assunção, R.; Balance, S.; Barberá, R.; Brodkorb, A.; Cattenoz, T.; Clemente, A.; et al. The harmonized INFOGEST in vitro digestion method: From knowledge to action. Food Res. Int. 2016, 88, 217–225. [Google Scholar] [CrossRef]
- Etcheverry, P.; Grusak, M.A.; Fleige, L.E. Application of in vitro bioaccessibility and bioavailability methods for calcium, carotenoids, folate, iron, magnesium, polyphenols, zinc, and vitamins B6, B12, D, and E. Front. Physiol. 2012, 3, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laparra, J.M.; Vélez, D.; Montoro, R.; Barberá, R.; Farré, R. Estimation of arsenic bioaccessibility in edible seaweed by an in vitro digestion method. J. Agric. Food. Chem. 2003, 51, 6080–6085. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.D.; Schricker, B.R.; Rasmussen, R.R.; Van Campen, D. An in vitro method for estimation of iron availability from meals. Am. J. Clin. Nutr. 1981, 34, 2248–2256. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Boisen, S.; Eggum, B. Critical evaluation of in vitro methods for estimating digestibility in simple-stomach animals. Nutr. Res. Rev. 1991, 4, 141–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, D.; Bordoni, A.; Brodkorb, A.; Capozzi, F.; Velickovic, T.C.; Corredig, M.; Cotter, P.D.; De Noni, I.; Gaudichon, C.; Golding, M. An international network for improving health properties of food by sharing our knowledge on the digestive process. Food Dig. 2011, 2, 23–25. [Google Scholar] [CrossRef]
- Afonso, C.; Cardoso, C.; Ripol, A.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Ventura, M.S.; Delgado, I.M.; Coelho, I.; Castanheira, I.; et al. Composition and bioaccessibility of elements in green seaweeds from fish pond aquaculture. Food Res. Int. 2018, 105, 271–277. [Google Scholar] [CrossRef]
- Soukoulis, C.; Tsevdou, M.; Andre, C.M.; Cambier, S.; Yonekura, L.; Taoukis, P.S.; Hoffmann, L. Modulation of chemical stability and in vitro bioaccessibility of beta-carotene loaded in kappa-carrageenan oil-in-gel emulsions. Food Chem. 2017, 220, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of proteins from two marine macroalgae, Ulva sp. and Gracilaria sp. for food application, and evaluating digestibility, amino acid composition and antioxidant properties of the protein concentrates. Food Hydrocoll. 2019, 87, 194–203. [Google Scholar] [CrossRef]
- Guerra, A.; Etienne-Mesmin, L.; Livrelli, V.; Denis, S.; Blanquet-Diot, S.; Alric, M. Relevance and challenges in modeling human gastric and small intestinal digestion. Trends Biotechnol. 2012, 30, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Thuenemann, E.C.; Mandalari, G.; Rich, G.T.; Faulks, R.M. Chapter 6-Dynamic gastric model (DGM). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 47–59. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Marteau, P.; Havenaar, R.; Veld, J.H.H.I.T. A multicompartmental dynamic computer-controlled model simulating the stomach and small intestine. Altern. Lab. Anim. 1995, 23, 197–209. [Google Scholar] [CrossRef]
- Etienne-Mesmin, L.; Livrelli, V.; Privat, M.; Denis, S.; Cardot, J.-M.; Alric, M.; Blanquet-Diot, S. Effect of a new probiotic Saccharomyces cerevisiae strain on survival of Escherichia coli 0157:H7 in a dynamic gastrointestinal model. Appl. Environ. Microbiol. 2011, 77, 1127–1131. [Google Scholar] [CrossRef] [Green Version]
- Larsson, M.; Minekus, M.; Havenaar, R. Estimation of the bioavailability of iron and phosphorus in cereals using a dynamic in vitro gastrointestinal model. J. Sci. Food Agric. 1997, 74, 99–106. [Google Scholar] [CrossRef]
- Verwei, M.; Freidig, A.P.; Havenaar, R.; Groten, J.P. Predicted serum folate concentrations based on in vitro studies and kinetic modeling are consistent with measured folate concentrations in humans. J. Nutr. 2006, 136, 3074–3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo Anson, N.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Antioxidant and anti-inflammatory capacity of bioaccessible compounds from wheat fractions after gastrointestinal digestion. J. Cereal Sci. 2010, 51, 110–114. [Google Scholar] [CrossRef]
- Torres-Escribano, S.; Denis, S.; Blanquet-Diot, S.; Calatayud, M.; Barrios, L.; Vélez, D.; Alric, M.; Montoro, R. Comparison of a static and a dynamic in vitro model to estimate the bioaccessibility of As, Cd, Pb and Hg from food reference materials Fucus sp. (IAEA-140/TM) and Lobster hepatopancreas (TORT-2). Sci. Total Environ. 2011, 409, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Bellmann, S.; Miyazaki, K.; Chonan, O.; Ishikawa, F.; Havenaar, R. Fucoidan from Cladosiphon okamuranus Tokida added to food has no adverse effect on availability for absorption of divalent minerals in the dynamic multicompartmental model of the upper gastrointestinal tract. Food Digestion 2014, 5, 19–25. [Google Scholar] [CrossRef]
- Blanquet, S.; Zeijdner, E.; Beyssac, E.; Meunier, J.-P.; Denis, S.; Havenaar, R.; Alric, M. A dynamic artificial gastrointestinal system for studying the behavior of orally administered drug dosage forms under various physiological conditions. Pharm. Res. 2004, 21, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Wickham, M.; Faulks, R.; Mann, J.; Mandalari, G. The design, operation, and application of a dynamic gastric model. Dissolut. Technol. 2012, 19, 15–22. [Google Scholar] [CrossRef]
- Vardakou, M.; Mercuri, A.; Barker, S.; Craig, D.; Faulks, R.; Wickham, M. Achieving antral grinding forces in biorelevant in vitro models: Comparing the USP Dissolution Apparatus II and the Dynamic Gastric Model with human in vivo data. AAPS PharmSciTech. 2011, 12, 620–626. [Google Scholar] [CrossRef] [Green Version]
