
Identification and Biochemical Characterization of a Surfactant-Tolerant Chondroitinase VhChlABC from Vibrio hyugaensis LWW-1


Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Strain LWW-1
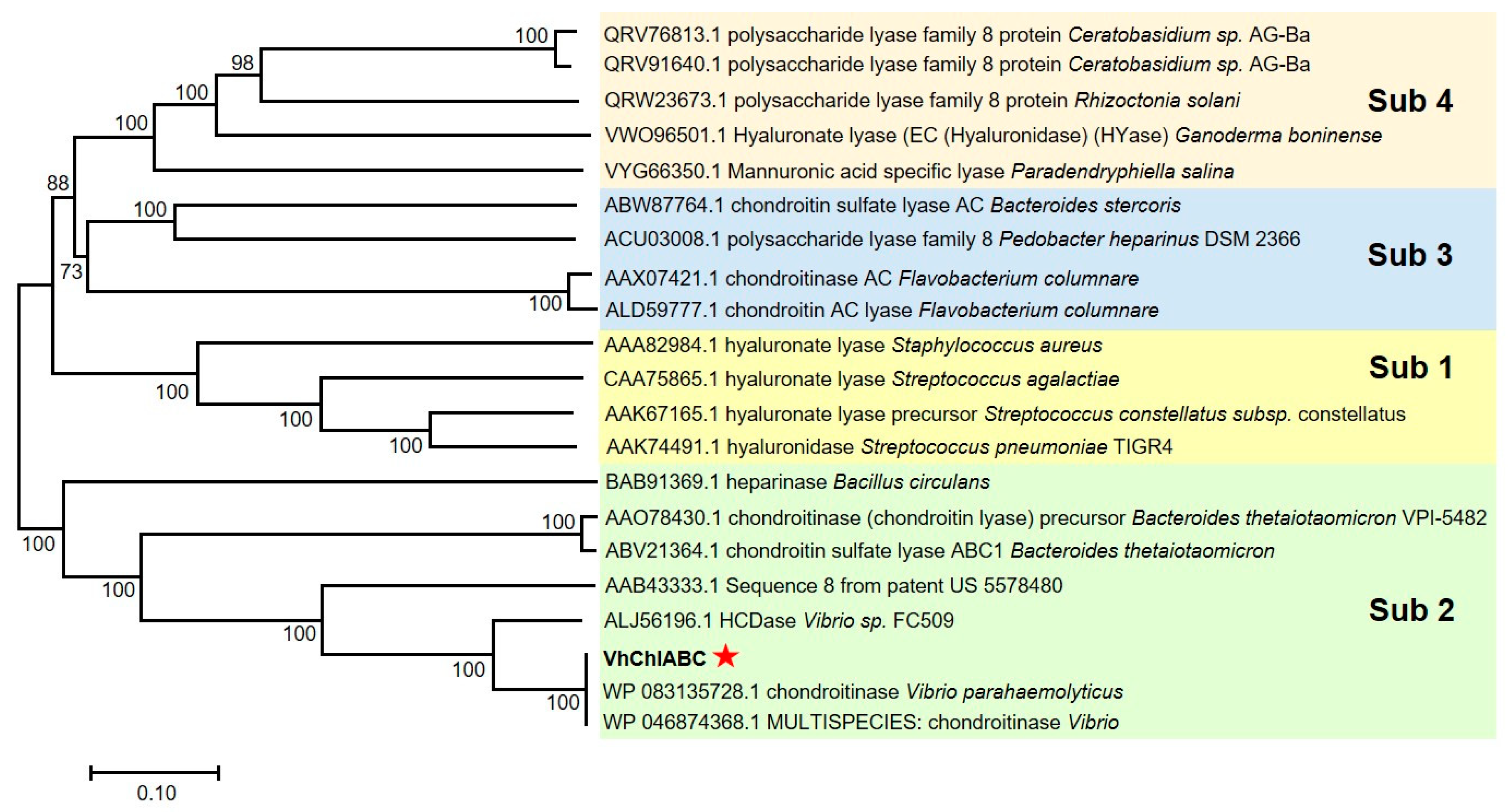
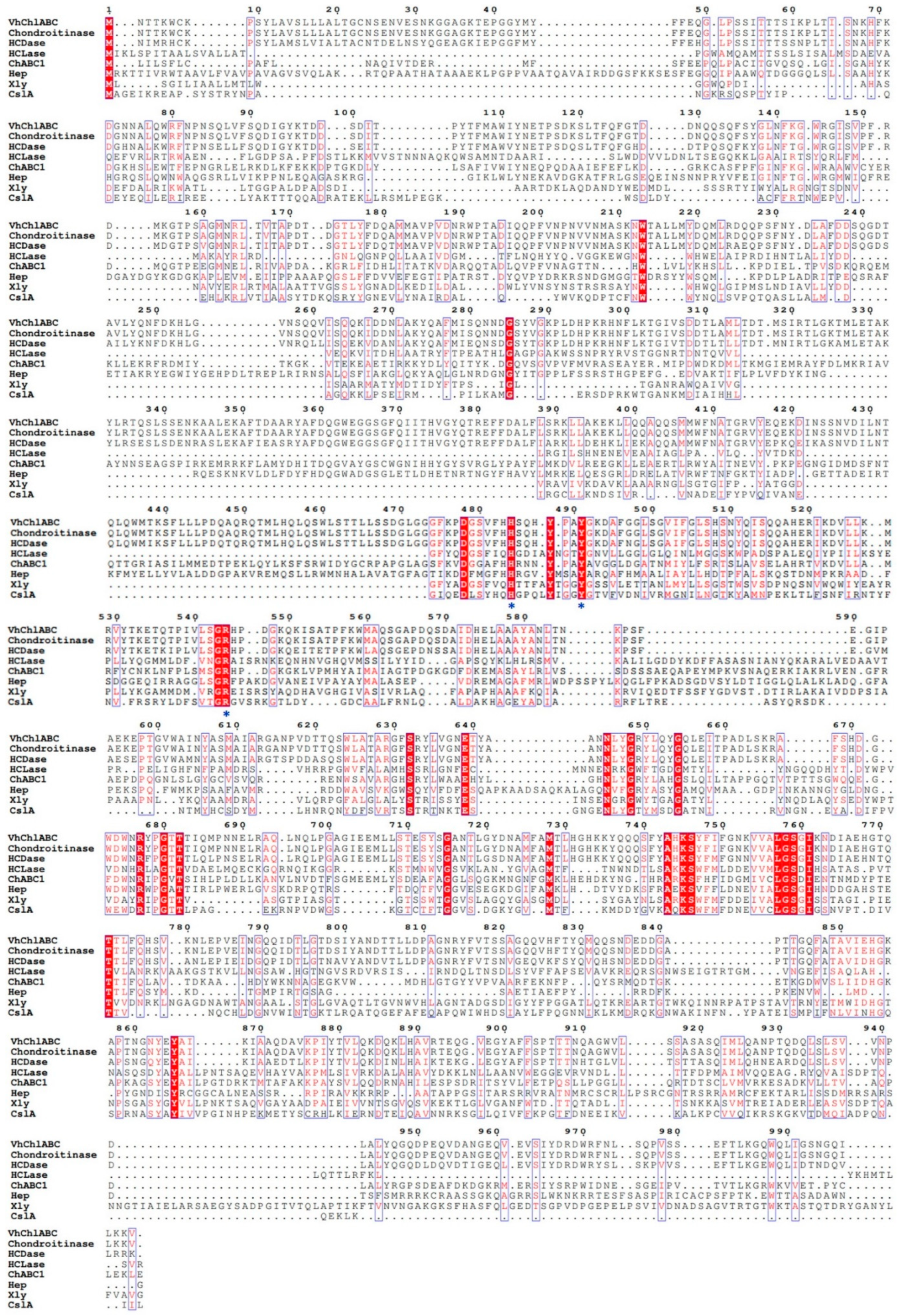
2.2. Identification and Sequence Analysis of CS-Degrading Enzyme
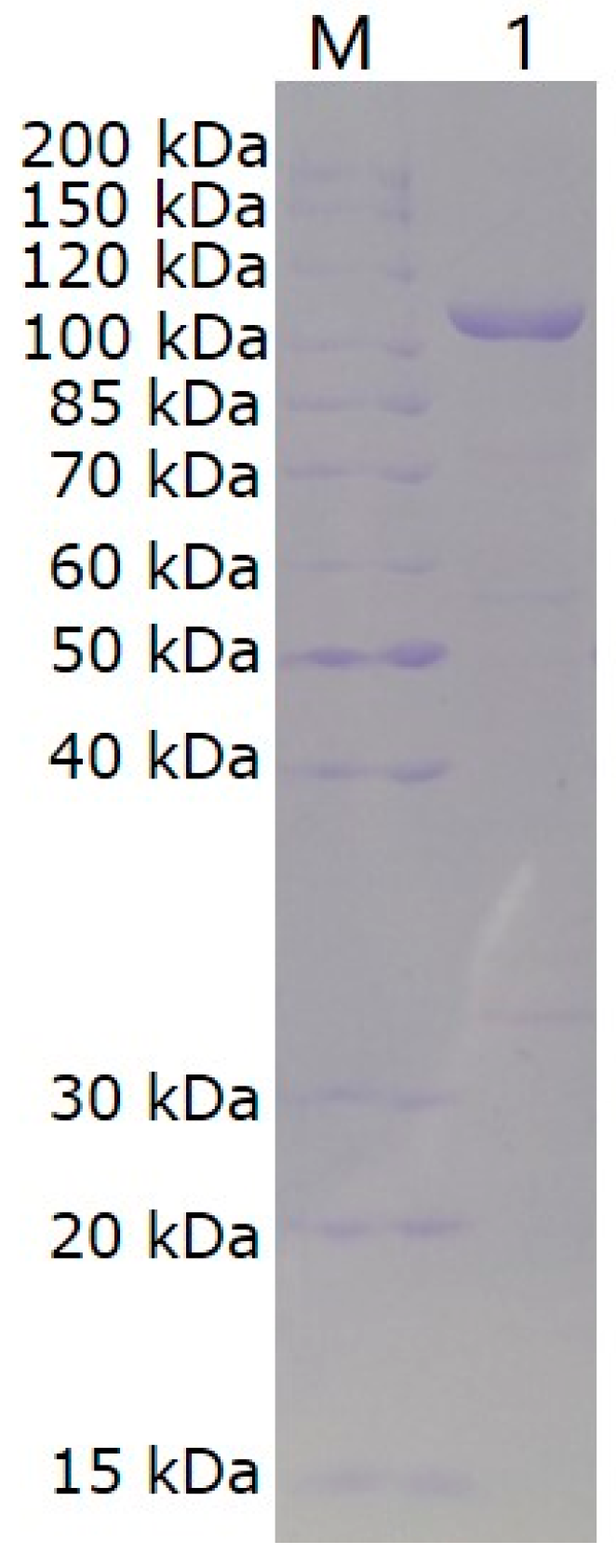
2.3. Recombinant Expression and Purification of VhChlABC
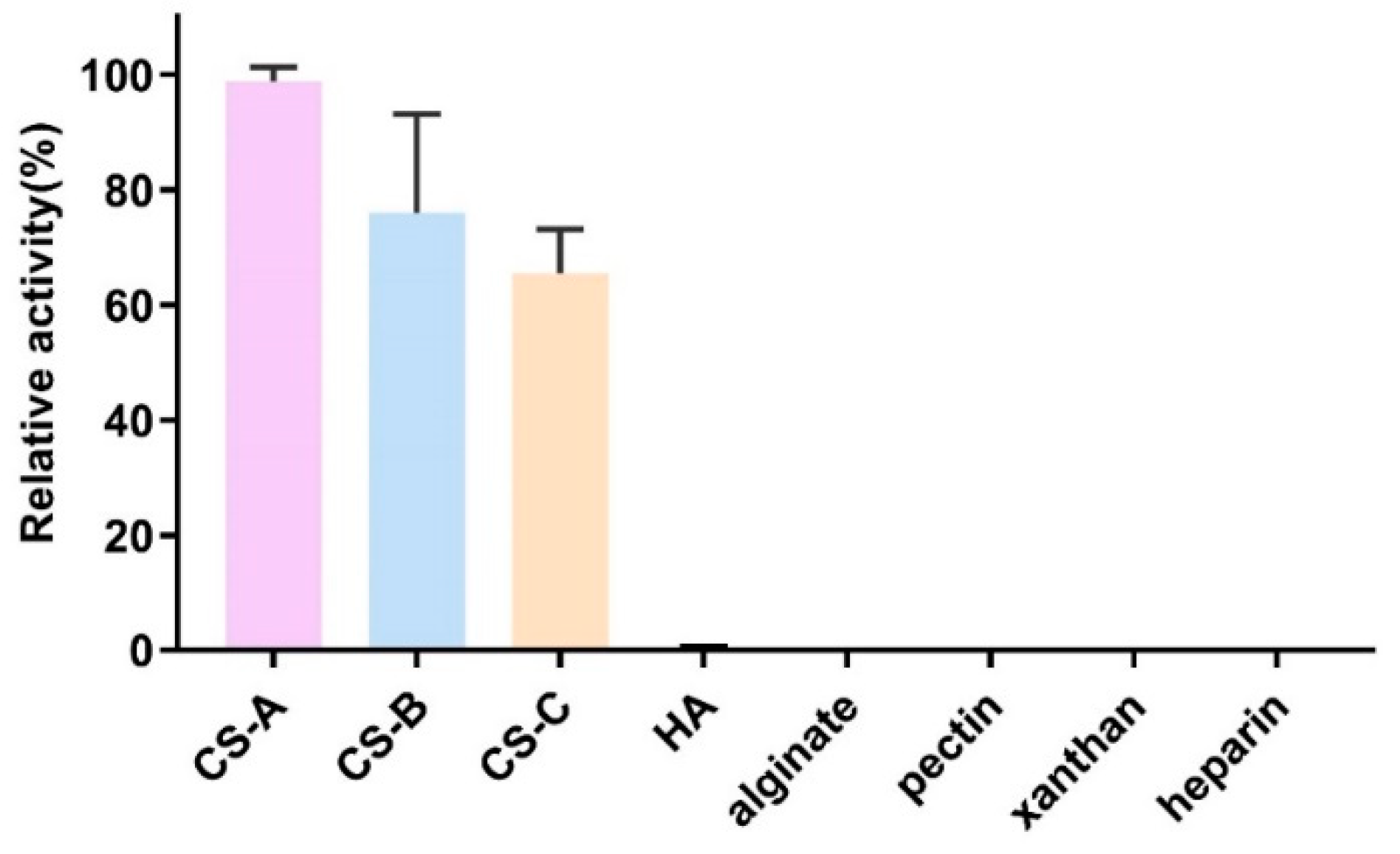
2.4. Substrate Specificity of VhChlABC
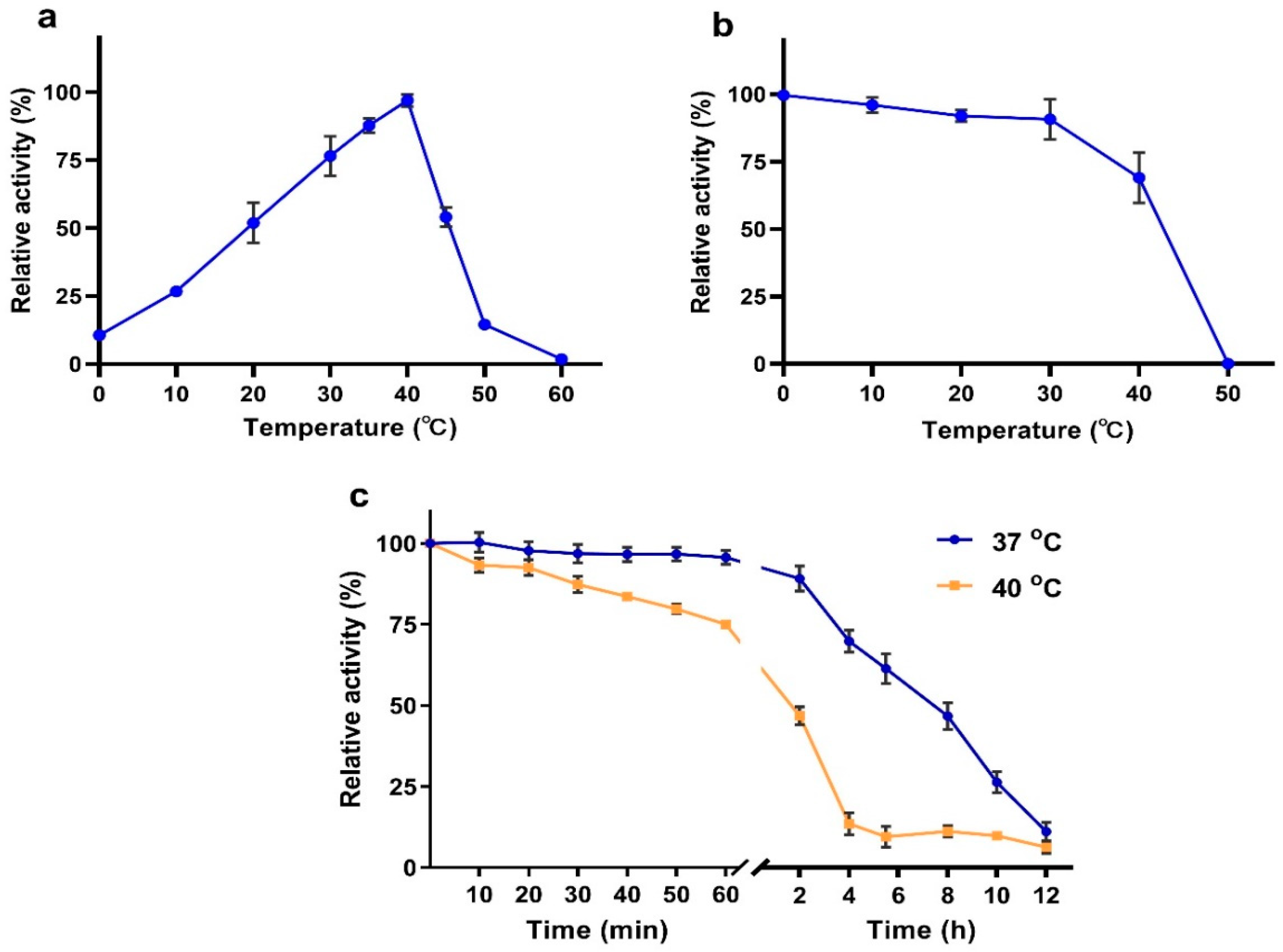
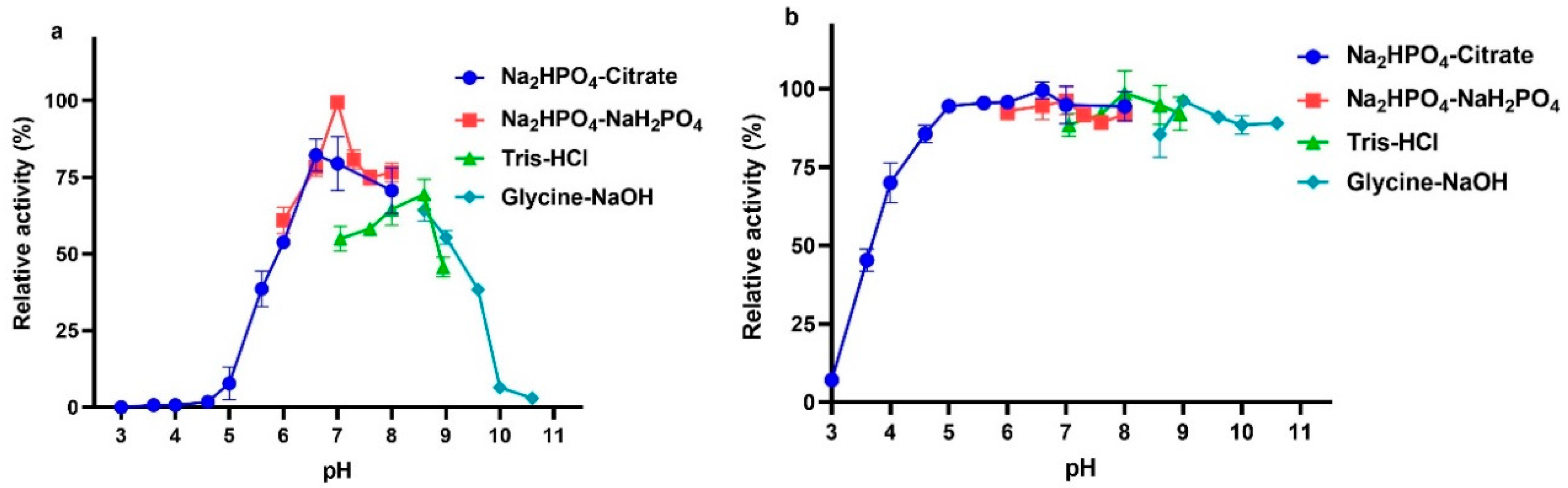
2.5. Effects of Temperature and pH on VhChlABC
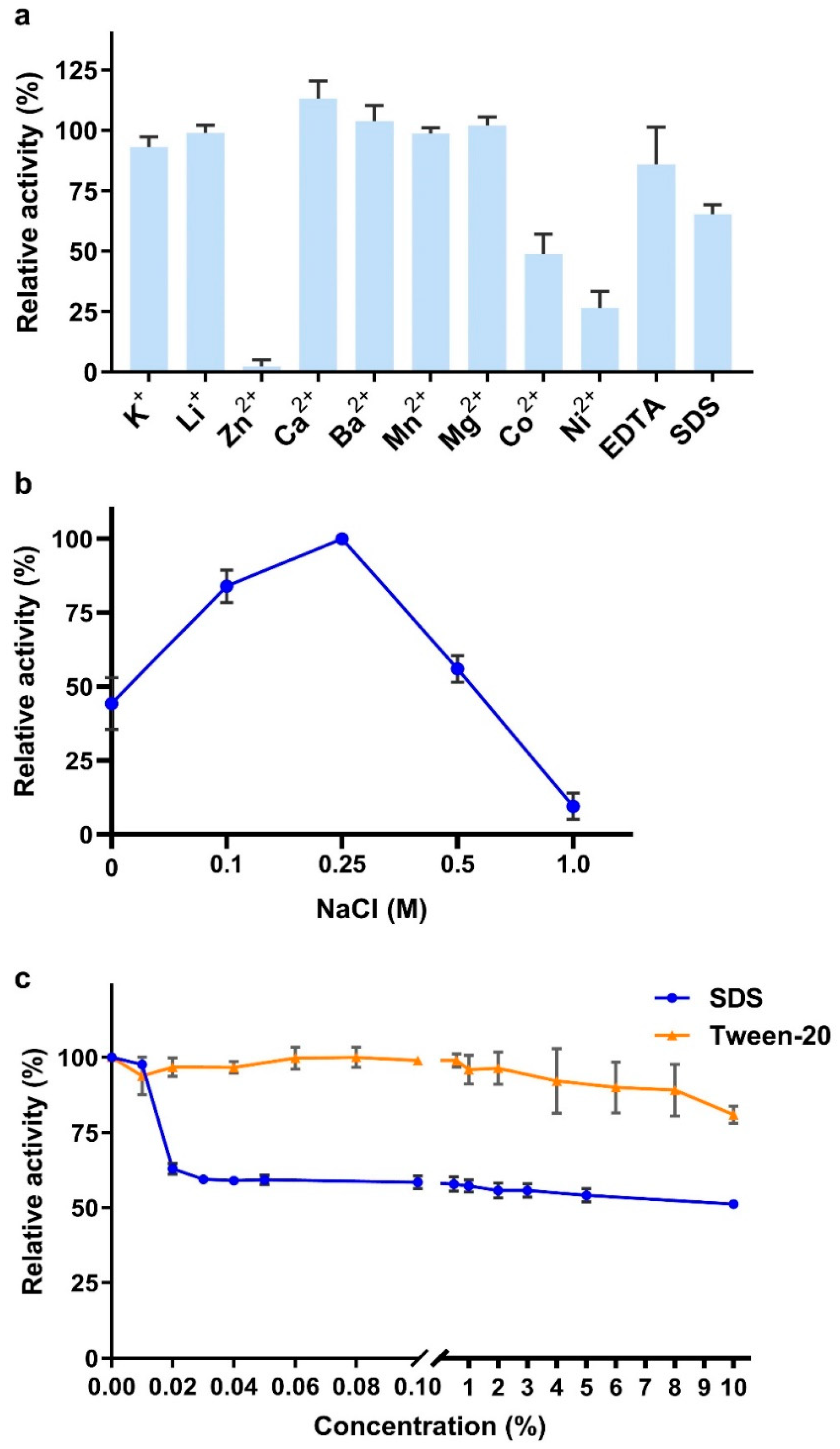
2.6. Effects of Metal Ions, Chelators, and Surfactants on VhChlABC
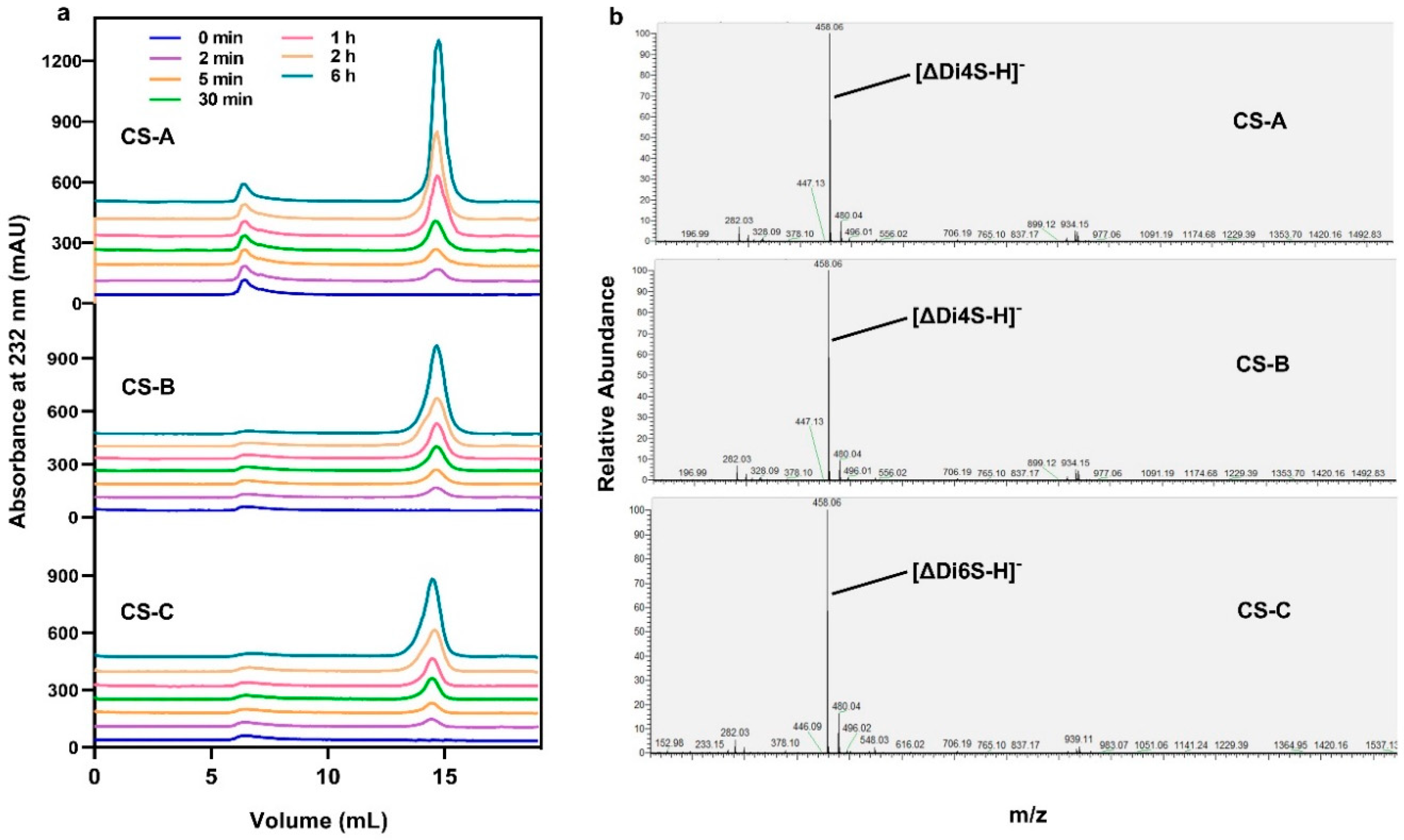
2.7. Degradation Mode and End Products of VhChlABC
3. Materials and Methods
3.1. Reagents
3.2. Isolation of Marine CS Lyase-Producing Bacteria
3.3. Identification of Strain LWW-1
3.4. Sequence Analysis of VhChlABC
3.5. Recombinant Expression and Purification of VhChlABC
3.6. Enzyme Activity Determination of VhChlABC
3.7. Substrates Specificity Analysis of VhChlABC
3.8. Biochemical Characterization of VhChlABC
3.9. Reaction Kinetics of VhChlABC towards CS
3.10. Reaction Mode and End Products of VhChlABC
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
Appendix B

References
- Kreuger, J.; Spillmann, D.; Li, J.P.; Lindahl, U. Interactions between heparan sulfate and proteins: The concept of specificity. J. Cell Biol. 2006, 174, 323–327. [Google Scholar] [CrossRef]
