
Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin

 , ,
, , 
Abstract
:1. Introduction
2. Results
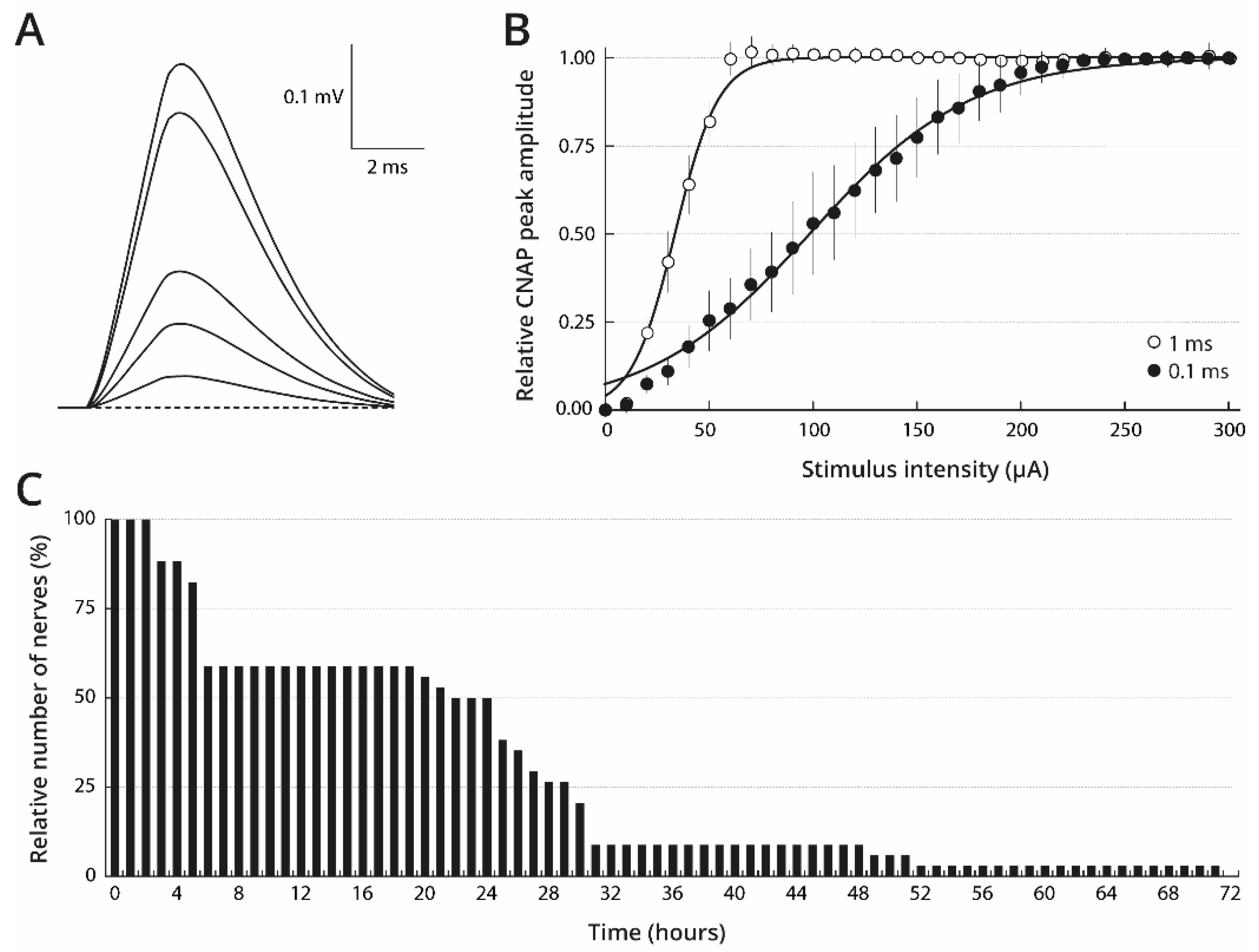
2.1. Stimulation and Maintaining Conditions of Isolated Oyster Nerves to Obtain Consistent CNAP
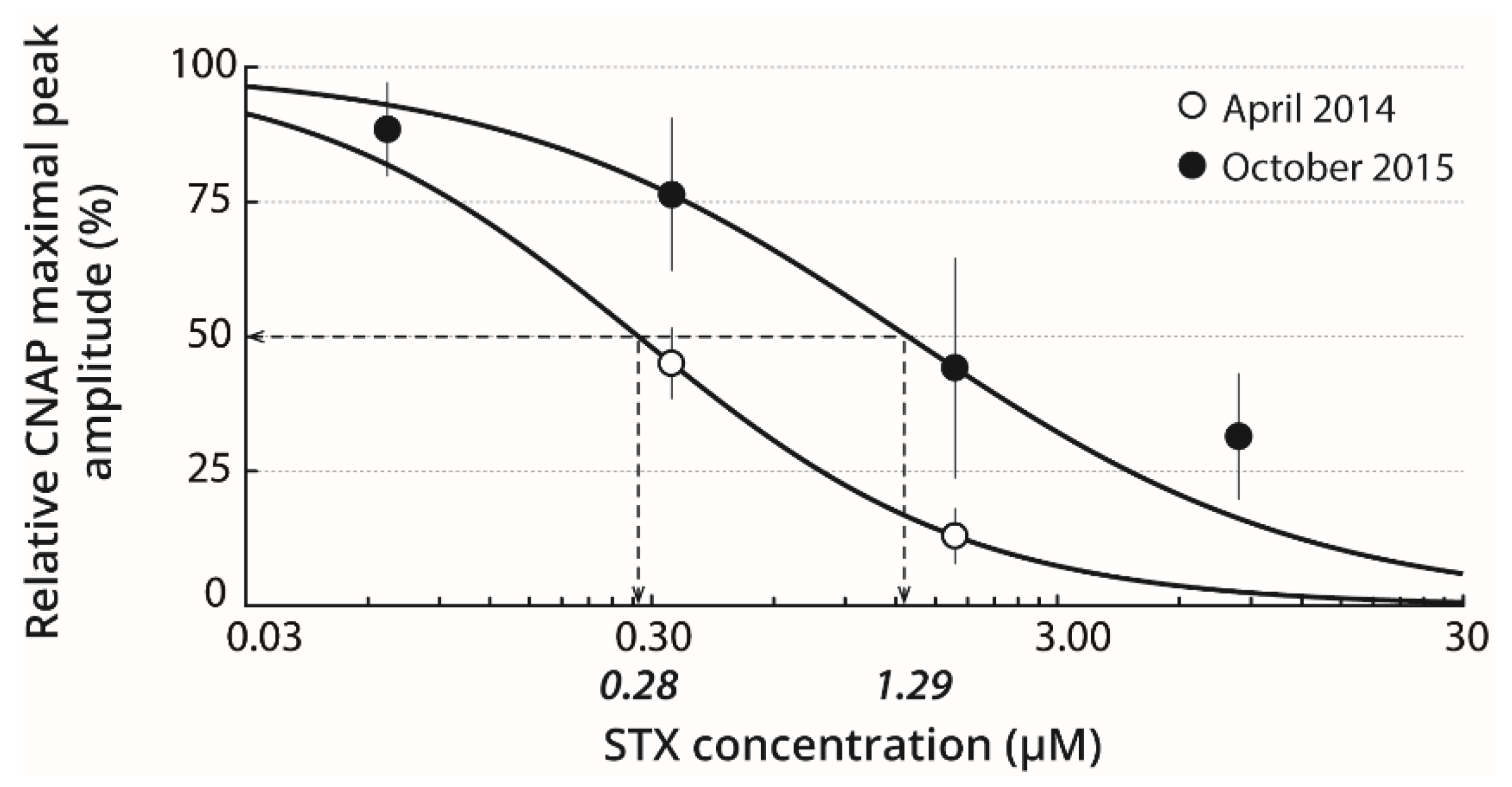
2.2. Basal STX Sensitivity of Isolated Nerves
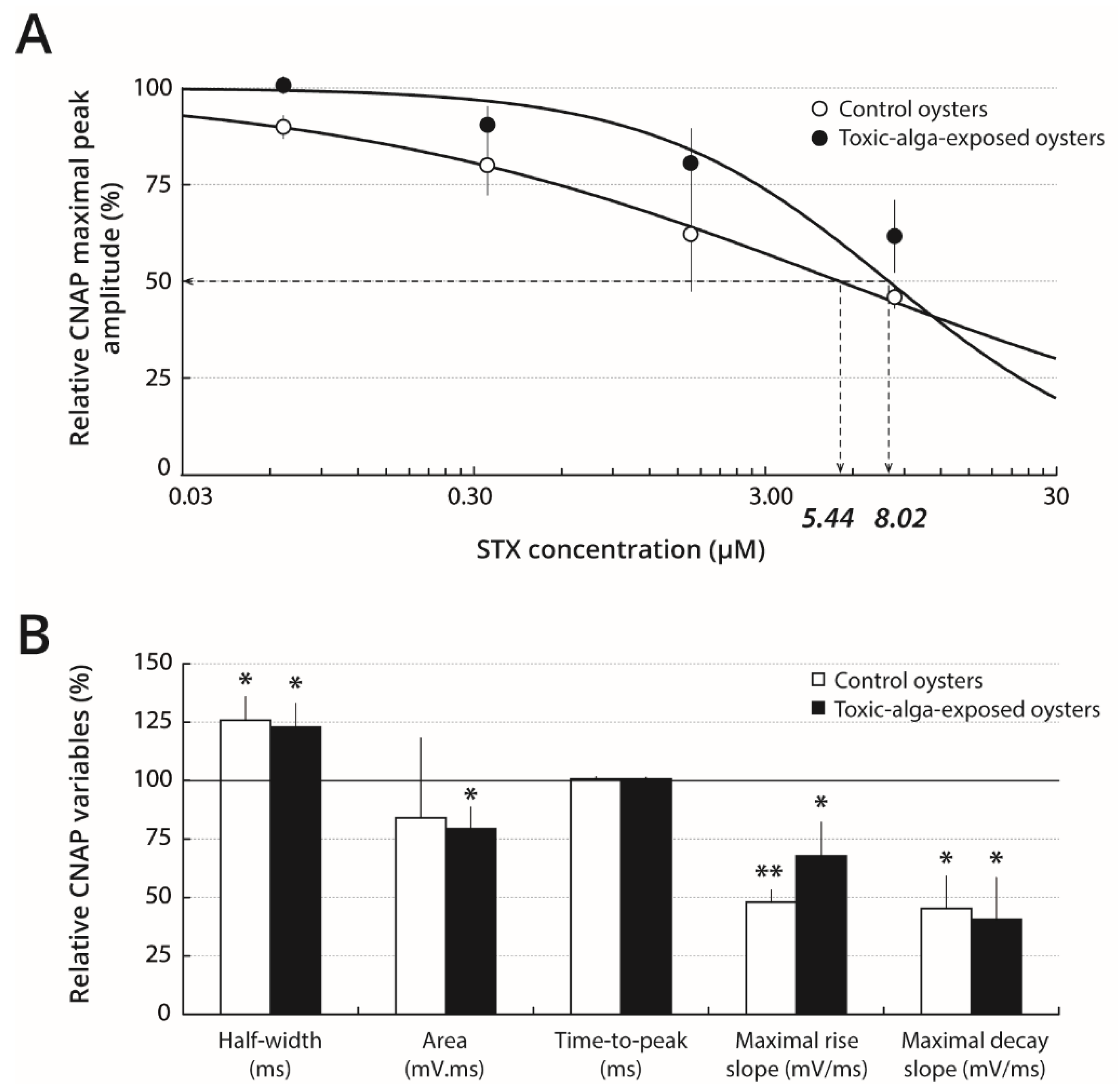
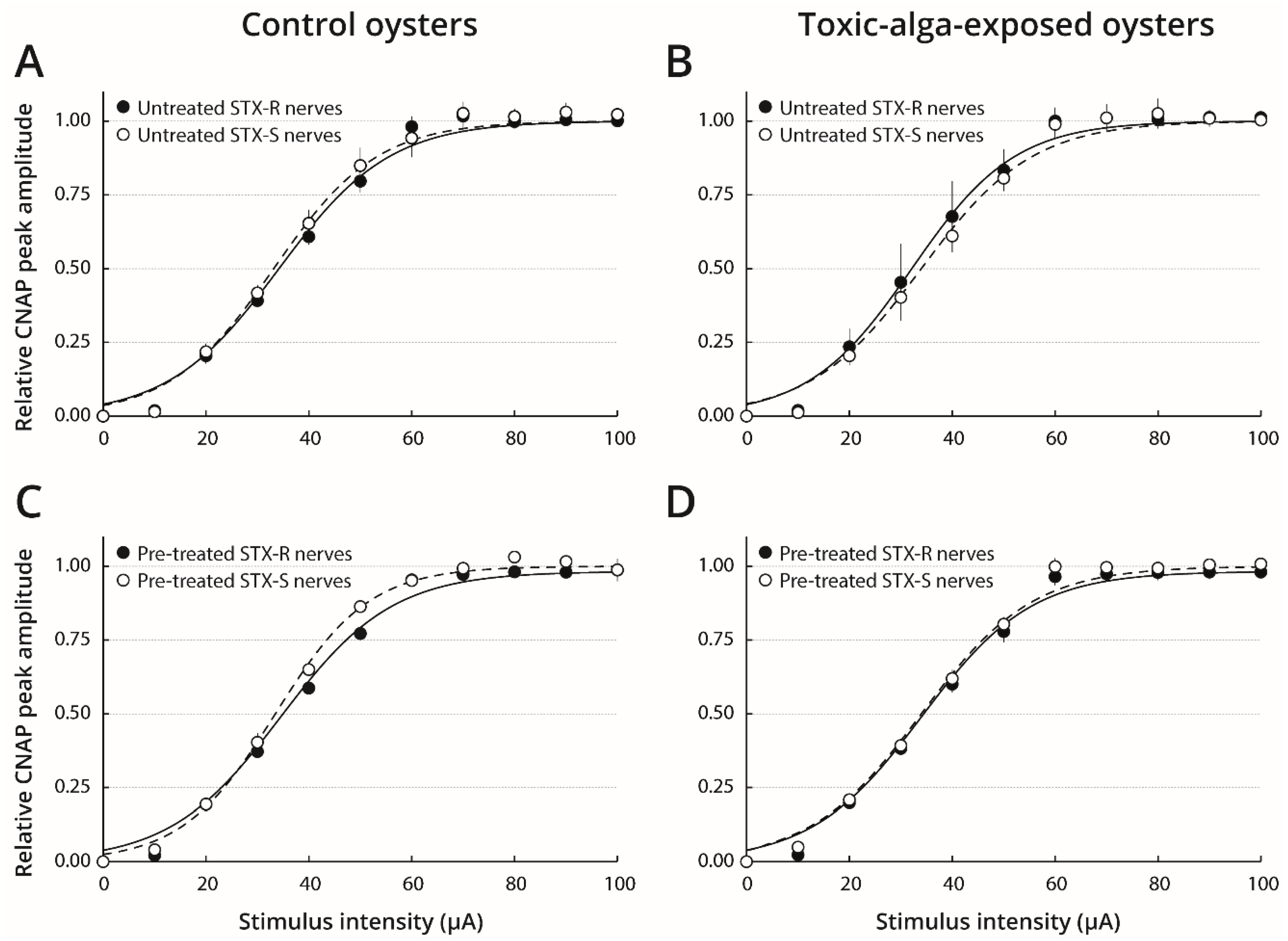
2.3. STX Sensitivity of Nerves Isolated from Oysters Pre-Exposed to Toxic or Non-Toxic Microalgae
2.4. TTX Sensitivity of Oyster Nerves
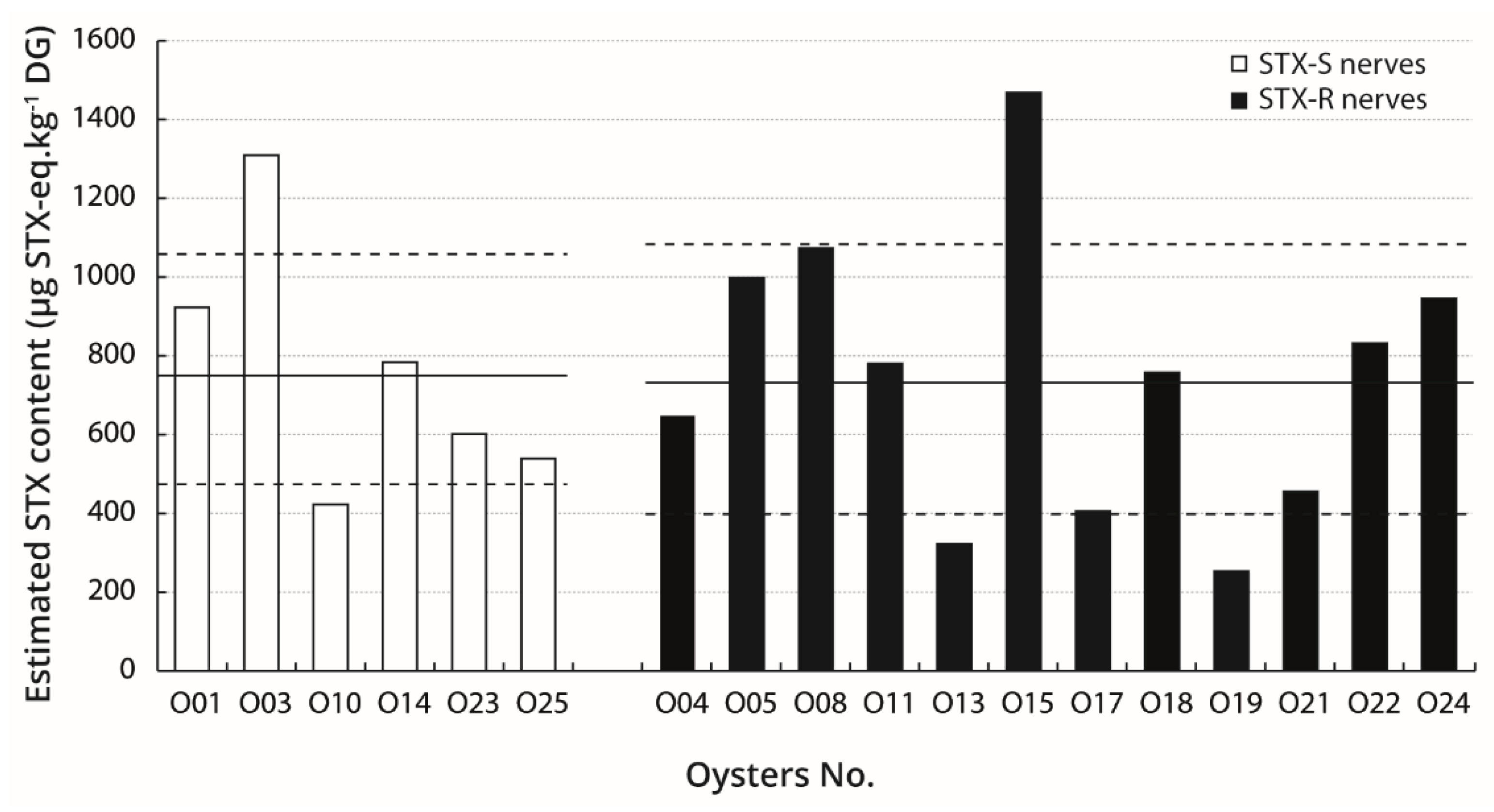
2.5. Relation Between STX Bio-Accumulation in Digestive Glands and Sensitivity of Nerves
3. Discussion
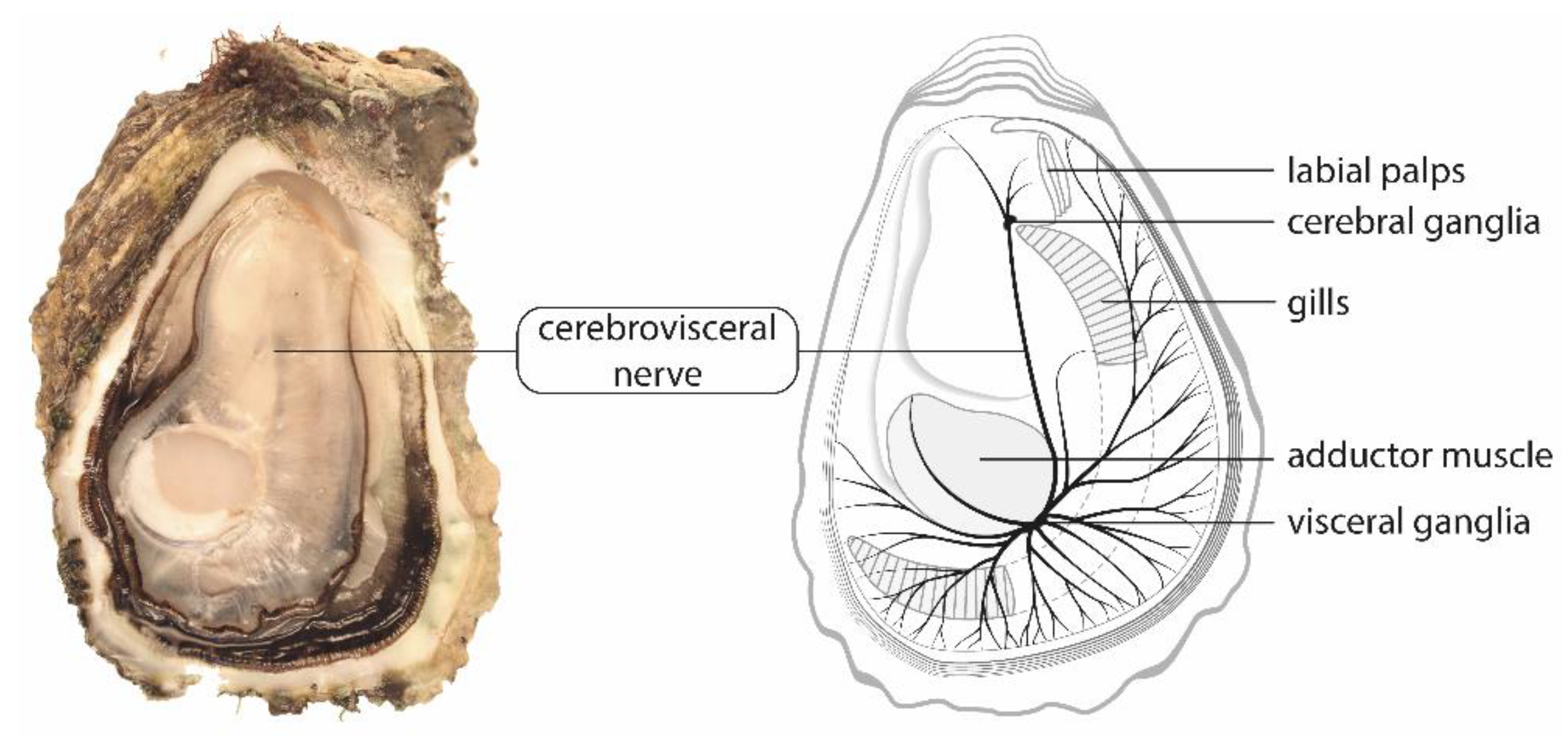
3.1. Cerebrovisceral Nerves of Pacific Oysters Are Suitable for Electrophysiological and Pharmacological Studies
3.2. Cerebrovisceral Nerves of Pacific Oysters Have Intermediate STX Sensitivity Compared to Other Bivalves
3.3. STX Sensitivity of Cerebrovisceral Nerves of C. gigas Oysters Varies Between Individuals
3.4. The Variability of PST Load in Oysters Does Not Appear Related to Sensitivity of Nerves
3.5. A Pre-Exposure of Pacific Oysters to PST-Producer Dinoflagellates Decreases STX Sensitivity of Nerves
4. Materials and Methods