- Marciani, L.; Gowland, P.A.; Fillery-Travis, A.; Manoj, P.; Wright, J.; Smith, A.; Young, P.; Moore, R.; Spiller, R.C. Assessment of antral grinding of a model solid meal with echo-planar imaging. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G844–G849. [Google Scholar] [CrossRef] [Green Version]
- Lea, T. Chapter 9-Epithelial cell models; general introduction. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 95–102. [Google Scholar] [CrossRef] [Green Version]
- Lv, Q.; He, Q.; Wu, Y.; Chen, X.; Ning, Y.; Chen, Y. Investigating the bioaccessibility and bioavailability of cadmium in a cooked rice food matrix by using an 11-day rapid Caco-2/HT-29 co-culture cell model combined with an in vitro digestion model. Biol. Trace Elem. Res. 2019, 190, 336–348. [Google Scholar] [CrossRef]
- Kuhre, R.E.; Wewer Albrechtsen, N.J.; Deacon, C.F.; Balk-Møller, E.; Rehfeld, J.F.; Reimann, F.; Gribble, F.M.; Holst, J.J. Peptide production and secretion in GLUTag, NCI-H716, and STC-1 cells: A comparison to native L-cells. J. Mol. Endocrinol. 2016, 56, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In vitro digestion of chestnut and quebracho tannin extracts: Antimicrobial effect, antioxidant capacity and cytomodulatory activity in swine intestinal IPEC-J2 cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Glahn, R.P.; Wien, E.M.; van Campen, D.R.; Miller, D.D. Caco-2 cell iron uptake from meat and casein digests parallels in vivo studies: Use of a novel in vitro method for rapid estimation of iron bioavailability. J. Nutr. 1996, 126, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Trigo, J.P.; Engström, N.; Steinhagen, S.; Juul, L.; Harrysson, H.; Toth, G.B.; Pavia, H.; Scheers, N.; Undeland, I. In vitro digestibility and Caco-2 cell bioavailability of sea lettuce (Ulva fenestrata) proteins extracted using pH-shift processing. Food Chem. 2021, 356, 129683. [Google Scholar] [CrossRef]
- Flores, S.R.L.; Dobbs, J.; Dunn, M.A. Mineral nutrient content and iron bioavailability in common and Hawaiian seaweeds assessed by an in vitro digestion/Caco-2 cell model. J. Food Compos. Anal. 2015, 43, 185–193. [Google Scholar] [CrossRef]
- Domínguez-González, M.R.; Chiocchetti, G.M.; Herbello-Hermelo, P.; Vélez, D.; Devesa, V.; Bermejo-Barrera, P. Evaluation of iodine bioavailability in seaweed using in vitro methods. J. Agric. Food. Chem. 2017, 65, 8435–8442. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J.A. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- Hayes, M. Food proteins and bioactive peptides: New and novel sources, characterisation strategies and applications. Foods 2018, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Popova, A.; Mihaylova, D. Antinutrients in plant-based foods: A review. Open Biotechnol. J. 2019, 13, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Fabiano, A.; Brilli, E.; Mattii, L.; Testai, L.; Moscato, S.; Citi, V.; Tarantino, G.; Zambito, Y. Ex vivo and in vivo study of Sucrosomial® iron intestinal absorption and bioavailability. Int. J. Mol. Sci. 2018, 19, 2722. [Google Scholar] [CrossRef] [Green Version]
- Ussing, H.H. The active ion transport through the isolated frog skin in the light of tracer studies. Acta Physiol. Scand. 1949, 17, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awati, A.; Rutherfurd, S.M.; Plugge, W.; Reynolds, G.W.; Marrant, H.; Kies, A.K.; Moughan, P.J. Ussing chamber results for amino acid absorption of protein hydrolysates in porcine jejunum must be corrected for endogenous protein. J. Sci. Food Agric. 2009, 89, 1857–1861. [Google Scholar] [CrossRef]
- Westerhout, J.; Wortelboer, H.; Verhoeckx, K. Chapter 24-Ussing chamber. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 263–273. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Liu, Y.; Zhao, B.; Tang, M.; Dong, H.; Zhang, L.; Lv, B.; Wei, L. Ex vivo and in situ approaches used to study intestinal absorption. J. Pharmacol. Toxicol. Methods 2013, 68, 208–216. [Google Scholar] [CrossRef]
- Agar, W.T.; Hird, F.J.R.; Sidhu, G.S. The uptake of amino acids by the intestine. Biochim. Biophys. Acta 1954, 14, 80–84. [Google Scholar] [CrossRef]
- Hillgren, K.M.; Kato, A.; Borchardt, R.T. In vitro systems for studying intestinal drug absorption. Med. Res. Rev. 1995, 15, 83–109. [Google Scholar] [CrossRef]
- Nossol, C.; Barta-Böszörményi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.-K.; Rothkötter, H.-J. Comparing two intestinal porcine epithelial cell lines (IPECs): Morphological differentiation, function and metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef] [Green Version]
- Ripken, D.; Hendriks, H. Chapter 23-Porcine ex vivo intestinal segment model. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 255–262. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Pantoja-Feliciano, I.G.; Zachos, N.C.; Racicot, K. Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Roeselers, G.; Ponomarenko, M.; Lukovac, S.; Wortelboer, H.M. Ex vivo systems to study host–microbiota interactions in the gastrointestinal tract. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 101–113. [Google Scholar] [CrossRef]