- Ly, M.; Laremore, T.N.; Linhardt, R.J. Proteoglycomics: Recent progress and future challenges. Omics 2010, 14, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishnoi, M.; Jain, A.; Hurkat, P.; Jain, S.K. Chondroitin sulphate: A focus on osteoarthritis. Glycoconj. J. 2016, 33, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.; Kitagawa, H. Recent advances in the study of the biosynthesis and functions of sulfated glycosaminoglycans. Curr. Opin. Struct. Biol. 2000, 10, 518–527. [Google Scholar] [CrossRef]
- Mikami, T.; Kitagawa, H. Biosynthesis and function of chondroitin sulfate. Biochim. Biophys. Acta 2013, 1830, 4719–4733. [Google Scholar] [CrossRef]
- Trowbridge, J.M.; Rudisill, J.A.; Ron, D.; Gallo, R.L. Dermatan sulfate binds and potentiates activity of keratinocyte growth factor (FGF-7). J. Biol. Chem. 2002, 277, 42815–42820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, S.; Akhter, M.P.; Seifert, M.F.; Dai, X.M.; Stanley, E.R. Developmental and functional significance of the CSF-1 proteoglycan chondroitin sulfate chain. Blood 2006, 107, 786–795. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent advances in the structural biology of chondroitin sulfate and dermatan sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef]
- Rani, A.; Patel, S.; Goyal, A. Chondroitin Sulfate (CS) Lyases: Structure, Function and Application in Therapeutics. Curr. Protein Pept. Sci. 2018, 19, 22–33. [Google Scholar] [CrossRef]