4.1. Oysters
4.2. Microalgal Cultures
4.3. Experimental Design for Exposure to Toxic and Non-Toxic Microalgae
4.4. ELISA Assay for Estimated STX Content analyses
4.5. Electrophysiological Recordings
4.6. Toxins
4.7. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Guallar, C.; Bacher, C.; Chapelle, A. Global and local factors driving the phenology of Alexandrium minutum (Halim) blooms and its toxicity. Harmful Algae 2017, 67, 44–60. [Google Scholar] [CrossRef] [Green Version]
- Lassus, P.; Baron, R.; Garen, P.; Truquet, P.; Masselin, P.; Bardouil, M.; Leguay, D.; Amzil, Z. Paralytic shellfish poison outbreaks in the Penzé estuary: Environmental factors affecting toxin uptake in the oyster. Crassostrea gigas. Aquat. Living Resour. 2004, 17, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.-Z.; Zhang, S.-F.; Zhang, Y.; Lin, L. Paralytic shellfish toxin biosynthesis in cyanobacteria and dinoflagellates: A molecular overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef]
- Catterall, W.A. From ionic currents to molecular mechanisms: The structure and function of voltage-gated sodium channels. Neuron 2000, 26, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Zhodi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Sellner, K.G.; Doucette, G.J.; Kirkpatrick, G.J. Harmful algal blooms: Causes, impacts and detection. J. Ind. Microbiol. Biotechnol. 2003, 30, 383–406. [Google Scholar] [CrossRef] [PubMed]