- van de Merbel, A.F.; van der Horst, G.; van der Mark, M.H.; van Uhm, J.I.M.; van Gennep, E.J.; Kloen, P.; Beimers, L.; Pelger, R.C.M.; van der Pluijm, G. An ex vivo tissue culture model for the assessment of individualized drug responses in prostate and bladder cancer. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Ripken, D.; van der Wielen, N.; Wortelboer, H.M.; Meijerink, J.; Witkamp, R.F.; Hendriks, H.F.J. Steviol glycoside rebaudioside a induces glucagon-like peptide-1 and peptide yy release in a porcine ex vivo intestinal model. J. Agric. Food. Chem. 2014, 62, 8365–8370. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.-M.; Maukonen, J. Chapter 25-One compartment fermentation model. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 281–292. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Tiihonen, K.; Mäkeläinen, H.; Rautonen, N.; Hasselwander, O.; Sworn, G. Chapter 5-Non-starch polysaccharides in the gastrointestinal tract. In Designing Functional Foods; McClements, D.J., Decker, E.A., Eds.; Woodhead Publishing: Cambridge, UK, 2009; pp. 126–147. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Use of a three-stage continuous culture system to study the effect of mucin on dissimilatory sulfate reduction and methanogenesis by mixed populations of human gut bacteria. Appl. Environ. Microbiol. 1988, 54, 2750–2755. [Google Scholar] [CrossRef] [Green Version]
- Molly, K.; Vande Woestyne, M.; Verstraete, W. Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Grootaert, C.; Marzorati, M.; Possemiers, S.; Verstraete, W.; Gérard, P.; Rabot, S.; Bruneau, A.; El Aidy, S.; Derrien, M. Microbial community development in a dynamic gut model is reproducible, colon region specific, and selective for Bacteroidetes and Clostridium cluster IX. Appl. Environ. Microbiol. 2010, 76, 5237–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possemiers, S.; Rabot, S.; Espín, J.C.; Bruneau, A.; Philippe, C.; González-Sarrías, A.; Heyerick, A.; Tomás-Barberán, F.A.; De Keukeleire, D.; Verstraete, W. Eubacterium limosum activates isoxanthohumol from hops (Humulus lupulus L.) into the potent phytoestrogen 8-prenylnaringenin in vitro and in rat intestine. J. Nutr. 2008, 138, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F. Incorporating a mucosal environment in a dynamic gut model results in a more representative colonization by Lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzorati, M.; Verhelst, A.; Luta, G.; Sinnott, R.; Verstraete, W.; de Wiele, T.V.; Possemiers, S. In vitro modulation of the human gastrointestinal microbial community by plant-derived polysaccharide-rich dietary supplements. Int. J. Food Microbiol. 2010, 139, 168–176. [Google Scholar] [CrossRef]
- Fu, Y.; Yin, N.; Cai, X.; Du, H.; Wang, P.; Sultana, M.S.; Sun, G.; Cui, Y. Arsenic speciation and bioaccessibility in raw and cooked seafood: Influence of seafood species and gut microbiota. Environ. Pollut. 2021, 280, 116958. [Google Scholar] [CrossRef]
- Calatayud, M.; Xiong, C.; Du Laing, G.; Raber, G.; Francesconi, K.; van de Wiele, T. Salivary and gut microbiomes play a significant role in in vitro oral bioaccessibility, biotransformation, and intestinal absorption of arsenic from food. Environ. Sci. Technol. 2018, 52, 14422–14435. [Google Scholar] [CrossRef]
- Boever, P.D.; Wouters, R.; Vermeirssen, V.; Boon, N.; Verstraete, W. Development of a six-stage culture system for simulating the gastrointestinal microbiota of weaned infants. Microb. Ecol. Health Dis. 2001, 13, 111–123. [Google Scholar] [CrossRef]
- Van de Wiele, T.; Van den Abbeele, P.; Ossieur, W.; Possemiers, S.; Marzorati, M. Chapter 27-The simulator of the human intestinal microbial ecosystem (SHIME®). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 305–317. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Walton, G.; Jiang, L.; Plummer, S.; Garaiova, I.; Gibson, G. Comparative analysis of intestinal tract models. Annu. Rev. Food Sci. Technol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Cueva, C.; Peláez, C.; Martínez-Cuesta, M.C.; Requena, T. Development of human colonic microbiota in the computer-controlled dynamic SIMulator of the GastroIntestinal tract SIMGI. LWT 2015, 61, 283–289. [Google Scholar] [CrossRef]
- Barroso, E.; Cueva, C.; Peláez, C.; Martínez-Cuesta, M.C.; Requena, T. Chapter 28-The computer-controlled multicompartmental dynamic model of the gastrointestinal system SIMGI. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 319–327. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.; Zihler, A.; Chassard, C.; Lacroix, C. Advances and perspectives in in vitro human gut fermentation modeling. Trends Biotechnol. 2011, 30, 17–25. [Google Scholar] [CrossRef]
- Nissen, L.; Casciano, F.; Gianotti, A. Intestinal fermentation in vitro models to study food-induced gut microbiota shift: An updated review. FEMS Microbiol. Lett. 2020, 367. [Google Scholar] [CrossRef]
- Wissenbach, D.K.; Oliphant, K.; Rolle-Kampczyk, U.; Yen, S.; Höke, H.; Baumann, S.; Haange, S.B.; Verdu, E.F.; Allen-Vercoe, E.; von Bergen, M. Optimization of metabolomics of defined in vitro gut microbial ecosystems. Int. J. Med. Microbiol. 2016, 306, 280–289. [Google Scholar] [CrossRef]
- Marzorati, M.; Vanhoecke, B.; De Ryck, T.; Sadaghian Sadabad, M.; Pinheiro, I.; Possemiers, S.; Van den Abbeele, P.; Derycke, L.; Bracke, M.; Pieters, J.; et al. The HMI™ module: A new tool to study the Host-Microbiota Interaction in the human gastrointestinal tract in vitro. BMC Microbiol. 2014, 14, 133. [Google Scholar] [CrossRef] [Green Version]
- Venema, K.; van den Abbeele, P. Experimental models of the gut microbiome. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 115–126. [Google Scholar] [CrossRef]