- Rauch, U.; Kappler, J. Chondroitin/Dermatan sulfates in the central nervous system: Their structures and functions in health and disease. Adv. Pharmacol. 2006, 53, 337–356. [Google Scholar]
- Shioiri, T.; Tsuchimoto, J.; Watanabe, H.; Sugiura, N. Sequence determination of synthesized chondroitin sulfate dodecasaccharides. Glycobiology 2016, 26, 592–606. [Google Scholar] [CrossRef] [Green Version]
- Clement, A.M.; Nadanaka, S.; Masayama, K.; Mandl, C.; Sugahara, K.; Faissner, A. The DSD-1 carbohydrate epitope depends on sulfation, correlates with chondroitin sulfate D motifs, and is sufficient to promote neurite outgrowth. J. Biol. Chem. 1998, 273, 28444–28453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasinathan, N.; Volety, S.M.; Josyula, V.R. Chondroitinase: A promising therapeutic enzyme. Crit. Rev. Microbiol. 2016, 42, 474–484. [Google Scholar] [CrossRef]
- Rosenzweig, E.S.; Salegio, E.A.; Liang, J.J.; Weber, J.L.; Weinholtz, C.A.; Brock, J.H.; Moseanko, R.; Hawbecker, S.; Pender, R.; Cruzen, C.L.; et al. Chondroitinase improves anatomical and functional outcomes after primate spinal cord injury. Nat. Neurosci. 2019, 22, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.W.; Maffei, L. Reactivation of ocular dominance plasticity in the adult visual cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef] [Green Version]