- Bricelj, V.M.; Connell, L.; Konoki, K.; MacQuarrie, S.P.; Scheuer, T.; Catterall, W.A.; Trainer, V.L. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: Occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Boullot, F.; Castrec, J.; Bidault, A.; Dantas, N.; Payton, L.; Perrigault, M.; Tran, D.; Amzil, Z.; Boudry, P.; Soudant, P.; et al. Molecular characterization of voltage-gated sodium channels and their relations with paralytic shellfish toxin bioaccumulation in the Pacific oyster Crassostrea gigas. Mar. Drugs 2017, 15, 21. [Google Scholar] [CrossRef]
- Haberkorn, H.; Tran, D.; Massabuau, J.C.; Ciret, P.; Savar, V.; Soudant, P. Relationship between valve activity, microalgae concentration in the water and toxin accumulation in the digestive gland of the Pacific oyster Crassostrea gigas exposed to Alexandrium minutum. Mar. Pollut Bull. 2011, 62, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Twarog, B.M.; Hidaka, T.; Yamagushi, H. Resistance to tetrodotoxin and saxitoxin in nerves of bivalve molluscs. Toxicon 1972, 10, 273–278. [Google Scholar] [CrossRef]
- Twarog, B.M. "Immunity" to paralytic shellfish toxin in bivalve molluscs. In Proceedings of the Second International Coral Reef Symposium, Brisbane, Australia, 22 June–2 July 1973; Cameron, A.M., Cambell, B.M., Cribb, A.B., Endean, R., Jell, J.S., Jones, O.A., Mather, P., Talbot, F.H., Eds.; The Great Barrier Reef Committee: Brisbane, Australia, 1974; Volume 1, pp. 505–512. [Google Scholar]
- Kvitek, R.G.; Beitler, M.K. Relative insensitivity of butter clam neurons to saxitoxin: A pre-adaptation for sequestering paralytic shellfish poisoning toxins as a chemical defense. Mar. Ecol. Prog. Ser. 1991, 69, 47–54. [Google Scholar] [CrossRef]
- Daigo, K.; Noguchi, T.; Miwa, A.; Nobufumi, K.; Hashimoto, K. Resistance of nerves from certain toxic crabs to paralytic shellfish poison and tetrodotoxin. Toxicon 1988, 26, 485–490. [Google Scholar] [CrossRef]
- Kao, C.Y.; Fuhrman, F.A. Differentiation of the actions of tetrodotoxin and saxitoxin. Toxicon 1967, 5, 25–34. [Google Scholar] [CrossRef]
- Anderson, P.A. Properties and pharmacology of a TTX-insensitive Na+ current in neurones of the jellyfish Cyanea capillata. J. Exp. Biol. 1987, 133, 231–248. [Google Scholar] [CrossRef]
- Johansen, J.; Kleinhaus, A.L. Saxitoxin differentiates between two types of Na+-dependent potentials in the Retzius cell of hirudinid leeches. J. Exp. Biol. 1987, 131, 351–363. [Google Scholar] [CrossRef]