- Wood, R.J.; Tamura, T. Methodological issues in assessing bioavailability of nutrients and other bioactive substances in dietary supplements: Summary of workshop discussion. J. Nutr. 2001, 131, 1396S–1398S. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Jahreis, G.; Hausmann, W.; Kiessling, G.; Franke, K.; Leiterer, M. Bioavailability of iodine from normal diets rich in dairy products-results of balance studies in women. Exp. Clin. Endocrinol. Diabetes 2001, 109, 163–167. [Google Scholar] [CrossRef]
- Lu, Y.L.; Li, S.J.; Liu, G.Y.; Li, X.C.; Yang, D.; Jia, J.Y.; Zhang, M.Q.; Zheng, H.C.; Yu, C.; Zhu, F.; et al. Oral bioavailability and mass balance studies of a novel anti-arrhythmic agent sulcardine sulfate in Sprague-Dawley rats and beagle dogs. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 453–459. [Google Scholar] [CrossRef]
- Shin, B.S.; Hong, S.H.; Bulitta, J.B.; Hwang, S.W.; Kim, H.J.; Lee, J.B.; Yang, S.D.; Kim, J.E.; Yoon, H.S.; Kim, D.J.; et al. Disposition, oral bioavailability, and tissue distribution of zearalenone in rats at various dose levels. J. Toxicol. Environ. Health A 2009, 72, 1406–1411. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.M.; Costa, N.M.B.; Nutti, M.R.; Tako, E.; Martino, H.S.D. Advantages and limitations of in vitro and in vivo methods of iron and zinc bioavailability evaluation in the assessment of biofortification program effectiveness. Crit. Rev. Food Sci. Nutr. 2018, 58, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- García, Y.; Díaz-Castro, J. Advantages and disadvantages of the animal models v. in vitro studies in iron metabolism: A review. Animal 2013, 7, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.N.; Rathbone, M.J.; Burgess, D.; Huynh, M. Breakout session summary from AAPS/CRS joint workshop on critical variables in the in vitro and in vivo performance of parenteral sustained release products. J. Control. Release 2010, 142, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.M.; Ferrucci, L.M.; Tangrea, J.A.; Schatzkin, A. Epidemiological and clinical studies of nutrition. Semin. Oncol. 2010, 37, 282–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Liu, C.; Wang, Y.; Wang, C.; Xie, M.; Qian, Y.; Fu, L. Application of in vitro and in vivo models in the study of food allergy. Food Sci. Hum. Wellness 2018, 7, 235–243. [Google Scholar] [CrossRef]
- Gueven, N.; Spring, K.J.; Holmes, S.; Ahuja, K.; Eri, R.; Park, A.Y.; Fitton, J.H. Micro RNA expression after ingestion of fucoidan; a clinical study. Mar. Drugs 2020, 18, 143. [Google Scholar] [CrossRef] [Green Version]
- Ikeda-Ohtsubo, W.; López Nadal, A.; Zaccaria, E.; Iha, M.; Kitazawa, H.; Kleerebezem, M.; Brugman, S. Intestinal microbiota and immune modulation in Zebrafish by fucoidan from Okinawa mozuku (Cladosiphon okamuranus). Front. Nutr. 2020, 7. [Google Scholar] [CrossRef]
- Roche-Lima, A.; Carrasquillo-Carrión, K.; Gómez-Moreno, R.; Cruz, J.M.; Velázquez-Morales, D.M.; Rogozin, I.B.; Baerga-Ortiz, A. The presence of genotoxic and/or pro-inflammatory bacterial genes in gut metagenomic databases and their possible link with inflammatory bowel diseases. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed, Cladosiphon okamuranus. Mar. Drugs 2015, 13, 48–64. [Google Scholar] [CrossRef]
- Kadena, K.; Tomori, M.; Iha, M.; Nagamine, T. Absorption study of mozuku fucoidan in Japanese volunteers. Mar. Drugs 2018, 16, 254. [Google Scholar] [CrossRef] [Green Version]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a fucoidan-specific antibody and measurement of fucoidan in serum and urine by sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, S.; Touvrey-Loiodice, M.; Poulet, L.; Drouillard, S.; Vincentelli, R.; Henrissat, B.; Skjåk-Bræk, G.; Helbert, W. Ancient acquisition of “alginate utilization loci” by human gut microbiota. Sci. Rep. 2018, 8, 8075. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Xu, H.; Wei, C.; Jiang, T.; Qin, S.; Zhang, W.; Cao, Y.; Hu, C.; Zhang, F.; Qiao, D.; et al. Horizontal transfer of a novel soil agarase gene from marine bacteria to soil bacteria via human microbiota. Sci. Rep. 2016, 6, 34103. [Google Scholar] [CrossRef] [PubMed]
- Pudlo, N.A.; Pereira, G.V.; Parnami, J.; Cid, M.; Markert, S.; Tingley, J.P.; Unfried, F.; Ali, A.; Campbell, A.; Urs, K.; et al. Extensive transfer of genes for edible seaweed digestion from marine to human gut bacteria. bioRxiv 2020, 2020.06.10.142968. [Google Scholar] [CrossRef]
- Hehemann, J.-H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef]
- Thomas, F.; Barbeyron, T.; Tonon, T.; Génicot, S.; Czjzek, M.; Michel, G. Characterization of the first alginolytic operons in a marine bacterium: From their emergence in marine Flavobacteriia to their independent transfers to marine Proteobacteria and human gut Bacteroides. Environ. Microbiol. 2012, 14, 2379–2394. [Google Scholar] [CrossRef]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.; Pourshahidi, L.K.; et al. Impact of a (poly) phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Sęczyk, Ł.; Świeca, M.; Kapusta, I.; Gawlik-Dziki, U. Protein-phenolic interactions as a factor affecting the physicochemical properties of white bean proteins. Molecules 2019, 24, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buitimea-Cantúa, N.E.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Phenolic–protein interactions: Effects on food properties and health benefits. J. Med. Food 2018, 21, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cheng, Z.; Wang, Y.; Fu, L. Dietary protein-phenolic interactions: Characterization, biochemical-physiological consequences, and potential food applications. Crit. Rev. Food Sci. Nutr. 2020, 1–27. [Google Scholar] [CrossRef]