- Vorobyov, V.; Kwok, J.C.; Fawcett, J.W.; Sengpiel, F. Effects of digesting chondroitin sulfate proteoglycans on plasticity in cat primary visual cortex. J. Neurosci. 2013, 33, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Wegrowski, Y.; Maquart, F.X. Chondroitin sulfate proteoglycans in tumor progression. Adv. Pharmacol. 2006, 53, 297–321. [Google Scholar]
- Denholm, E.M.; Lin, Y.Q.; Silver, P.J. Anti-tumor activities of chondroitinase AC and chondroitinase B: Inhibition of angiogenesis, proliferation and invasion. Eur. J. Pharmacol. 2001, 416, 213–221. [Google Scholar] [CrossRef]
- Prabhakar, V.; Capila, I.; Soundararajan, V.; Raman, R.; Sasisekharan, R. Recombinant expression, purification, and biochemical characterization of chondroitinase ABC II from Proteus vulgaris. J. Biol. Chem. 2009, 284, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lu, D.; Wang, S.; Wei, L.; Wang, W.; Li, F. Identification and biochemical characterization of a novel chondroitin sulfate/dermantan sulfate lyase from Photobacterium sp. Int. J. Biol. Macromol. 2020, 165, 2314–2325. [Google Scholar] [CrossRef] [PubMed]
- Muir, E.; De Winter, F.; Verhaagen, J.; Fawcett, J. Recent advances in the therapeutic uses of chondroitinase ABC. Exp. Neurol. 2019, 321, 113032. [Google Scholar] [CrossRef]
- Prabhakar, V.; Capila, I.; Bosques, C.J.; Pojasek, K.; Sasisekharan, R. Chondroitinase ABC I from Proteus vulgaris: Cloning, recombinant expression and active site identification. Biochem. J. 2005, 386, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Shaya, D.; Hahn, B.S.; Park, N.Y.; Sim, J.S.; Kim, Y.S.; Cygler, M. Characterization