- Hort, V.; Arnich, N.; Guérin, T.; Lavison-Bompard, G.; Nicolas, M. First Detection of Tetrodotoxin in Bivalves and Gastropods from the French Mainland Coasts. Toxins 2020, 12, 599. [Google Scholar] [CrossRef]
- Patria, F.P.; Pekar, H.; Zuberovic-Muratovic, A. Multi-Toxin Quantitative Analysis of Paralytic Shellfish Toxins and Tetrodotoxins in Bivalve Mollusks with Ultra-Performance Hydrophilic Interaction LC-MS/MS-An In-House Validation Study. Toxins 2020, 12, 452. [Google Scholar] [CrossRef]
- Dhanji-Rapkova, M.; Turner, A.D.; Baker-Austin, C.; Huggett, J.F.; Ritchie, J.M. Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas). Mar. Drugs 2021, 19, 84. [Google Scholar] [CrossRef] [PubMed]
- Chapelle, A.; Le Gac, M.; Labry, C.; Siano, R.; Quere, J.; Caradec, F.; Le Bec, C.; Nezan, E.; Doner, A.; Gouriou, J. The Bay of Brest (France) a new risky site for toxic Alexandrium minutum blooms and PSP shellfish contamination. Harmful Algae News 2015, 51, 4–5. [Google Scholar]
- Pousse, É.; Flye-Sainte-Marie, J.; Alunno-Bruscia, M.; Hégaret, H.; Jean, F. Sources of paralytic shellfish toxin accumulation variability in the Pacific oyster Crassostrea gigas. Toxicon 2018, 144, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Walne, P.R. Experiments in the large-scale culture of the larvae Ostrea edulis. Fish. Investig. Lond. Ser. II 1966, 25, 1–53. [Google Scholar]