- Imbs, T.; Zvyagintseva, T. Phlorotannins are polyphenolic metabolites of brown algae. Russ. J. Mar. Biol. 2018, 44, 263–273. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, D.; Sun, J.; Liu, X.; Jiang, L.; Guo, H.; Ren, F. Interaction of plant phenols with food macronutrients: Characterisation and nutritional–physiological consequences. Nutr. Res. Rev. 2013, 27, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mignet, N.; Seguin, J.; Chabot, G.G. Bioavailability of polyphenol liposomes: A challenge ahead. Pharmaceutics 2013, 5, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Jakobek, L.; Matić, P. Non-covalent dietary fiber-polyphenol interactions and their influence on polyphenol bioaccessibility. Trends Food Sci. Technol. 2019, 83, 235–247. [Google Scholar] [CrossRef]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of in vitro digestion on composition, bioaccessibility and antioxidant activity of food polyphenols-a non-systematic review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef]
- Sookkasem, A.; Chatpun, S.; Yuenyongsawad, S.; Wiwattanapatapee, R. Alginate beads for colon specific delivery of self-emulsifying curcumin. J. Drug Deliv. Sci. Technol. 2015, 29, 159–166. [Google Scholar] [CrossRef]
- Hussain, M.B.; Hassan, S.; Waheed, M.; Javed, A.; Farooq, M.A.; Tahir, A. Chapter 5-Bioavailability and metabolic pathway of phenolic compounds. In Plant Physiological Aspects of Phenolic Compounds; Soto-Hernández, M., Palma-Tenango, M., García-Mateos, R., Eds.; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar] [CrossRef] [Green Version]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Carregosa, D.; Carecho, R.; Figueira, I.; Santos, C.N. Low-molecular weight metabolites from polyphenols as effectors for attenuating neuroinflammation. J. Agric. Food Chem. 2020, 68, 1790–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Hu, M. Natural polyphenol disposition via coupled metabolic pathways. Expert Opin. Drug Metab. Toxicol. 2007, 3, 389–406. [Google Scholar] [CrossRef]
- Suetsuna, K.; Maekawa, K.; Chen, J.-R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, D.C.; Lebrun, I. Cryptides: Buried secrets in proteins. Peptides 2007, 28, 2403–2410. [Google Scholar] [CrossRef]
- Hayes, M.; García-García, M.; Fitzgerald, C.; Lafarga, T. Chapter 27-Seaweed and milk derived bioactive peptides and small molecules in functional foods and cosmeceuticals. In Biotechnology of Bioactive Compounds: Sources and Applications; Gupta, V.K., Tuohy, M.G., Eds.; John Wiley & Sons: Oxford, UK, 2015; pp. 669–691. [Google Scholar] [CrossRef]

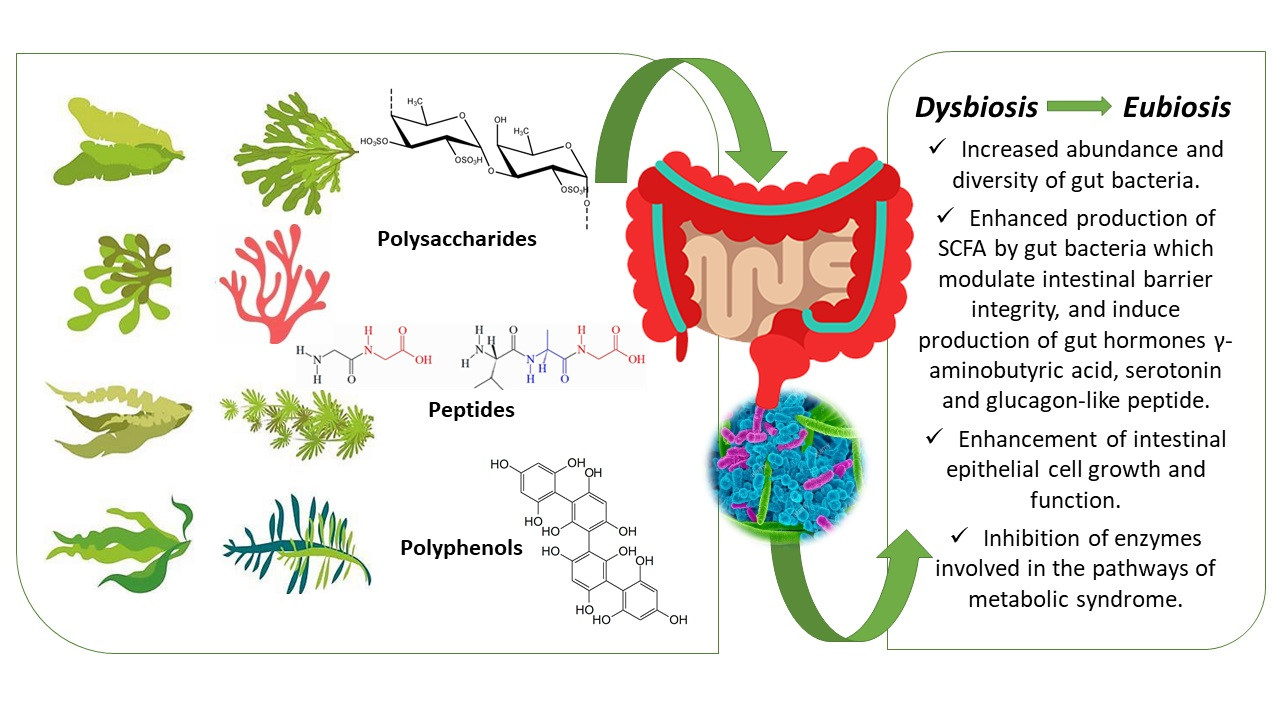{kind=link}
| Polysaccharide | Seaweed | Extraction Method | Study Type | Statistically Significant Effects | Ref. |
|---|---|---|---|---|---|
| * (i) Crude polysaccharide-rich extract (>1 kDa) (CE) (ii) Depolymerised crude extract (>1 kDa) (DE) | L. digitata | (i) (CE) Hot acid and ethanol precipitation (0.1 M HCl) (ii) (DE) Fenton’s reaction with iron and hydrogen peroxide | Simulated in vitro colonic digestion | After 24 h fermentation, compared to cellulose control:
Ratio of propionate to acetate beneficially reduced by CE and DE (both p < 0.05) after 24, 36, and 48 h. | [147] |
| * Porphyran, ulvan and laminarin | Pyropia, Ulva and Laminaria | Ethanol (80%) | Simulated in vitro colonic digestion | After 24 h fermentation, growth of bacterial genera compared to fructooligosaccharide (FOS) control: Porphyran increased Lactobacilli (10.7%, p < 0.05). Ulvan increased Bacteroides (6.7%, p < 0.05). Laminarin increased Bifidobacteria (8.3%, p < 0.05) and Bacteroides (13.8%, p < 0.05). Negative results: no significant increase at 24 h in total SCFA, butyrate, lactate or acetate by laminarin, ulvan or porpyran compared to FOS. | [132] |
| * (i) Crude extract fraction (CF) (ii) Low MW fraction (LPF) (iii) High MW fraction (HPF) | E. radiata | (i) Enzymatic (Viscozyme-β-glucanase, hemicellulase, arabanase, xylanase) (ii and iii) Viscozyme and ethanol precipitation | Simulated in vitro colonic digestion | Increases (log10 cells/mL) after 24 h fermentation (all p < 0.05 compared to controls):
| [10] |
| * (i) Low MW polysaccharide (LMW) (primarily laminarin) (ii) High MW polysaccharide acidic water extract (HMW) (primarily fucoidan and alginate) (iii) High MW polysaccharide water and ethanol precipitate (HMWW) (primarily fucoidan and alginate) | E. radiata | (i) Enzymatic (cellulase) (ii) Acidic water (pH 4.5) (iii) Water and ethanol precipitation | Simulated in vitro colonic digestion | 24 h post fermentation (all differences p < 0.05): (i) LMW increased Bifidobacteria from 5.51 ± 0.15 log10 cells/mL (in cellulose fermented control) to 6.55 ± 0.08 log10 cells/mL; Lactobacillus from 4.73 ± 0.13 (cellulose) to 5.28 ± 0.19 log10 cells/mL and Bacteroidetes from 5.09 ± 0.06 (cellulose) to 6.02 ± 0.09 log10 cells/mL. Negative results: no significant increase by LMW on populations of F. prausnitzii, Clostridium leptum, Ruminococcus bromii, E. coli or Enterococcus. (ii) HMW increased C. coccoides from 5.74 ± 0.75 (cellulose) to 7.07 ± 0.04 log10 cells/mL, E. coli from 6.09 ± 0.41 (cellulose) to 7.52 ± 0.07 log10 cells/mL and Enterococcus from 5.02 ± 0.31 (cellulose) to 6.63 ± 0.11 log10 cells/mL. Negative results: no significant increase by HMW in any other bacterial populations. (iii) HMWW increased E. coli from 6.09 ± 0.41 (cellulose) to 7.01 ± 0.17 log10 cells/mL and Enterococcus from 5.02 ± 0.31 (cellulose) to 5.80 ± 0.33 log10 cells/mL. HMWW also had a negative effect on several bacterial populations—Bifidobacteria reduced from 5.51 ± 0.15 (cellulose) to 3.21 ± 0.61 log10 cells/mL, Bacteroidetes from 5.09 ± 0.06 (cellulose) to 4.08 ± 0.12 log10 cells/mL, Lactobacillus 4.73 ± 0.13 log10 cells/mL (cellulose) to not detected (ND), C. coccoides from 5.74 ± 0.75 log10 cells/mL (cellulose) to ND, C. leptum from 6.23 ± 0.28 log10 cells/mL (cellulose) to ND and R. bromii from 6.20 ± 0.06 (cellulose) to 4.87 ± 0.29 log10 cells/mL. SCFA increases in seaweed ferments vs. cellulose control after 24 h (all p < 0.05):