of chondroitin sulfate lyase ABC from Bacteroides thetaiotaomicron WAL2926. Biochemistry 2008, 47, 6650–6661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Zhang, J.; Zhang, J.; Jiang, Y.; Shen, Z.; Guan, H.; Jiang, X. Purification and characterization of chondroitinase ABC from Acinetobacter sp. C26. Int. J. Biol. Macromol. 2017, 95, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Jiang, Z.; Chang, J.; Han, B.; Liu, W.; Peng, Y. Purification, characterization of Chondroitinase ABC from Sphingomonas paucimobilis and in vitro cardiocytoprotection of the enzymatically degraded CS-A. Int. J. Biol. Macromol. 2018, 115, 737–745. [Google Scholar] [CrossRef]
- Wang, W.; Cai, X.; Han, N.; Han, W.; Sugahara, K.; Li, F. Sequencing of chondroitin sulfate oligosaccharides using a novel exolyase from a marine bacterium that degrades hyaluronan and chondroitin sulfate/dermatan sulfate. Biochem. J. 2017, 474, 3831–3848. [Google Scholar] [CrossRef]
- Han, W.; Wang, W.; Zhao, M.; Sugahara, K.; Li, F. A novel eliminase from a marine bacterium that degrades hyaluronan and chondroitin sulfate. J. Biol. Chem. 2014, 289, 27886–27898. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Sun, J.; Zhao, M.; Wang, Z.; Gong, Q.; Yu, W. Cloning and characterization of two chondroitin sulfate ABC lyases from Edwardsiella tarda LMG2793. Enzym. Microb. Technol. 2021, 143, 109701. [Google Scholar] [CrossRef]