- Benoit, E.; Charpentier, G.; Mateu, L.; Luzzati, V.; Kado, R. Electrophysiology of the olfactory nerve of the pike, Esox lucius: A pilot study on optimal experimental conditions. Cybium 2000, 24, 241–248. [Google Scholar]
- Luzzati, V.; Benoit, E.; Charpentier, G.; Vachette, P. X-ray scattering study of pike olfactory nerve: Elastic, thermodynamic and physiological properties of the axonal membrane. J. Mol. Biol. 2004, 343, 199–212. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maximal Peak Amplitude (mV) | Half-Width (ms) | Area (mV.ms) | Time-to-Peak (ms) | Maximal Rise Slope (mV/ms) | Maximal Decay Slope (mV/ms) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before SPS | After SPS | Before SPS | After SPS | Before SPS | After SPS | Before SPS | After SPS | Before SPS | After SPS | Before SPS | After SPS | |
| Control oysters (n = 21 nerves) | 0.374 ± 0.097 | 0.358 ± 0.081 | 5.124 ± 0.794 | 5.025 ± 0.607 | 2.088 ± 0.336 | 1.995 ± 0.316 | 56.079 ± 0.396 | 55.995 ± 0.249 | 0.250 ± 0.090 | 0.252 ± 0.076 | 0.116 ± 0.047 | 0.105 ± 0.038 |