| [148] |
| ** (i) Polysaccharide fraction (PF) (primarily fucoidan and alginate) (ii) Whole seaweed (WS) | E. radiata | (i) Enzymatic (Viscozyme) (ii) Whole dried E. radiata | In vivo trial with healthy Sprague-Dawley rats (7 d, 5% PF or 5% WS added to feed) | After 7 days supplementation (all differences p < 0.05): Reduction in potentially pathogenic Enterococci in WS group (6.04 ± 0.09 log10 cells/mL) vs. control (5.59 ± 0.08 log10 cells/mL) Increase in butyrate-producing F. prausnitzii in PF group (5.32 ± 0.11 log10 cells/mL) vs. control (4.87 ± 0.11 log10 cells/mL) 2-fold increase in caecal digesta mass 1.36 ± 0.17 (PF) vs. 0.60 ± 0.06 g/100 g BM (control) Putrefactive microbial products reduced (all values µg/g caecal digesta):
| [149] |
| * (i) conventional chemical extraction (CCE) (11.9% fucoidan) (ii) microwave-assisted extraction (MAE) (5.71% fucoidan) (iii) ultrasound-assisted extraction (UAE) (4.56% fucoidan) (iv) enzyme-assisted extraction (EAE) (3.89% fucoidan) | A. nodosum | (i, ii, and iii) Ethanol followed by acidic water (0.01 M HCl) (iv) Cellulase, acetate buffer (pH 4.5) | L. casei and L. delbrueckii ssp. bulgaricus broth cultures, 3.75% (v/v). A. nodosum extracts added at 0.1%, 0.3% and 0.5% (w/v) | All differences p < 0.05 compared to non-supplemented control medium: Increase in L. delbrueckii ssp. bulgaricus by CCE, MAE, UAE and EAE at 0.1%, 0.3% and 0.5%. Increase (24.5%) in L. casei only by MAE at 0.5% inclusion. Negative results:
| [87] |
| * Crude sulphated polysaccharide (716 kDa) (90% galactose, 9.07% sulphate) | C. pilulifera | Acidic extraction (0.0.1 M HCl) and ethanol precipitation | Simulated in vitro saliva, gastric, small intestinal and colonic digestion | After 24 h, all differences p < 0.05 compared to inulin control: Increase in Bacteroides, Parabacteroides, Megamonas and Veillonella. Increase in total SCFA (22.17 ± 0.82 mmol/L) vs. control (16.17 mmol/L ± 0.39). Negative results:
| [150] |
| * (i) Polysaccharides (SJP) (138 kDa) (Fucose:galactose:glucuronic acid:mannose, molar ratio of 4.1:3.6:1.2: 1.0). (ii) Oligosaccharides (SJO) | S. japonica | (i) Methanol, dichloromethane, water and ethanol (ii) Methanol, dichloromethane, water and ethanol, followed by 0.6 M HCl | Simulated in vitro colonic digestion | After 24 h, all differences p < 0.05 compared to FOS control
| [91] |
| ** Crude sulphated polysaccharide (SP) (28.807 kDa) (Galactose (59.7%), galacturonic acid (19.8%), xylose (7.1%) and sulphate (8.8%)) | G. pacificum | Ultrasound-assisted water extraction followed by ethanol, acetone and petroleum precipitation | In vivo trial with lincomycin hydrochloride induced diarrhoeal mice (9 days, 75 mg SP/kg BM) | After 9 d, seaweed polysaccharide group vs. non-supplemented normal recovery group (all differences p < 0.05): Increase in beneficial Bacteroides, Oscillospira and Bifidobacterium. Decrease in Parabacteroides, Sutterella and AF12. Reduction in inflammatory cytokines, TNF-α, IL-1β and IL-2. Improved (lower) diarrhoea status scores, water intake, and less weight loss. Increase in total SCFA, acetate and propionate. | [151] |
| ** Fucoidan (300 kDa) (60% fucose, 14.3% sulphate) | C. okamuranus | Method not specified | In vivo trial with Traf3 ip2-mutant psoriasis mice (fucoidan diet group n = 14, normal diet group n = 9, 63 days, 1% fucoidan added to feed) | Fucoidan group vs. cellulose control group (all differences p < 0.05). After 56 days:
| [89] |
| ** Laminarin and fucoidan (10% laminarin,8% fucoidan and 82% ash) | Laminaria hyperborea | Method not specified | In vivo trial (10 pregnant sows/treatment) (10 g/days seaweed extract from day 107 of gestation until weaning (day 26)) and ex vivo lipopolysaccharide (LPS) immunological challenge | Compared with non-supplemented group, seaweed extract supplemented (SWE) sows had:
Piglets suckling SWE sows had:
| [101] |
| Polyphenol | Seaweed | Extraction Method | Study Type | Statistically Significant Effects | Ref. |
|---|---|---|---|---|---|
| * Phlorotannin enriched fraction | E. radiata | Ethanol (90%) | Simulated in vitro colonic digestion | Increases (all p < 0.05) in Bacteroidetes (6.52 ± 0.04 log10 cells/mL) compared to the cellulose control (6.40 ± 0.05 log10 cells/mL); F. prausnitzii (6.57 ± 0.05 log10 cells/mL) compared to cellulose and inulin controls (6.17 ± 0.04 and 6.07 ± 0.06 log10 cells/mL, respectively); C. coccoides (7.97 ± 0.05 log10 cells/mL) compared to inulin and cellulose controls (7.57 ± 0.06 and 7.40 ± 0.05 log10 cells/mL, respectively); and E. coli (8.09 ± 0.02 log10 cells/mL) compared to inulin and cellulose controls (6.81 ± 0.03 and 6.94 ± 0.03 log10 cells/mL, respectively). | [10] |