- Hong, S.W.; Kim, B.T.; Shin, H.Y.; Kim, W.S.; Lee, K.S.; Kim, Y.S.; Kim, D.H. Purification and characterization of novel chondroitin ABC and AC lyases from Bacteroides stercoris HJ-15, a human intestinal anaerobic bacterium. Eur. J. Biochem. 2002, 269, 2934–2940. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Linhardt, R.J.; Laliberté, M.; Gu, K.; Zimmermann, J. Purification, characterization and specificity of chondroitin lyases and glycuronidase from Flavobacterium heparinum. Biochem. J. 1995, 312 Pt 2, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Hamai, A.; Hashimoto, N.; Mochizuki, H.; Kato, F.; Makiguchi, Y.; Horie, K.; Suzuki, S. Two distinct chondroitin sulfate ABC lyases. An endoeliminase yielding tetrasaccharides and an exoeliminase preferentially acting on oligosaccharides. J. Biol. Chem. 1997, 272, 9123–9130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.; Nikapitiya, C.; Lee, Y.; Whang, I.; Kim, S.J.; Kang, D.H.; Lee, J. Cloning, purification and biochemical characterization of beta agarase from the marine bacterium Pseudoalteromonas sp. AG4. J. Ind. Microbiol. Biotechnol. 2010, 37, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Bulut, H.; Valjakka, J.; Yuksel, B.; Yilmazer, B.; Turunen, O.; Binay, B. Effect of Metal Ions on the Activity of Ten NAD-Dependent Formate Dehydrogenases. Protein J. 2020, 39, 519–530. [Google Scholar] [CrossRef]