| Toxic-alga-exposed oysters (n = 25 nerves) | 0.269 ± 0.102 ** | 0.303 ± 0.095 * | 6.201 ± 1.156 ** | 5.780 ± 1.026 ** | 1.822 ± 0.530 | 1.930 ± 0.405 | 56.364 ± 0.346 * | 56.265 ± 0.308 ** | 0.179 ± 0.076 ** | 0.193 ± 0.084 * | 0.075 ± 0.036 ** | 0.087 ± 0.047 |
| Maximal Peak Amplitude (mV) | Half-Width (ms) | Area (mV.ms) | Time-to-Peak (ms) | Maximal Rise Slope (mV/ms) | Maximal Decay Slope (mV/ms) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| STX-S Nerves | STX-R Nerves | STX-S Nerves | STX-R Nerves | STX-S Nerves | STX-R Nerves | STX-S Nerves | STX-R Nerves | STX-S Nerves | STX-R Nerves | STX-S Nerves | STX-R Nerves | |
| Control oysters (n = 8–10 nerves) | 0.393 ± 0.051 | 0.317 ± 0.069 * | 4.705 ± 0.387 | 5.294 ± 0.652 * | 1.935 ± 0.292 | 1.923 ± 0.339 | 55.992 ± 0.301 | 56.004 ± 0.250 | 0.274 ± 0.068 | 0.176 ± 0.073 * | 0.126 ± 0.030 | 0.086 ± 0.033 * |
| Toxic-alga-exposed oysters (n = 6–12 nerves) | 0.387 ± 0.074 | 0.339 ± 0.083 | 5.154 ± 0.374 | 5.445 ± 0.915 | 1.891 ± 0.248 | 2.035 ± 0.377 | 56.389 ± 0.354 | 56.139 ± 0.308 | 0.215 ± 0.298 | 0.196 ± 0.038 | 0.133 ± 0.035 | 0.100 ± 0.050 |
| I50 (μA) | S (μA−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| Untreated STX-S | Pre-Treated STX-S | Untreated STX-R | Pre-Treated STX-R | Untreated STX-S | Pre-treated STX-S | Untreated STX-R | Pre-Treated STX-R | |