| ** Polyphenols (3 kDa) (luteolin-6-c-glucoside, regiolone, neoeriocitrin and estr-5(10)-ene-3,17-diol) | E. prolifera | Ultrasound assisted ethanol extraction (55%) and ultrafiltration (3 kDa) | In vivo trial with diabetic mice (4 weeks, 300 mg polyphenol extract/kg BM/day) | Reduction after 14 days (p < 0.05) in mean BM of E. prolifera-fed diabetic group compared to model diabetic group. Reduction after 28 days (p < 0.05) in mean fasting blood glucose levels of E. prolifera-fed diabetic group and glucose tolerance increased (p < 0.05) compared to the model diabetic group. Increase in Alistipes (p < 0.05) in E. prolifera-fed diabetic group compared to model diabetic group. Hypoglycaemic effect via increase (p < 0.01) in phosphatidylinositol 3-kinase and suppression (p < 0.05) of c-Jun N-terminal kinase in E. prolifera-fed diabetic group livers compared to model diabetic group. | [170] |
| ** Polyphenol-rich fraction (primarily phlorotannins, phenolic acids and gallocatechin derivatives) | L. trabeculata | Microwave assisted methanol extraction, solvent fractionation and macroporous resin adsorption separation | In vivo trial with diabetic rats (4 weeks, 200 mg/day phlorotannin extract/kg BM) | Increase in genera of the phylum Bacteroidetes in the PE group compared to the DC group: Odoribacter (p < 0.008), Muribaculum (p < 0.005), Alistipes (p < 0.006), Lachnospiraceae (p < 0.015) and Parabacteroides (p < 0.022). Decrease in Proteobacteria, and ratio of Firmicutes to Bacteroidetes (p < 0.05 PE vs. DC group). Increase in total SCFA (491.31 ± 10.39 (DC), 1276.34 ± 16.86 μg/g (PE) (p < 0.01)), acetic acid (377.77 ± 3.46 (DC), 1202.49 ± 11.55 μg/g (PE) (p < 0.01)) and butyric acid (10.18 ± 0.58 (DC), 39.77 ± 1.85 μg/g (PE) (p < 0.01)). Reduction in the PE group versus the DC group in: fasting blood glucose (10.55 ± 0.94 vs. 13.99 ± 0.87 mmol/L (p < 0.05)), serum insulin (14.69 ± 0.11 vs. 17.70 ± 0.22 mU/L (p < 0.01)), HOMA-IR insulin resistance value (6.89 ± 0.42 vs. 11.01 ± 0.98 (p < 0.01)), total cholesterol (4.92 ± 0.14 vs. 5.64 ± 0.16 mmol/L (p < 0.01)), triglycerides (0.99 ± 0.04 vs. 1.43 ± 0.10 mmol/L (p < 0.01)), LDL cholesterol (0.68 ± 0.03 vs. 1.06 ± 0.06 (p < 0.01)), glycated serum protein (2.15 ± 0.16 vs. 2.74 ± 0.15 (p < 0.01)) and non-esterified fatty acids (1.86 ± 0.05 vs. 2.02 ± 0.11 mmol/L (p < 0.05)). | [210] |
| (i) * Phlorotannin (HMW > 10 kDa) (ii) Phlorotannin (LMW 1–10 kDa) | A. nodosum | Ethanol | (a) In vitro gastrointestinal digestion and colonic fermentation (b) H2O2 induced DNA damage in HT-29 colon cancer cells | (a) Reduction in MW of phlorotannins (89.9% HMW, 62.0% LMW) by colonic fermentation, compared to enzymatic gastric digestion (5.4% HMW, 52.8% LMW), suggesting phlorotannins may potentially be metabolised by human gut bacteria. (b) Compared to the control, HMW and LMW phlorotannin extracts at a concentration of 500 μg/mL inhibited (p < 0.01) HT-29 colon cancer cell proliferation (number of cells by division), HMW inhibited (p < 0.05) HT-29 cell growth (mass accumulation) at concentrations of 250 and 500 μg/mL. H2O2 induced DNA damage in HT-29 cells reduced by post-gastric digested HMW extract (p < 0.01) and HMW and LMW post-colonic fermented extracts (both p < 0.001). | [216] |
| Seaweed | Extraction Method | Amino Acid Sequence | Bioactivity | Ref. |
|---|---|---|---|---|
| * † U. lactuca | Enzymatic (Papain), MWCO filtration, preparative RP-HPLC and in silico enzyme cleavage simulation | (i) Ala-Thr-Lys-Pro-Ala-Asn (ii) Ser-Gly-Ala-Ala-Ser-Ala-Ser-Gly-Ala-Ala (iii) Ala-Gly-Gly-Pro-Asn-Gln-Pro-Pro-Asn (iv) Ala-Ala-Asn-Ile-Thr-Val-Pro-Ala-Ala-Asn (v) Glu-Ala-Glu-Pro-Ala-Glu-Ala-Ala (vi) Gly-Ala-Ala-Pro-Thr-Pro-Pro-Ser-Pro-Pro-Pro-Ala-Thr-Lys-Pro-Ser-Thr-Pro-Pro-Lys-Pro-Pro-Thr (vii) Pro-Pro-Asn-Pro-Pro-Asn-Pro-Pro-Asn Amino acid sequences not defined: (a) crude seaweed protein (b) full peptide hydrolysate (c) 1 kDa-UFH (ultra-filtered hydrolysate) (d) 3 kDa-UFH (e) 10 kDa-UFH | Peptides (i) to (vii) ACE-I, DPP-IV, and enzyme 3-hydroxy-3-methyl-glutaryl-CoA reductase inhibition (in silico predictive activity) In vitro ACE-I inhibitory activity (%) (all assayed at conc. of 1mg/mL): (a) crude seaweed protein 79.87 ± 0.18% (b) full peptide hydrolysate 82.37 ± 0.05% (c) 1 kDa-UFH (ultra-filtered hysrolysate) 93.03 ± 0.87% (d) 3 kDa-UFH 86.64 ± 2.17% (e) 10 kDa-UFH 88.12 ± 0.02% | [9] |
| * P. palmata | Enzymatic (Papain) | Ile-Arg-Leu-Ile-Ile-Val-Leu-Met-Pro-Ile-Leu-Met-Ala | Renin inhibition (58.97 ± 1.26% inhibition in vitro at 1 mg/mL) | [217] |
| * P. palmata | Enzymatic (Protease) | (i) Ile-Leu-Ala-Pro (ii) Leu-Leu-Ala-Pro (iii) Met-Ala-Gly-Val-Asp-His-Ile | DPP-IV inhibition IC50 values in vitro: (i) 43.40 ± 1.40 μM (ii) 53.67 ± 0.82 μM (iii) 159.37 ± 13.67 μM | [218] |
| * P. palmata | Enzymatic (Papain) | Asn-Ile-Gly-Lys | PAF-AH inhibition IC50 value in vitro 2.32 ± 2.12 mM | [219] |
| * Porphyra (Laver—species not specified) | Enzymatic (Viscozyme, Alcalase, Neutrase, Pepsin and Trypsin) | (i) Gly-Gly-Ser-Lys (ii) Glu-Leu-Ser | α-amylase inhibition IC50 values in vitro: (i) 2.58 ± 0.08 mM (ii) 2.62 ± 0.05 mM | [220] |
| * P. palmata | Thermolysin hydrolysis | (i) Leu-Arg-Tyr (ii) Val-Tyr-Arg-Thr | ACE-I inhibition IC50 values in vitro: (i) 0.044 μM (ii) 0.14 μM | [228] |
| *,** U. pinnatifida | Enzymatic (Protease) | (i) Val-Tyr (ii) Ile-Tyr (iii) Phe-Tyr (iv) Ile-Trp (v) Ala-Trpvi) Val-Trp (vii) Leu-Trp | ACE-I inhibition IC50 values in vitro: (i) 35.2 μM (ii) 6.1 μM (iii) 42.3 μM (iv) 1.5 μM (v) 18.8 μM(vi) 3.3 μM (vii) 23.6 μM In vivo antihypertensive effect in spontaneously hypertensive rats (single oral dose, 1 mg/kg of BW). Blood pressure decreases (pre-administration vs. 9 h post): (i) Val-Tyr (228.2 ± 3.4 vs. 206.7 ± 9.5 mmHg) (p < 0.05) (ii) Ile-Tyr (205.6 ± 5.2 vs. 184.3 ± 4.5 mmHg) (p < 0.05) (iii) Phe-Tyr (208.7 ± 4.4 vs. 193.0 ± 5.1 (p < 0.01) (iv) Ile-Trp (213.3 ± 3.4 vs. 199.5 ± 5.9) (p < 0.05) | [229] |