- Jimoh, A.A.; Lin, J. Biosurfactant: A new frontier for greener technology and environmental sustainability. Ecotoxicol. Environ. Saf. 2019, 184, 109607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Su, H.; Wang, X.; Tang, L.; Hu, J.; Yu, W.; Han, F. Cloning and characterization of a novel chondroitinase ABC categorized into a new subfamily of Polysacch. lyase family 8. Int. J. Biol. Macromol. 2020, 164, 3762–3770. [Google Scholar] [CrossRef]
- Dmitrieva, N.; Yu, L.; Viapiano, M.; Cripe, T.P.; Chiocca, E.A.; Glorioso, J.C.; Kaur, B. Chondroitinase ABC I-mediated enhancement of oncolytic virus spread and antitumor efficacy. Clin. Cancer Res. 2011, 17, 1362–1372. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Meng, H.; Gu, D.; Li, Y.; Jia, M. Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol. Res. 2019, 222, 43–51. [Google Scholar] [CrossRef]
- Patil, S.; Chaudhari, B. A simple, rapid and sensitive plate assay for detection of microbial hyaluronidase activity. J. Basic Microbiol. 2017, 57, 358–361. [Google Scholar] [CrossRef]
- Yamagata, T.; Saito, H.; Habuchi, O.; Suzuki, S. Purification and properties of bacterial chondroitinases and chondrosulfatases. J. Biol. Chem. 1968, 243, 1523–1535. [Google Scholar] [CrossRef]
- Han, Y.; Yu, R.; Gao, P.; Lu, X.; Yu, W. The hydrogen-bond network around Glu160 contributes to the structural stability of chitosanase CsnA from Renibacterium sp. QD1. Int. J. Biol. Macromol. 2018, 109, 880–887. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Specific Activity (U/mg) | Total Protein (mg) | Total Activity (U) | Fold Purification | Yield (%) |
|---|---|---|---|---|---|