| Control oysters (n = 8–10 nerves) | 33.16 ± 0.95 | 33.47 ± 1.06 | 34.18 ± 0.51 | 34.59 ± 0.62 | 10.10 ± 0.59 | 9.09 ± 0.24 | 10.77 ± 0.33 | 10.81 ± 0.28 |
| Toxic-alga-exposed oysters (n = 6–12 nerves) | 33.97 ± 0.74 | 34.10 ± 0.82 | 32.12 ± 3.81 | 34.06 ± 0.97 | 10.82 ± 0.23 | 10.58 ± 0.18 | 10.04 ± 2.10 | 10.70 ± 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boullot, F.; Fabioux, C.; Hégaret, H.; Boudry, P.; Soudant, P.; Benoit, E. Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin. Mar. Drugs 2021, 19, 380. https://doi.org/10.3390/md19070380
Boullot F, Fabioux C, Hégaret H, Boudry P, Soudant P, Benoit E. Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin. Marine Drugs. 2021; 19(7):380. https://doi.org/10.3390/md19070380
Chicago/Turabian StyleBoullot, Floriane, Caroline Fabioux, Hélène Hégaret, Pierre Boudry, Philippe Soudant, and Evelyne Benoit. 2021. "Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin" Marine Drugs 19, no. 7: 380. https://doi.org/10.3390/md19070380
APA StyleBoullot, F., Fabioux, C., Hégaret, H., Boudry, P., Soudant, P., & Benoit, E. (2021). Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin. Marine Drugs, 19(7), 380. https://doi.org/10.3390/md19070380