| * U. pinnatifida | Enzymatic (Pepsin) | (i) Ala-Ile-Tyr-Lys (ii) Tyr-Lys-Tyr-Tyr (iii) Lys-Phe-Tyr-Gly (iv) Tyr-Asn-Lys-Leu | ACE-I inhibition IC50 values in vitro:((i) 213 μM (ii) 64.2 μM (iii) 90.5 μM (iv) 21.0 μM | [230] |
| * P. palmata | Enzymatic (Protease) | Ser-Asp-Ile-Thr-Arg-Pro-Gly-Gly-Asn-Met | Antioxidant activity after simulated gastrointestinal digestion: Oxygen radical absorbance capacity 152.43 ± 2.73 nM Trolox equivalents (TE)/µmol peptide and ferric reducing antioxidant power activity 21.23 ± 0.90 nM TE/µmol peptide, | [231] |
| Peptide | Seaweed | Study Type | Statistically Significant Effects | Ref. |
|---|---|---|---|---|
| * Ala-Leu-Glu-Gly-Gly-Lys-Ser-Ser-Gly-Gly-Gly-Glu-Ala-Thr-Arg-Asp-Pro-Glu-Pro-Thr | P. yezoensis | In vitro rat intestinal epithelial cells—investigating the modulation of cell differentiation. | At concentrations of 125–1000 ng/mL, the peptide, dose-depenently (p < 0.05):
| [244] |
| * Ala-Leu-Glu-Gly-Gly-Lys-Ser-Ser-Gly-Gly-Gly-Glu-Ala-Thr-Arg-Asp-Pro-Glu-Pro-Thr | P. yezoensis | In vitro rat intestinal epithelial cells—investigating the epidermal growth factor receptor signalling pathway and Ras/Raf-p42/p44 MAPK signalling pathway, mediating signal transduction from cell surface to nucleus. | At concentrations of 125–1000 ng/mL, the peptide dose-dependently(p < 0.05):
| [247] |
| In Vitro Bioaccessibility Methods | Advantages | Limitations |
|---|---|---|
| Solubility and Dialysability |
|
|
| Static GI models |
|
|
| INFOGEST static in vitro model |
|
|
| Dynamic GI models |
|
|
| Cell models |
|
|
| Ex vivo bioavailability methods |
|
|
| In vitro fermentation models |
|
|
| In vivo bioavailability methods |
|
|
| Seaweed | Polyphenol | Extraction Method | Study Type | Observed Effects | Ref. |
|---|---|---|---|---|---|
| *; *** A. nodosum | Phlorotannins | Ethanolic crude phlorotannin extract (CE) and high-molecular-weight (HMW) fraction (>10 kDa) by tangential flow ultrafiltration. Combined as CE (58%) and HMW (42%) | (i) In vitro gastrointestinal enzymatic digestion, colonic fermentation, and dialysis to simulate absorption into the bloodstream. (ii) Cross-sectional human clinical trial (12 male, 12 female, healthy 18–65 years-old) (one capsule 101.89 mg phlorotannins). Blood and urine collected (0 to 24 h). | Phlorotannin metabolites detected in 15 of 24 participants after 24 h (total phlorotannins ranged from 0.011–7.76 μg/mL in blood plasma and from 0.15–33.52 μg/mL in urine). | [206] |
| *** A. nodosum | Phlorotannins | Ethanol CE extract and HMW fraction (>10 kDa) by tangential flow ultrafiltration. Combined as CE (57%) and HMW (43%) | 24 week crossover study (8 weeks, 100 mg phlorotannin/d, or placebo capsule) (39 men, 41 women, mean BMI 30.2, mean age 42.7 years-old), 8 weeks washout phase, then repeat 8 weeks intervention or placebo treatment. Plasma and urine collected before/after each phase (0, 8, 16 and 24 weeks). | Polyphenol metabolites (0.5–11.8 mg/day total polyphenols) detected in 36 of 78 participants. | [358] |
| Seaweed | Peptide | Extraction Method | Study Type | Statistically Significant Effects Post-Digestion | Ref. |
|---|---|---|---|---|---|
| ** U. pinnatifida | (i) Tyr-His (ii) Lys-Tyr (iii) Phe-Tyr (iv) Ile-Tyr | Hot water | In vivo study in spontaneously hypertensive rats. (a) Single oral administration of each dipeptide (50 mg/kg BM) (b) Continuous administration for 7 days (10 mg/day/kg BM) | (a) All dipeptides decreased (p < 0.05) blood pressure after single oral dose:
| [374] |
| *; ** U. pinnatifida | (i) Ile-Trp (ii) Val-Trp (iii) Ile-Tyr (iv) Ala-Trp (v) Leu-Trp (vi) Val-Tyr (vii) Phe-Tyr | Enzymatic (Protease from Bacillus stearothermophilus) and HPLC separation to butanol-soluble fractions |
| (a) No loss in ACE-I inhibitory activity post in vitro digestion.IC50 values: (i) Ile-Trp 1.5 µM (ii) Val-Trp 3.3 µM (iii) Ile-Tyr 6.1 µM (iv) Ala-Trp 18.8 µM (v) Leu-Trp 23.6 µM (vi) Val-Tyr 35.2 µM (vii) Phe-Tyr 42.3 µM (b) In vivo antihypertensive effect in spontaneously hypertensive rats (single oral dose, all 1 mg/kg of BW). Blood pressure decreases (pre-administration vs. 9h post): (i) Val-Tyr (228.2 ± 3.4 vs. 206.7 ± 9.5 mmHg) (p < 0.05) (ii) Ile-Tyr (205.6 ± 5.2 vs. 184.3 ± 4.5 mmHg) (p < 0.05) (iii) Phe-Tyr (208.7 ± 4.4 vs. 193.0 ± 5.1 (p < 0.01) (iv) Ile-Trp (213.3 ± 3.4 vs. 199.5 ± 5.9) (p < 0.05) Captopril control (238.7 ± 6.9 vs. 224.9 ± 4.1 (p < 0.05) | [229] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shannon, E.; Conlon, M.; Hayes, M. Seaweed Components as Potential Modulators of the Gut Microbiota. Mar. Drugs 2021, 19, 358. https://doi.org/10.3390/md19070358
Shannon E, Conlon M, Hayes M. Seaweed Components as Potential Modulators of the Gut Microbiota. Marine Drugs. 2021; 19(7):358. https://doi.org/10.3390/md19070358
Chicago/Turabian StyleShannon, Emer, Michael Conlon, and Maria Hayes. 2021. "Seaweed Components as Potential Modulators of the Gut Microbiota" Marine Drugs 19, no. 7: 358. https://doi.org/10.3390/md19070358
APA StyleShannon, E., Conlon, M., & Hayes, M. (2021). Seaweed Components as Potential Modulators of the Gut Microbiota. Marine Drugs, 19(7), 358. https://doi.org/10.3390/md19070358