| Fermentation broth | 3.66 | 106.22 | 388.77 | 1 | 100 |
| Nickel column | 17.54 | 3.70 | 64.90 | 4.79 | 16.69 |
| Substrate | Km (μM) | Vmax (μmol·min−1) | kcat (min−1) | kcat/Km (min−1·mM−1) |
|---|---|---|---|---|
| CS-A | 2.90 ± 0.35 | 1.06 ± 0.04 | 5310 ± 200 | 1.83 ± 0.069 |
| CS-B | 4.29 ± 0.33 | 0.90 ± 0.02 | 4468 ± 123 | 1.04 ± 0.029 |
| CS-C | 2.67 ± 0.22 | 0.70 ± 0.01 | 3465 ± 72 | 1.30 ± 0.027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Wang, X.; Yin, C.; Li, Y.; Wu, H.; Yu, W.; Han, F. Identification and Biochemical Characterization of a Surfactant-Tolerant Chondroitinase VhChlABC from Vibrio hyugaensis LWW-1. Mar. Drugs 2021, 19, 399. https://doi.org/10.3390/md19070399
Su J, Wang X, Yin C, Li Y, Wu H, Yu W, Han F. Identification and Biochemical Characterization of a Surfactant-Tolerant Chondroitinase VhChlABC from Vibrio hyugaensis LWW-1. Marine Drugs. 2021; 19(7):399. https://doi.org/10.3390/md19070399
Chicago/Turabian StyleSu, Juanjuan, Xiaoyi Wang, Chengying Yin, Yujiao Li, Hao Wu, Wengong Yu, and Feng Han. 2021. "Identification and Biochemical Characterization of a Surfactant-Tolerant Chondroitinase VhChlABC from Vibrio hyugaensis LWW-1" Marine Drugs 19, no. 7: 399. https://doi.org/10.3390/md19070399
APA StyleSu, J., Wang, X., Yin, C., Li, Y., Wu, H., Yu, W., & Han, F. (2021). Identification and Biochemical Characterization of a Surfactant-Tolerant Chondroitinase VhChlABC from Vibrio hyugaensis LWW-1. Marine Drugs, 19(7), 399. https://doi.org/10.3390/md19070399